
Abstract
(1) The role of activation of Rho-kinase in the pathogenesis of cognitive deficit and neuronal damage caused by chronic global ischemia is not clear. In this study, hydroxyfasudil, a Rho-kinase inhibitor, was found to improve the learning and memory performance significantly in rats with ischemia induced by chronic cerebral hypoperfusion after permanent bilateral carotid artery ligation (BCAL). This was observed by the administration of hydroxyfasudil (1 mg/kg or 10 mg/kg, once per day for 30 days) to ischemic rats and the measurements of escape latency and time spent in the target quadrant among the ischemic, sham, and ischemic plus hydroxyfasudil rats by the method of Morris water maze. (2) In electrophysiological study, hydroxyfasudil abolished the inhibition of long-term potentiation (LTP) in rats with ischemia. Morphologically, it also markedly reduced pathological changes such as neuronal cells loss and nuclei shrinkage in cortex and hippocampus of ischemic rats. Biochemical analysis showed that the inhibition of Rho-kinase by hydroxyfasudil reduced the amount of MDA and increased the activities of SOD and GPx in ischemic rats that had increased MDA and decreased SOD and GPx activities. (3) To explore mechanism (s) of the beneficial effects of hydroxyfasudil in ischemia, we performed immunohistochemistry and RT-PCR analyses of NMDA NR2B subunit and for the first time found that hydroxyfasudil increased the expression of NR2B in cortex and hippocampus at both protein and mRNA levels. (4) Taken together, our data further support the notion that the inhibition of Rho-kinase provides neuroprotective effects in cerebral ischemia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rho, a small GTP-binding protein, may act as a molecular switch in various cellular functions including vascular smooth muscle cell contraction (Fukata et al. 2001), actin cytoskeleton organization (van Nieuw Amerongen and van Hinsbergh 2001), cell adhesion and motility (Horwitz and Parsons 1999), cytokinesis (Narumiya 1996), and gene expression (Takemoto et al. 2002). It can modulate the Ca2+ sensitization of vascular smooth muscle cells by inhibiting myosin phosphatase activity. Therefore, there have been some research interests on the effect of Rho on the tonus of blood vessels and the role of Rho in the pathogenesis of vasospasm (Tachibana et al. 1999), arteriosclerosis (Shimokawa et al. 2002), and hypertension (Masumoto et al. 2001). Among known Rho effectors, Rho-associated Rho-kinase is best characterized. Hydroxyfasudil, a synthetic compound, is a specific inhibitor of Rho kinase (Stephen et al. 2000). It can be a very useful tool for elucidating the roles of Rho/Rho-kinase pathway in vitro and in vivo. Although the fasudil has anti-vasospastic effect which has been used for the treatment of cerebral vasospasm after subarachnoid hemorrhage (Tachibana et al. 1999), the role of Rho/Rho kinase pathway in ischemic brain damage remains incompletely understood.
It was reported that hydroxyfasudil significantly improved neurological functions and reduced the size of infarct in the brain, at least in two animal models of transient focal cerebral ischemia brought on by middle cerebral artery occlusion in mice (Yoshiyuki et al. 2005) and cerebral microembolization in rats (Shin-ichi et al. 2001). Inactivation of Rho with clostridium botulinum exoenzyme C3 transferase promoted neurite outgrowth (Lehmann et al. 1999). Similarly, inhibition of p160ROCK, a downstream effector of Rho, with Y-27632 facilitated neurite outgrowth and accelerated locomotor recovery after corticospinal tract lesions in adult rats (Fournier et al. 2003). Therefore, the Rho/ROCK pathway may also have an important role in the pathophysiology of neuronal injury (Sung et al. 2003).
In contrast, the role of the Rho/Rho-kinase pathway in chronic ischemia brain damage has not been well studied. It has been reported that the reductions of local cerebral blood flow and local cerebral glucose utilization in forebrain regions with chronic global ischemia after BCAL in conscious Wistar rats could be overcome by rho kinase inhibitor HA1077 (Satoh et al. 1996). However, whether Rho/Rho-kinase is involved in the pathogenesis of cognitive deficit and neuronal damage induced by chronic global ischemia remains unknown. In the present study, we have investigated the relationship of Rho/Rho-kinase pathway and the progression of memory deficit and neuronal injury using Rh-kinase inhibitor hydroxyfasudil in the rats with ischemia induced by BCAL. It has been known that Rho/Rho-kinase pathway is involved in the modulation of N-methyl-d-aspartate (NMDA) receptor function (Nakazawa et al. 2003; Noerenberg et al. 1999). NMDA R2B (NR2B) may be associated with learning and memory processing and disruption of the gene for NR2B in mice causes perinatal lethality (Zhou et al. 2006). Both larger long-term potentiation (LTP) in the hippocampus and superior learning and memory were observed in naive NR2B transgenic mice even after the 10th–12th generation of breeding (Tang et al. 2001). In addition, angiotensin II infusion causes Rho-kinase activation and NADPH oxidase subunit mRNA expression in rat aorta, both of which are significantly suppressed by the Rho-kinase inhibitor fasudil (Higashi et al. 2003). Therefore, we have also explored potential mechanisms responsible for inhibition of Rho-kinase activity by hydroxyfasudil in chronic ischemia.
Materials and Methods
Chemicals
Hydroxyfasudil was from Tocris Cookson Ltd. (Britol, UK). Assay kits for malondialdehyde (MDA), superoxide dismutase (SOD), glutathione peroxidase (GPx), and total protein quantitation were purchased from Nanjing Jiancheng Bio-Tek Co. (Nanjing, China). Rabbit anti-mouse NR2B monoclonal IgG was from Santa Cruz Biotechnology (Santa Cruz, CA). Trizol reagent was obtained from GIBCO (Grand Island, NY). Taq DNA polymerase and reverse transcription kit were from Promega (Madison, WI, USA). RNase A, dNTP, and Oligo (dT) were from Sigma (St. Louis, MO, USA).
Animals
Adult male Sprague-Dawley rats weighing 200–250 g were used (Experimental Animal Center, Tongji Medical College, Huazhong University of Science and Technology). They were housed five per cage in a temperature and humidity-controlled room (temperature: 22 ± 1°C, humidity: 60%) with free access to food and water. The rats were kept on a 12-h light/dark cycle and adapted to these conditions for at least 7 days before experiments. All rats were randomly divided into groups of sham, ischemia and ischemia treated with hydroxyfasudil (1 mg/kg and 10 mg/kg). The sham group was treated with only saline without induction of ischemia. The ischemia group was treated with either saline or the drug (hydroxyfasudil, 1 mg/kg and 10 mg/kg) by intraperitoneal injection (IP), once per day for 30 days (d) after ischemia.
Surgery
The rats were anesthetized with chloral hydrate (350 mg/kg, IP). Bilateral common carotid arteries were exposed through a midline neck incision, double ligated with 4–0 type surgical silk, and cut between the ligations in ischemia rats. The sham rats received the same operation except for the last ligation step. During ischemia, body temperature was maintained at 37.5 ± 0.5°C by means of a heating lamp until the rat recovered to thermal homeostasis.
Water Maze Test
The spatial learning and memory performances of rats were evaluated using Morris water maze. The training protocol was similar to that of Bombi et al. (2003). Rats received two blocks of training over five days. Each block consisted of four trials with a 15–20 min rest period between trials. The starting position was randomized among four quadrants, and each trial lasted for 180 s or until the rats were able to locate the platform that was placed approximately 1 cm below the surface of the water and kept in the same position throughout the experiments. Rats that did not find the platform in 180 s were guided to the platform. Latency to escape onto the hidden platform was recorded. On the sixth training day, each rat was subjected to a 180 s probe trial in which the platform was removed and the time spent in Q3 was recorded. To examine the possibility that the difference in spatial learning between groups, if any, was associated with vision impairment, the escape latency of each rat was determined with the visible platform at 1 cm above water surface after the probe trial. Swimming activities were monitored by a video camera linked to a computer-based image analyzer. Tests were performed at hours between 18:00 and 22:00.
Long Term Potentiation
Rats were anesthetized by urethane (1.2 g/kg IP) and the body temperatures kept at 37°C via a constant temperature water cycling system. The animals were fixed in a stereotaxic frame (SN-3, Narishige, Japan) according to bregma and lambda in the same horizontal plane. A stainless steel bipolar stimulating electrode was inserted into the perforant path (6.8–7.0 mm posterior to bregma, 4.3–4.5 mm lateral to midline, and 3.0–4.0 mm vertical to dura). A recording electrode was placed in the pyramidal cell layer of CA3 (3.2–3.5 mm posterior to bregma, 3.2–3.5 mm lateral to midline and 3.0–3.5 mm vertical to dura).
Field responses evoking a frequency of 0.5 Hz and 150 ms in duration were acquired, amplified, monitored and analyzed with SMUP-PC biology signal processing system (Second Military Medical University, China). The amplitude of the population spike (PS) height was measured. The stimulation intensity for the tests and train pulses was set to 50–60% of the maximum population spike amplitude, as determined before the test pulse series. A single test stimulus was delivered at 2-s intervals, and the averaged responses of 10 times were measured every 5 min throughout the experiment. High-frequency stimulation (HFS) consisted of 50 trains at 0.5 Hz, each composed of four pulses at 500 Hz. After tetanus stimulation, the amplitude of the population spike height was measured for 1 h in the perforant path–CA3. The averaged PS amplitude of 10 different time points within an hour before HFS was considered as the baseline value. The percentage of the ratio of absolute PS amplitude to baseline value was used to represent the PS amplitude level. It was defined as a successful induction of LTP if the amplitude of population spike change exceeded 20% (Bliss et al. 1993; Kiyoshi et al. 2001).
Morphology
Four rats chosen randomly from each group were anesthetized with chloral hydrate (350 mg/kg, IP), and then perfused transcardially with normal saline followed by 4% paraformaldehyde. All brains were then fixed in the same fixative at 4°C, dehydrated and then embedded in paraffin blocks. Coronal sections of 5 μm were stained with hematoxylin-eosin (H & E).
Immunohistochemistry
Tissue sections were deparaffinized and wet through graded alcohol. Endogenous peroxidase activity was blocked by incubation in 10% hydrogen peroxide for 10 min. After rinsed with PBS three times, the sections were blocked with 1:10 normal goat serum to minimize nonspecific background staining. After rabbit anti-mouse NR2B monoclonal antibody (dilution 1:100) was applied to the samples and incubated for 24 h at 4°C, the sections were incubated with biotinylated goat anti-rabbit IgG (1:100) for 1 h at 37°C, followed by the steps according to SABC kit protocol. The sections were subsequently incubated with diaminobenzidine 0.5 g/l and observed under light microscope.
Reverse Transcriptase–Polymerase chain reaction
After the rats were decapitated, cerebral cortex and hippocampus were dissected on ice respectively. The RNA isolation was performed by standard procedure with Trizol reagent and the RNA used for assay dissolved in RNase free water. A260/A280 ratios of purified RNA were between 1.6 and 1.8.
Semi-quantitative RT–PCR was done as described previously (Wang et al. 1994; Wong et al. 1994). The glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as housekeeping gene for an internal standard. The primer sequences are as follows (Lai et al. 2000): NR2B 5′-TGCACAATTACTCCTCGACG-3′ and 5′-TCCGATTCTTCTTCTGAGCC-3′ G3PDH 5′-ATGGTGAAGGTCGGTGTGAAC-3′ and 5′-GCTGACAATCTTGAGGGAGT-3′. RT–PCR was performed using Minicycle (M.J. Biotech) and calibrated with GAPDH and NR2B primer pairs. Reverse transcription of 5 μg of total RNA was performed in final volume of 20 μl using 15 units of MMLV reverse transcriptase, 0.5 μg oligo (dT) as a primer and 1 mM of each deoxyribonucleotide during one cycle: at 42°C for 50 min and at 70°C for 15 min with subsequent cooling to 4 °C. PCR was carried out under the following conditions: denaturation at 94 °C for 45 s, annealing at 50 °C for 40 s, and extension at 72°C for 40 s (for the final cycle, extension was 10 min). The cDNAs for NR2B and GAPDH were amplified for 30 cycles. Control amplifications were done either without RT or without RNA. After amplification, PCR products were isolated on a 2% agarose gel containing 200 μg/l ethidium bromides. The size of the amplified DNA fragments for NR2B and GAPDH were 222 and 437 bp respectively. The amount of NR2B mRNA was expressed as a ratio of densitometric readings divided by those of GAPDH. The quantitative results are derived from data analysis with Gel Pro Analyzer 4 program.
Oxidative Stress Parameters
Rats were decapitated and the brain dissected to separate cerebral cortex and hippocampus on ice. The tissues were then homogenized in ice-cold saline of 1:10 (w/v) dilution. The MDA level, SOD, and GPx activities were determined according to the manufacturer’s instructions. Protein concentration was measured by the Coomassie blue protein-binding assay using bovine serum albumin (BSA) as a standard.
Statistical Analysis
All results were presented as mean ± SD. Group differences in the escape latency in the training test of Morris water maze were analyzed by two-way analysis of variance (ANOVA). Data collected from the probe trials, the RT-PCR analysis and the biochemical studies were analyzed using one-way ANOVA followed by Duncan’s test. Group differences of the LTP were analyzed by one-way analysis of variance (ANOVA) followed by the Fisher LSD test (Least-significant difference). Statistical analysis was performed using SPSS 12.0 for windows.
Results
All experiments were performed in rats 30 days after surgeries for BCAL or sham operation. For the relative long-term effect of Rho-kinase inhibition, the administration of hydroxyfasudil was started following surgery to ischemic rats daily for 30 days.
Learning and Memory Performance of Rats Observed by Water Maze Tests
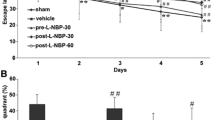
The rats with chronic hypoperfusion spent more time finding the platform than sham-operated rats did [P < 0.05]. This prolonged latency was shortened by relative long-term inhibition of Rho-kinase with hydroxyfasudil [P < 0.05, Fig. 1A]. In the probe trials, the swimming time in the target quadrant (Q3) was used to evaluate the retention performance. Sham group and hydroxyfasudil-treated ischemic group swam longer in Q3 than ischemic group [P < 0.05]. The percentages of time spent in Q3 were 39.89 ± 5.68% for sham group, 20.17 ± 7.87% for ischemia group, 32.00 ± 4.00% and 33.70 ± 4.77% for ischemia group treated with hydroxyfasudil at 1 mg/kg and 10 mg/kg group, respectively (Fig. 1B). Observation of the typical swimming tracks indicated that ischemic rats often searched for the platform in an inappropriate way, resulting in the longer latency to locate the platform (Fig. 1C).

Effect of hydroxyfasudil on water maze performance deficits 30 days after BCAL in rats. (A) Each point represents the mean of escape latency results from the start point onto the hidden platform for each day. (B) The bar graph shows percentage of the time spent in the target quadrant (Q3) in 180 s probe trial (no platform). (C) Typical swim-tracking paths recorded in probe trial on the sixth training day. Following cerebral ischemia, the rats were given intraperitoneal injection of saline or hydroxyfasudil 1 mg/kg or 10 mg/kg, once daily for 30 days. The sham group was given only saline. For A and B, each data point represents mean ± SD, n = 8−10; a P < 0.05 and aa P < 0.01 when compared to sham group; b P < 0.05 and bb P < 0.01 compared to ischemia group
In this study, there was no significant difference for ischemic rats treated with hydroxyfasudil between the low-dose and the high-dose [P > 0.05]. All rats reached the platform in a short period of less than 30 s in the test of a visible platform task, and there was no difference between any of the two groups. This indicated that the difference in spatial performance between groups was unlikely associated with vision impairments.
LTP in CA3 Region of Hippocampus of Rats
The induction rate of LTP in the ischemia group (71.43%) was much lower than that in sham-operated group (100%). Low-dose (1 mg/kg) and high-dose (10 mg/kg) of relative long-term hydroxyfasudil treatment significantly raised LTP from 71.43% in ischemic group to 83.33% and 85.71% respectively [P < 0.05].
LTP was inhibited following BCAL and relative long-term inhibition of Rho-kinase with hydroxyfasudil treatment of ischemic rats restored LTP to the similar level of the sham-operated (Fig. 2A and B). In terms of the population spike height, the average percentage ratios of pre-tetanus to post-tetanus in sham, ischemia, and ischemia plus hydroxyfasudil at 1 mg/kg and 10 mg/kg groups were 187.56 ± 12.21%, 143.65 ± 5.36%, 162.07 ± 9.32%, and 172.36 ± 13.47%, respectively (Fig. 2C). There was no significant difference for ischemic rats treated with hydroxyfasudil between the low-dose and the high-dose [P > 0.05].

In vivo effect of hydroxyfasudil on LTP in the perforant path-CA3 synapses of rats with chronic hypoperfusion after BCAL. (A) PS amplitude was measured before (left) and after (right) tetanus stimulation. LTP was inhibited in the ischemia group as compared to sham group. Hydroxyfasudil at 10 mg/kg abolished the inhibition. (B) The linear graph illustrates the relative PS amplitude alteration at different time point within the 60 min after tetanus stimulation. (C) PS amplitudes calculated from pre-tetanus PS height divided by post-tetanus PS height times 100% (mean ± SD, n = 5−7). Note that significantly higher levels of PS amplitude in hydroxyfasudil treated ischemia group than those in ischemia group alone. a P < 0.05 and aa P < 0.01 when compared to sham group; b P < 0.05 and bb P < 0.01 compared to ischemia group
Morphological Changes of Rat Cortex and Hippocampus
In order to see if the BCAL caused any direct pathological alterations in rats with chronic hypoperfusion, H & E was used to stain the brain tissues of rats as described in the method. In the cortex and hippocampus marked morphological changes were visualized in ischemic group: neuronal cell loss, glial proliferation, nuclei shrinkage, and dark staining of neurons, especially in hippocampal CA1 region. Relative long-term treatment (30 d) in ischemic rats with hydroxyfasudil (10 mg/kg) markedly reduced these pathological changes (Fig. 3).

In vivo effect of hydroxyfasudil on morphologic changes induced by chronic hypoperfusion ischemia in cortical and hippocampal regions of rats. Sections from three rats in each group were examined. a and d: Sham group; b and e: Ischemia group; c and f: Ischemia group treated with hydroxyfasudil 10 mg/kg group. Magnification × 100
Immunohistochemistry and RT-PCR Analyses for Expression of NR2B of Rats
Photomicrographs of immunohistochemical localization of NR2B protein in cortical and hippocampal neuronal cells were illustrated in Fig. 4A. The rats with chronic cerebral hypoperfusion exhibited a remarkable decrease in NR2B reactivity. However, in the hydroxyfasudil-treated ischemia group, obvious increment of immunoreactivity was noted in cortex and hippocampus.

NR2B expression in cortical and hippocampal regions of rats 30 days after BCAL. (A) Immunohistochemistry of NR2B in sham group (a and d), ischemia group (b and e) and ischemia group treated with hydroxyfasudil (c and f). A substantial decrease in NR2B immunoreactivity after ischemia was noted in cortex and hippocampus. Relative long-term hydroxyfasudil treatment (30 days) at 10 mg/kg restored this reactivity. Magnification × 100. (B) RT-PCR analysis of NR2B mRNA in sham group (lane 1), ischemia group (lane 2) and ischemia group treated with hydroxyfasudil (lane 3). (C) The bar graph at bottom shows the quantitative results (mean ± SD, n = 3) of integrated optical density. Data are expressed as a percentage of GAPDH messenger RNA levels in the same samples. Compared with the sham group, there was a markedly decrease in NR2B mRNA levels in the ischemia group, which was elevated by hydroxyfasudil treatment. NR2B: 222 bp product; GAPDH: 473 bp product. The experiments were repeated three times. a P < 0.05 and aa P < 0.01 when compared to sham group; b P < 0.05 and bb P < 0.01 compared to ischemia group
Compared with the sham group, there was a markedly decrease in NR2B mRNA in the ischemia group [P < 0.05]. Relative long-term hydroxyfasudil treatment at high-dose (10 mg/kg) significantly increased the mRNA level of NR2B in the cortex and hippocampus of BCAL rats [P < 0.05] (Fig. 4B and C).
MDA Level, SOD and GPx Activities in Rats 30 Days After BCAL
In sham rats, the concentration of MDA was 6.10 ± 0.69 (nM/mg protein), the activity of SOD 1.05 ± 0.12 (U/mg protein) and GPx 14.8 ± 1.1 (U/mg protein) in the cortex whereas MDA was 5.62 ± 0.9, SOD 1.25 ± 0.23 and GPx 10.4 ± 3.3 in the hippocampus, respectively. As shown in Fig. 5, chronic cerebral hypoperfusion resulted in a significant increase in MDA level (up to 17% and 43%) and markedly decrease in SOD (up to 53% and 44%) and GPx (up to 36% and 24%) activities in the cortex and hippocampus of rats compared to sham group [P < 0.05]. Relative long-term hydroxyfasudil (1 mg/kg or 10 mg/kg) treatment attenuated these changes in chronic ischemia [P < 0.05] and was dose-dependent.

In vivo effect of hydroxyfasudil on MDA level (A), SOD (B) and GPx (C) activities in cortical and hippocampal regions of rats 30 days after BCAL. Compared with sham group, MDA was increased and the activities of SOD and GPx decreased in ischemia group, whereas hydroxyfasudil reduced MDA and enhanced SOD and GPx activities inischemic rats in dose-dependent manner. The data (n = 6; mean ± SD) are expressed as a percentage of the values in sham group for MDA (nM/mg protein], SOD [U/mg protein] and GPx [U/mg protein]. a P < 0.05 and aa P < 0.01 when compared to sham-operated group; b P < 0.05 and bb P < 0.01 compared to ischemia group
Discussion
Stroke is the second most common cause of cognitive impairment and dementia. The accumulation of lacunar infarcts, infarcts, ischemic white matter disease and cerebral hypoperfusion are the most common causes of cognitive impairment/dementia due to stroke. Detecting stroke early and defining appropriate treatment that can prevent or substantially delay the onset and progression of cognitive impairment/dementia are challenging. The present study demonstrates that Rho/Rho-kinase pathways may be involved in the progression of cognitive deficit and neuronal damage in rats with chronic cerebral hypoperfusion after BCAL and Rho-kinase inhibitor hydroxyfasudil could ameliorate the cognitive impairment and neuronal injury in these rats. For the purpose of behavioral study and relative long-term effects of Rho-kinase inhibition, the model of chronic cerebral ischemia was chosen. It was observed that 30 days after the operation, the rats developed marked cognitive deficits along with neuron degeneration. The spatial learning and memory deficits were indicated by longer escape latency and shorter time spent in the target quadrant. The significant neuronal loss and nuclei condensation were also shown in the cortex and hippocampus especially in CA1 region. Therefore the model that was created was in accordance with previous reports (Wang et al. 2000), indicating that chronic cerebral ischemia is related to impairment of learning and memory and to neuronal death. In our experiments, the administration of Rho-kinase inhibitor hydroxyfasudil at either 1 mg/kg or 10 mg/kg to ischemic rats shortened the escape latency and prolonged the time spent in quadrant. The statistic calculation was significant compared to ischemic rats without Rho-kinase inhibition (Fig.1). The histology pictures exhibited the attenuation of the neuronal loss and nuclei condensation after hydroxyfasudil was given to the ischemic rats (Fig. 3). Our data provide novel information for the importance of Rho-kinase inhibition in improving the deficit of cognition and the ability of spatial learning and memorizing in chronic ischemia.
Since the discovery of LTP (Long-Time Potentiation) in the dentate gyrus (Bliss et al. 1973), LTP has also been widely investigated as an electrophysiological basis in learning and memory. The direct perforant path projections to the CA3 and CA1 regions of the hippocampus are substantial and the average pyramidal cells in the CA3 region also receive a direct, monosynaptic input from the entorhinal cortex (EC), via the fibers of the perforant path (PP) (Berzhanskaya et al. 1998; Martinez et al. 2002). Furthermore, the previous studies (Do et al. 2002) and our recent research (Zheng et al. 2006; Sun et al. 2004) demonstrated that stimulation of the perforant path could activate CA3 pyramidal cells and evoked characteristic field potentials. Therefore, the recordings of CA3 region field potential by the stimulation of perforant path are feasible to study the mechanism of hippocampal information process and to reveal the interfering effects of pharmacologic agents. Ischemia or hypoxia could inhibit the synaptic transmission by preventing the production and storage of ATP and neurotransmitter (Lyubkin et al. 1997). Our study showed that the induction rate and the average amplitude of LTP in the ischemia group were lower than those in sham group. Relative long-term administration of hydroxyfasudil abolished nearly total inhibition of LTP in rats with chronic cerebral ischemia (Fig. 2), indicating that hydroxyfasudil could facilitate the synaptic transmission in hippocampus. These electrophysiology data are consistent with our results described as above, in which hydroxyfasudil could ameliorate the learning and memory impairment (Fig.1) and reduce neuronal injury (Fig. 3) in those chronic ischemic rats. Our data are also in line with previous reports that the regulation of Rho GTPase-signaling pathways is important for the generation and maintenance of dendritic spines and branches in hippocampal pyramidal neurons that may eventually contribute to dynamic changes in response to synaptic stimulation under conditions that generate long-term potentiation (Nakayama et al. 2000).
NMDARs are composed of NR1 and NR2 subunits. NR2B has been implicated in modulating functions such as learning, memory processing, and feeding behaviors, as well as being involved in a number of neurological disorders (Jennifer et al. 2003; Tang et al. 1999). Our immunohistochemical study showed a substantial decrease in NR2B immunoreactivity in cortical and hippocampal neurons 30 days after BCAL. When hydroxyfasudil was given to ischemic rats daily for 30 days, the same time length for chronic ischemia, it significantly increased NR2B immunoreactivity, which was decreased in chronic global ischemia (Fig. 4A). Moreover, the RT-PCR analysis showed that NR2B mRNA levels were reduced by BCAL whereas hydroxyfasudil up-regulated the expression of the receptor mRNA in chronic hypo-perfusion rats (Fig. 4B and 4C). Accumulative evidence indicates that NR2B-containing NMDARs exclusively induce LTP (Berberich et al., 2005; Zhao et al., 2005). In addition, overexpression of NR2B in the forebrains of transgenic mice led to enhanced activation of NMDA receptors, facilitating synaptic potentiation in response to stimulation and the mice exhibited superior ability in various tests of learning and memory (novel object recognition task, fear conditioning, fear extinction, and water maze) (Tang et al. 1999). Thus, one of the possible mechanisms for the neuroprotective effect of inhibition of Rho-kinase by hydroxyfasudil might be the enhancement of NR2B mRNA and protein expression.
There are massive data showing that the free radicals are highly reactive molecules that initiate radical chain reactions and damage cellular marcomolecules, ultimately leading to neuron degeneration and death (Coyle et al. 1993). In the present study, we measured the concentration of MDA that is produced by free-radical catalyzed peroxidation of unsaturated fatty acid in cell membranes and the activities of SOD and GPx that play important roles in protection against oxidative stress-induced injury. We observed that rats 30 days after BCAL had increased amount of MDA that was then decreased by hydroxyfasudil (Fig. 5A). In contrast, both SOD and GPx activities were decreased in ischemic rats. Interestingly and importantly, most of their activities were restored by hydroxyfasudil (Fig. 5B and C). Together, systemic oxidative stress was increased and the ability of anti-oxidative stress impaired in this chronic cerebral hypoperfusion model, which was significantly recovered by hydroxyfasudil. Cerebral ischemia causes oxidative stress and produces superoxide by NADPH oxidase in neutrophil (Juurlink and Sweeney 1997). Moreover, a possible correlation of Rho-kinase with oxidative stress has been demonstrated in some in vitro and in vivo studies (Bailey et al. 2005). Other recent studies have shown that angiotensin II stimulates activity of NADPH oxidase, expression of p22phox and p67phox in vascular smooth muscle cells. Its activation is mediated via the Rho/Rho-kinase pathway (Higashi et al. 2003). Therefore, the suppressive effect of hydroxyfasudil on oxidative stress may, in part, be responsible for the beneficially neuroprotective effects of hydroxyfasudil treatment.
In conclusion, our results demonstrated that activation of the Rho/Rho-kinase pathway is related to the neuronal damage and the pathogenesis of the spatial learning and memory impairment in BCAL rats. The relative long-term hydroxyfasudil treatment improved spatial learning and memory performance, increased the induction rate and amplitude of LTP and attenuated neuronal damage in this model. The neuroprotective effects of Rho-kinase inhibition by hydroxyfasudil could be attributed to at least two aspects: (1) upregulation of NR2B mRNA and protein expression; (2) reduction in oxidative stress. However, still little is known about the potential mechanisms of Rho-kinase inhibitor responsible for these actions. These preliminary data merit further studies to elucidate the underlying mechanisms in details.
References
Bailey SR, Mitra S, Flavahan S, Flavahan NA (2005) Reactive oxygen species from smooth muscle mitochondria initiate cold-induced constriction of cutaneous arteries. Am J Physiol Heart Circ Physiol 289:H243–H250
Berberich S, Punnakkal P, Jensen V, Pawlak V, Seeburg PH, Hvalby O, Köhr G (2005) Lack of NMDA receptor subtype selectivity for hippocampal long-term potentiation. J Neurosci 25:6907–6910
Berzhanskaya J, Urban NN, Barrionuevo G (1998) Electrophysiological and pharmacological characterization of the direct perforant path input to hippocampal area CA3. J Neurophysiol 79:2111–2118
Bliss TVP, Lomo T (1973) Long-lasting potentiation of synaptic transmission in the denate area of the perforant path. J Physiol 232:331–56
Bliss TVP, Collingridge GL (1993) A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361(6407):31–39
Bombi L, YoungKook C, Hocheol K (2003) Protective effects of methanol extract of Acori graminei rhizome and Uncariae Ramulus et Uncus on ischemia-induced neuronal death and cognitive impairments in the rat. Life Sciences 74:435–450
Coyle JT, Puttfarcken P (1993) Oxidative stress, glutamate, and neurodegenerative disorders. Science 262:689–695
Do VH, Martinez CO, Martinez JL, Derrick BE (2002) Long-term potentiation in direct perforant path projections to the hippocampal CA3 region in vivo. J Neurophysiol 87:669–678
Fournier AE, Takizawa BT, Strittmatter SM (2003) Rho kinase inhibition enhances axonal regeneration in the injured CNS. J Neurosci 23:1416–1423
Fukata Y, Amano M, Kaibuchi K (2001) Rho-Rho-kinase pathway in smooth muscle contraction and cytoskeletal reorganization of non-muscle cells. Trends Pharmacol Sci 22:32–39
Higashi M, Shimokawa H, Hattori T, Hiroki J, Mukai Y, Morikawa K, Ichiki T, Takahashi S, Takeshita A (2003) Long-term inhibition of Rho-kinase suppresses angiotensin II-induced cardiovascular hypertrophy in rats in vivo: effect on endothelial NAD (P) H oxidase system. Circ Res 93: 767–775
Horwitz AR, Parsons JT (1999) Cell migration: movein’ on. Science 286:1102–1103
Jennifer ML, Aaron J (2003) The N-methyl-d-aspartate receptor subunit NR2B: localization, functional properties, regulation, and clinical implications. Pharmacol Therap 97:55–85
Juurlink BH, Sweeney MI (1997) Mechanisms that result in damage during and following cerebral ischemia. Neurosci Biobehav Rev 21:121–128
Kiyoshi M, Hiroko T, Ken-ichi, Ueno, Machiko M, Mitsuhiro Y (2001) Aminoguanidine prevented the impairment behavior and hippocampal long-term potentiation following transient cerebral ishemia. Behavioural. Brain Research 120:159–169
Lai SK, Wong CKC, Yang MS, Yung KKL (2000) Changes in expression of N-methyl-d-aspartate receptor subunits in the rat neostriatum after a single dose of antisense oligonucleotide specific for N-methyl-d-aspartate receptor 1 subunit. Neuroscience 98:493–500
Lehmann M, Fournier A, Selles-Navarro I, Dergham P, Sebok A, Leclerc N, Tigyi G, McKerracher L (1999) Inactivation of Rho signaling pathway promotes CNS axon regeneration. J. Neurosci 19:7537–7547
Lyubkin M, Durand DM, Haxhiu MA (1997) Interaction between tetanus Long-termpotentiation and hypoxia-induced potentiation in the rat hippocampus. Neurophysiol 78(5):2475–2482
Martinez CO, Do VH, Martinez JL, Derrick BE (2002) Associative long-term potentiation (LTP) among extrinsic afferents of the hippocampal CA3 region in vivo. Brain Res 940:86–94
Masumoto A, Hirooka Y, Shimokawa H, Hironaga K, Setoguchi S, Takeshita A (2001) Possible involvement of Rho-kinase in the pathogensis of hypertension in humans. Hypertension 38:1307–1310
Nakayama AY, Harms MB, Luo L (2000) Small GTPases Rac and Rho in the maintenance of dendritic spines and branches in hippocampal pyramidal neurons. J Neurosci 20:5329–5338
Nakazawa T, Watabe AM, Tezuka T, Yoshida Y, Yokoyama K, Umemori H, Inoue A, Okabe S, Manabe T, Yamamoto T (2003) p250GAP, a novel brain-enriched GTPase-activating protein for Rho family GTPases, is involved in the N-methyl-d-aspartate receptor signaling. Mol Biol Cell 14:2921–2934
Narumiya S (1996) The small GTPase Rho: cellular functions and signal transduction. J Biochem (Tokyo) 120:215–228
Noerenberg W, Hofmann F, Illes P, Aktories K, Meyer DK (1999) Rundown of somatodendritic N-methyl-d-aspartate (NMDA) receptor channels in rat hippocampal neurones: evidence for a role of the small GTPase RhoA. Br J Pharmacol 127:1060–1063
Satoh S, Ikegaki I, Suzuki Y, Asano T, Shibuya M, Hidaka H (1996) Neuroprotective properties of a protein kinase inhibitor against ischaemia-induced neuronal damage in rats and gerbils. Br J Pharmacol 118:1592–1596
Shimokawa H, Hiramori K, Iinuma H, Hosoda S, Kishida H, Osada H, Katagiri T, Yamauchi K, Yui Y, Minamino T, Nakashima M, Kato K (2002) Anti-anginal effect of fasudil, a Rho-kinase inhibitor, in patients with stable effort angina: a multicenter study. J Cardiovasc Pharmacol 40:751–761
Shin-ichi S, Tatsuma U, Kazuyuki T, Teruki K, Ichiro I, Yasuo S, Toshio A (2001) Pharmacological profile of hydroxy fasudil as a selective rho kinase inhibitor on ischemic brain damage. Life Sciences 69:1441–1453
Stephen PD, Helen R, Matilde C, Philip C (2000) Specificity and mechanism of action of some commonly used protein kinase inhibitors. Biochem 351:95–105
Sun T, Hu HZ, Xu XL, Yu SB, Ouyang CH, Qu L, Lv Q, Guo LJ (2004) Effects of glutamic acid and its receptor antagonist on evoked potentials in hippocampus CA3region in rats. Chinese Pharmacol Bull 20l:414–417
Sung JK, Miao L, Calvert JW, Huang L, Harkey HL, Zhang JH (2003) A possible role of RhoA/Rho-kinase in experimental spinal cord injury in rat. Brain Res 959:29–38
Tachibana E, Harada T, Shibuya M, Saito K, Takayasu M, Suzuki Y, Yoshida J (1999) Intra-arterial infusion of fasudil hydrochloride for treating vasospasm following subarachnoid haemorrhage. Acta Neurochir (Wien) 141:13–19
Takemoto M, Sun J, Hiroki J, Shimokawa H, Liao JK (2002) Rho-kinase mediates hypoxia-induced downregulation of endothelial nitric oxide synthase. Circulation 106:57–62
Tang YP, Shimizu E, Dube GR, Rampon C, Kerchner GA, Zhuo M, Liu G, Tsien JZ (1999) Genetic enhancement of learning and memory in mice. Nature 401:63–69
Tang YP, Wang H, Feng R, Kyin M, Tsien JZ (2001) Differential effects of enrichment on learning and memory function in NR2B transgenic mice. Neuropharmacology 41(6):779–790
van Nieuw Amerongen GP, van Hinsbergh VWM (2001) Cytoskeletal effects of Rho-like small guanine nucleotide-binding proteins in the vascular system. Arterioscler Thromb Vasc Biol 21:300–311
Wang LM, Han YF, Tang XC (2000) Huperzine A improves cognitive deficits caused by chronic cerebral hypoperfusion in rats. Eur J Pharmacol 398:65–72
Wong H, Anderson WD, Cheng T, Riabowol KT (1994) Monitoring mRNA expression by polymerase chain reaction: the “primer-dropping” method. Anal Biochem 223:251–258
Yoshiyuki R, Hyung HK, Huang ZH, Minoru S, Kazuo Y, Toshio A, Michael A, Moskowitz, James KL (2005) Inhibition of Rho Kinase (ROCK) leads to increased cerebral blood flow and stroke protection. Stroke 36:2251–2257
Zhao MG, Toyoda H, Lee YS, Wu LJ, Ko SW, Zhang XH, Jia Y, Shum F, Xu H, Li BM, Kaang BK, Zhuo M (2005) Roles of NMDA NR2B subtype receptor in prefrontal long-term potentiation and contextual fear memory. Neuron 47:859–872
Zheng M, Guo LJ, Xu XL, Hu HZ, Zong XG (2006) ZD7288 inhibits the synaptic transmission of the pathway from perforant pathway (PP) fibers to CA3 region in rat hippocampus. Acta Pharmaceutica Sin 41:565–571
Zhou M, Baudry M (2006) Developmental changes in NMDA neurotoxicity reflect developmental changes in subunit composition of NMDA receptors. J Neurosci 26(11):2956–2963
Author information
Authors and Affiliations
Corresponding author
Additional information
Lin Huang and Zhi He contribute equally to the paper.
Rights and permissions
About this article
Cite this article
Huang, L., He, Z., Guo, L. et al. Improvement of Cognitive Deficit and Neuronal Damage in Rats with Chronic Cerebral Ischemia via Relative Long-term Inhibition of Rho-kinase. Cell Mol Neurobiol 28, 757–768 (2008). https://doi.org/10.1007/s10571-007-9157-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10571-007-9157-x