
Abstract
The intergenerational transmission of psychopathology is one of the strongest known risk factors for childhood disorder and may be a malleable target for prevention and intervention. Anxious parents have distinct parenting profiles that impact socioemotional development, and these parenting effects may result in broad alterations to the biological and cognitive functioning of their children. Better understanding the functional mechanisms by which parental risk is passed on to children can provide (1) novel markers of risk for socioemotional difficulties, (2) specific targets for intervention, and (3) behavioral and biological indices of treatment response. We propose a developmental model in which dyadic social dynamics serve as a key conduit in parent-to-child transmission of anxiety. Dyadic social dynamics capture the moment-to-moment interactions between parent and child that occur on a daily basis. In shaping the developmental trajectory from familial risk to actual symptoms, dyadic processes act on mechanisms of risk that are evident prior to, and in the absence of, any eventual disorder onset. First, we discuss dyadic synchrony or the moment-to-moment coordination between parent and child within different levels of analysis, including neural, autonomic, behavioral, and emotional processes. Second, we discuss how overt emotion modeling of distress is observed and internalized by children and later reflected in their own behavior. Thus, unlike synchrony, this is a more sequential process that cuts across levels of analysis. We also discuss maladaptive cognitive and affective processing that is often evident with increases in child anxiety symptoms. Finally, we discuss additional moderators (e.g., parent sex, child fearful temperament) that may impact dyadic processes. Our model is proposed as a conceptual framework for testing hypotheses regarding dynamic processes that may ultimately guide novel treatment approaches aimed at intervening on dyadically linked biobehavioral mechanisms before symptom onset.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Intergenerational transmission is one of the strongest known risk factors for childhood psychopathology. There are three main pathways for intergenerational transmission: (1) genetic inheritance (Eley et al., 2015; Gregory & Eley, 2007); (2) fetal programming through maternal experiences during pregnancy (Lin et al., 2019; Ostlund et al., 2019); and (3) the active and passive socialization of emotion and stress response through parent–child interactions (Askew & Field, 2008; Dunne & Askew, 2013; Hane et al., 2008; Hastings, Rubin, Smith, & Wagner, 2019). Here, we focus on intergenerational pathways in the emergence of anxiety, one of the most common disorders in children affecting 5–10% of children by age 5 (Essau et al., 1999; Fergusson et al., 1993; McGee et al., 1990), with a lifetime prevalence of 32% by age 18 (Beesdo-Baum & Knappe, 2012). Observational studies have reinforced the link between parental anxiety and the emergence of child anxiety, noting high levels of co-morbidity (Biederman et al., 2001; Rosenbaum et al., 1988, 1992). The shared presence of anxiety in the parent–child dyad suggests that we can find unique processes in the emergence of psychopathology that reside within the dyad, rather than the individual, and potentially identify tractable dyadic-level interventions.
In this article, we propose a developmental model in which dyadic social dynamics serve as a key conduit in parent-to-child transmission of anxiety based on prior work illustrating that dyadic parent–child processes contribute something unique to the transmission of risk from parent to child. That is, these dyadic interactions provide explanatory and predictive power even when taking into account individual parent- and child-level variables (Lunkenheimer, Brown, & Fuchs, 2021; Lunkenheimer et al., 2017, 2020; Moore et al., 2013).
The role of biological intermediaries in parent-to-child anxiety transmission is evident in non-human primate (macaque) models of fearful temperament, indicating high heritability (Williamson et al., 2003), which can then be traced to parallel parent–offspring brain function, particularly in the amygdala (Fox et al., 2018; Fox et al., 2015; Fox et al., 2015). Importantly, genetically informed studies in humans note that while genetic factors account for approximately 30–40% of variance in anxiety symptoms (Eley et al., 2015; Hettema et al., 2001), there is a clear additional environmental pathway that helps explain the association between anxiety in the child and parent (Eley et al., 2015). While studies often cannot disentangle genetic and environmental mechanisms of risk, there is clear evidence that targeting parent–child relations can nonetheless decrease anxiety risk in children (Chronis-Tuscano et al., 2018; Danko et al., 2018; Rapee et al., 2010).
Indeed, both clinical and developmental models of psychopathology have long-centered bidirectional or transactional processes that toggle back and forth from parents to children (Bell, 1968; Hudson & Rapee, 2001; Johnco et al., 2021; Sameroff, 1975). Even though most studies focus on parent-to-child relations, child psychopathology can prospectively predict parent psychopathology. For example, child depression in middle childhood predicts parent depression two years later (Wesseldijk et al., 2018). Silverman and colleagues (Silverman et al., 2021) found that cognitive behavioral therapy (CBT) targeted at child anxiety was associated with post-treatment parental anxiety even though parental anxiety was not the target of any of the study conditions. These bidirectional relations are evident even when one can account for shared risk via genetically informed models (Cioffi et al., 2021). Most often, associations between parent and child are most robust when assessed as a chronic exposure (Borelli et al., 2015; Grabow et al., 2017; Pemberton et al., 2010) versus focusing on a single exposure (Hails et al., 2018). The importance of chronic exposure suggests that smaller time intervals within daily interactions build on each other to generate larger-scale impacts on development and psychopathology risk.
Although our review is not focused on global constructs of parenting as traditionally defined by the developmental psychology literature [e.g., parenting styles, attachment (Baumrind, 1971; Belsky, 1984)], previous research has found associations between parent-linked behaviors and child well-being. Indeed, anxious parents have distinct parenting profiles that impact socioemotional development (Kiel & Buss, 2011, 2014; Kiel & Hummel, 2017; Kiel et al., 2016). For example, anxious parents can be over-protective (Degnan et al., 2008; Hastings et al., 2019; Rubin et al., 1999) or over-solicitous (Buss & Kiel, 2011; Kiel & Buss, 2012; Kiel et al., 2016) in the face of actual or potential child distress.
While these behaviors are motivated by a desire to minimize negative experiences for their child, they can inadvertently prevent children from working to overcome initial anxious tendencies. Indeed, the everyday activities of school and play are often sufficient to ameliorate anxious tendencies in young children (Almas et al., 2011; Phillips et al., 2011; Rudasill & Rimm-Kaufman, 2009). Over-protective parenting behaviors may result in broad alterations to the biological and cognitive functioning of their children (Brooker et al., 2011, 2015; Torpey et al., 2013). Recent work suggests that children show preferential fear-learning when observing a parent versus an unfamiliar adult and this effect is potentiated if the parent is anxious (Silvers et al., 2020). Thus, children at familial risk for anxiety may be exposed to more anxiety-linked behaviors that they, in turn, are more likely to internalize then express.
Extant studies largely examine intergenerational transmission by capturing aggregate measures of parent and child traits or behaviors averaged over set time windows (Anaya & Pérez-Edgar, 2019; Belsky et al., 2012; Brooker & Buss, 2014; Guyer et al., 2015; Hastings et al., 2019; Kiel & Buss, 2011, 2014; Kiff et al., 2011). Although this approach has created a robust knowledge base, parent-to-child transmission is not fully captured by examining child or parental traits and behaviors in isolation. Rather than focus on broader parenting practices or behaviors, many of which have been found to only loosely correlate with child anxiety (McLeod et al., 2007), our model is intended to probe the underlying dynamic, granular, moment-to-moment interactions between parent and child. Repeated daily interactions with caregivers attune the child to parental expressions and regulation of fear and distress, which influence the child’s own responses to events (Leerkes et al., 2020). We have a relatively limited understanding of the relations between global risk and moment-to-moment interactions between parent and child.
Thus, the current review argues that we should focus not only on the bidirectional and dyadic relation between parent and child but also explicitly consider the time window of observation. That is, by examining moment-to-moment interactions, occurring within minutes or even seconds or milliseconds, we may be able to generate new insights into long-term developmental and clinical patterns. An emphasis on dynamic relations also reflects larger calls in the field to focus on active mechanisms that influence the full range of psychological and biobehavioral profiles. For example, the National Institute of Mental Health has called on researchers to examine development and the environment as “bidirectional influences” on the transdiagnostic processes of psychopathology through its Research Domain Criteria (RDoC) (Casey et al., 2014; Sanislow et al., 2010).
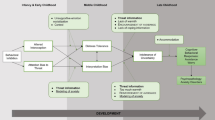
Better understanding the functional mechanisms by which parental traits are passed on to children can provide (1) novel markers of risk for socioemotional difficulties; (2) specific targets for intervention; and (3) both behavioral and biological indices of treatment response. Specifically, by functional mechanism, we mean one of the critical mechanistic pathways by which parent psychopathology can lead to child psychopathology. Ideally, this mechanism would be capable of modification through targeted intervention or prevention. The current review highlights the importance of incorporating novel methods and analytics for capturing dynamic relations. Here, we present a conceptual developmental model (Fig. 1) focusing on dyadic social dynamics as a key conduit in parent-to-child transmission of anxiety. Dyadic social dynamics capture the moment-to-moment interactions between parent and child that occur on a daily basis, as well as the parental behavioral and socioemotional patterns that children observe.

Conceptual model of trajectory of parent-to-child anxiety transmission. Across development parent anxiety influences the development of child anxiety symptoms through dyadic social dynamics. Over time, emerging child anxiety symptoms can also influence these dyadic interactions. This process can be moderated by a number of individual or contextual factors, illustrated here with child fearful temperament
We note that, although dyadic social dynamics are discussed here in the context of the parent–child relationship, they are present between any two interacting individuals, which distinguishes these fine-grained, dyadic behaviors from general parenting practices. The impact of these dynamics may of course be potentiated in the parent–child relationship in that it uniquely combines long term and repeated exposure and the pivotal role parents play in shaping a child’s environment and daily activities. In shaping the developmental trajectory from familial risk to actual symptoms, dyadic processes act on markers of risk that are evident prior to, and in the absence of, any eventual disorder onset. We will delineate two cases of dyadic social dynamics.
First, we will review dyadic synchrony or the moment-to-moment coordination of behavioral, neural, and physiological processes between parent and child. We will discuss how novel dynamic measures can be leveraged to capture patterns of concordance. Second, we will discuss how overt emotion modeling of distress is observed and internalized by children and later reflected in their own behavior. In doing so we discuss the task and analytic considerations that should be addressed in order to apply the model to research. In conclusion, we highlight future directions, which include a focus on contextual or individual factors that can impact dyadic processes as well as the promise of leveraging dyadic processes for treatment and prevention.
Dyadic Social Dynamics as a Mechanism for Anxiety Transmission
Parental anxiety markedly increases anxiety risk in children. Indeed, up to 33% of children with anxious parents will present with a corresponding psychiatric diagnosis (OR = 1.92) (Johnson et al., 2001). Our ability to target underlying biological transmission mechanisms, such as genetic predisposition or fetal programming of the stress response, is limited and generally untested. Consequently, there exists a robust literature examining how parental behavior can exacerbate or ameliorate genetic and prenatal risk. In particular, over-protective behavior and psychological control tend to restrict children’s behavior and may actively encourage dependency (Hastings et al., 2019), which has been associated with social withdrawal and anxiety both concurrently (Chronis-Tuscano et al., 2018; Rubin et al., 2018) and prospectively (Coplan et al., 2008; Degnan et al., 2008) from toddlerhood through early elementary school. In addition, children at temperamental risk for psychopathology appear particularly sensitive to parenting behaviors, as noted below (Hastings et al., 2019).
Parental behavior, in turn, is a core component of adaptive calibration models (ACM) which examine how the environment “gets under the skin” to shape psychobiological development (Del Giudice et al., 2011; Ellis & Boyce, 2008; Ellis, Boyce, Belsky, Bakermans-Kranenburg, & Van IJzendoorn, 2011). Prior work has focused on broad measures (e.g., over-protection) averaged over set time windows. These measures can provide a stable and robust metric for analysis, which can then be supplemented by measures that capture dynamic interactions in the moment (Shih, Quiñones‐Camacho, Karan, & Davis, 2019). It is in these fine-grained interactions that parents may model distinct emotional and stress responses linked to anxiety. Focusing on the temporal context of parenting (Cole, Lougheed, & Ram, 2018; Davis, Brooker, & Kahle, 2020) provides novel ways of assessing moment-to-moment synchrony during interactions, the spill-over effects of observed emotionally salient events, and the long-term arc of change in anxiety-relevant mechanisms and outcomes. Thus, by moving across different time windows researchers can ask interconnected questions, such as (1) Concurrently, how do patterns of dyadic social dynamics vary across parent–child pairs? (2) Across contexts, to what extent does variation in dyadic patterns help predict or explain anxiety risk? (3) Over time, can we predict socioemotional profiles and anxiety risk from earlier patterns captured during dynamic dyadic interactions?
Dyadic Synchrony
Definition of Dyadic Synchrony
The link between partners engaged in a dyadic interaction reflects a contingent relation between the two members of the dyad (Davis et al., 2020; Davis et al., 2018). Synchrony, operationalized behaviorally as the ‘temporal co-ordination of discrete micro-level signals between dyadic partners’ (Azhari et al., 2019; Bornstein, 2013), is the most basic unit of analysis via which partners are linked. The specific time scale of the interaction of interest often varies with the measure of interest. For example, studies of cardiac synchrony can have thousands of data points embedded within an interaction only a few minutes long. Studies coding for facial expressions, in contrast, will often have fewer discrete data points as they oscillate in slower frequencies (e.g., Fig. 2). However, synchrony measures are computationally intense, so most studies rely on relatively short epochs on the order of 5 to 10 s embedded within interactions that last minutes. One counter example can be found in studies of diurnal cortisol synchrony (e.g., Laurent, Sbrilli, Dawson, Finnegan, & Ramdas‐Neal, 2021) or parent–child autonomic synchrony over the course of a day (Smith et al., 2019).

Example of emotion synchrony in a mother–infant dyad engaged in free play. The solid lines reflect the average emotion level for the mother (green) and infant (orange). The dashed lines capture fluctuations in emotion for each dyadic partner. The temporal linkages between fluctuations are captured in analytic synchrony scores (Color figure online)
Within these epochs, synchronized dyads match, reciprocate, and jointly expand upon one another’s actions (e.g., vocalizations, motor activity, and emotions). Synchronized dyads may also exhibit parallel and coordinated autonomic activity, likely reflecting co-created arousal states (Davis et al., 2017; Davis et al., 2018). Synchrony is an emergent property of the dyad’s shared experience, not simply reflecting shared space, proximity, or exposure to an event or stimulus (Fishburn et al., 2018). Higher-order social processes, such as engagement and mutual attunement, further potentiate this synchronization (Wass et al., 2020). This basic synchrony, in turn, can serve as the foundation for other attachment-related dyadic constructs, such as co-regulation (Cole & Hollenstein, 2018; Hollenstein, 2013; Lunkenheimer et al., 2011). In following, synchronized signals may provide the latent interpersonal scaffolding onto which more deliberate socialization and regulatory processes are built.
Thus, fine-level measures (e.g., heart beats) are embedded within epochs that can then be used to understand developmental change over longer-time windows, including on the order of years. Dyadic synchrony may also enable the transmission of adaptive or maladaptive, behavioral strategies for addressing uncertainty in the environment by attuning the child to the caregiving context. Thus, we argue that high and low synchrony are not inherently positive or negative indicators of adaptive development, as specific patterns of covariation in biobehavioral systems between parents and children may differentially associate with familial risk for specific disorders (Lunkenheimer, Tiberio, Skoranski, Buss, & Cole, 2018; Smith et al., 2019; Wass, Clackson, & Leong, 2018).
Measurement of Dyadic Synchrony
Traditionally, dyadic synchrony, as defined in the developmental psychology literature, is measured behaviorally using second-by-second or frame-by-frame video coding schemes (Feldman, 2007; Feldman & Greenbaum, 1997; Feldman et al., 1996; Harrist & Waugh, 2002; Kellerman et al., 2020; Tronick, 1989; Tronick & Cohn, 1989). These involve careful hand coding by trained research assistants, often noting the vocalizations, eye contact, actions, or emotional expression of each member of the dyad independently and then examining timepoints in which these behaviors coordinate (Beebe et al., 2011; Granat et al., 2017; Moore & Calkins, 2004). This can include lagged time courses in which one member of the dyad might lead and another might follow.
Additionally, recent research has drawn attention to underlying biological components of dyadic synchrony. Dyadic synchrony of the autonomic nervous system has been measured via heart rate (Creaven et al., 2014; Suveg et al., 2016), respiratory sinus arrhythmia (Lunkenheimer et al., 2015; Ostlund et al., 2017), pre-ejection period (Helm et al., 2018), and salivary alpha amylase (Laurent et al., 2012), thought to reflect stress, emotional arousal, and engagement in interpersonal interactions. This method has made advances in the mother–child attachment literature from infancy through childhood (Field et al., 1989; Giuliano et al., 2015; Lunkenheimer et al., 2021), but also in intra-fetal mother–offspring dyads (Van Leeuwen et al., 2009). Dyadic synchrony in the hypothalamic–pituitary–adrenal (HPA) axis has been assessed via attunement in both cortisol reactivity to a stressor and diurnal cortisol rhythms (Laurent et al., 2021; Pratt et al., 2017), also reflecting dyadic indices of stress regulation.
Neural methods for the measurement of dyadic synchrony include both electroencephalography (EEG) and functional near-infrared spectroscopy (fNIRS). Using these methods, the time course of the neural signal is measured in each individual and then compared for moments in which the rise or fall of the neural signal of one individual predicts that of the other (Cui et al., 2012; Fishburn et al., 2018; Kinreich et al., 2017; Liu, Duan, Dai, Pelowski, & Zhu, 2021). EEG methods for measuring neural synchrony capitalize on the high temporal resolution of the method and the mobility of multiple rather than single devices between subjects. The latter has been particularly useful in ecologically valid contexts, such as schools (Bevilacqua et al., 2019; Dikker et al., 2017), musical collaboration (Lindenberger et al., 2009; Müller, Sänger, & Lindenberger, 2013) (i.e., a jazz band), or museums (Dikker et al., 2021). fNIRS, a method for measuring the hemodynamic response of blood oxygen in the brain (Ferrari & Quaresima, 2012) with higher spatial resolution relative to EEG, has been particularly useful for identifying specific neural regions that synchronize between parents and children in such cases as stressful interaction or competition (Quiñones‐Camacho et al., 2019; Reindl et al., 2018), language acquisition (Piazza, Cohen, Trach, & Lew-Williams, 2021), or developmental disorders (Kruppa et al., 2020; Quiñones-Camacho et al., 2021; Quiñones-Camacho et al., 2021; Su et al., 2020).
It is important to note the unique time scale of each biobehavioral system and consider theory regarding whether synchrony of that system is meaningful on the specified time scale. For example, neural activation may manifest on the order of milliseconds, whereas behavioral responses may play out across seconds or minutes. Therefore, methodological designs for the study of synchrony may differ depending on the biobehavioral system in question. More consideration of methodological differences and similarities across biobehavioral systems is needed to refine our understanding of the function of dyadic synchrony between parent and child (DePasquale, 2020; Helm et al., 2018).
Dyadic Synchrony in the Parent–Child Psychopathology Context
Parent–child dyadic synchrony is arguably most meaningful in the context of challenging or goal-oriented interactions and thus is typically studied in tasks involving challenging prompts, stimuli, or changing conditions designed to trigger behavioral changes of interest in the dyad. For example, in early parent–child interaction tasks, stimuli may involve experimental alterations to expected parent behavior (e.g., the Still Face Paradigm) (Moore & Calkins, 2004) or the goal of completing a difficult puzzle that requires parental scaffolding of children’s efforts (Hoyniak et al., 2021; Lunkenheimer et al., 2018; MacNeill, Fu, Buss, & Pérez-Edgar, in press). Using these paradigms, there is an emerging literature examining how a parent and child synchronize both behavioral and neural and physiological activities (Hasson et al., 2012). While research on parent–child neural synchrony is still scarce, the few published studies suggest that increased parent–child neural synchrony is linked with better emotion regulation (Reindl et al., 2018), less parenting stress (Azhari et al., 2019), and greater behavioral reciprocity (Nguyen et al., 2020), lending additional support for biological synchrony as a mechanism facilitating the transmission of behavioral strategies.
Dyadic synchrony is also modulated based on characteristics of the parent and child, such as parental harshness and psychopathology (Giuliano et al., 2015; Ham & Tronick, 2009; Laurent et al., 2011; Moore et al., 2009; Woody et al., 2016). For example, more maternal psychological aggression (e.g., intent to impose fear) (Lunkenheimer et al., 2018) and greater severity of child maltreatment (Lunkenheimer et al., 2018; Lunkenheimer et al., 2018) have been related to lower physiological synchrony. Synchrony has also been related to temperamental negativity and behavior problems in childhood (Lunkenheimer et al., 2018; Lunkenheimer et al., 2018; Lunkenheimer et al., 2018). Quiñones-Camacho and colleagues (Quiñones-Camacho, Hoyniak, Wakschlag, & Perlman, 2021) found that parent–child dyadic synchrony, measured neurally using fNIRS, positively predicted decreases in internalizing, but not externalizing, symptoms from 4 to 6 years of age. Thus, the mounting, multi-system evidence available underscores the association between synchrony and outcomes and supports future work testing the potential that synchrony acts as a key mechanism in the transmission and/or development of psychopathology in relation to underlying regulatory difficulties.
There is emerging evidence that synchrony may potentiate risk for poor outcomes, particularly in the context of anxiety (Beebe et al., 2011; Granat et al., 2017; Smith et al., 2019; Wass et al., 2020). Children in parent–child dyads that are highly synchronized behaviorally or physiologically exhibit better regulatory abilities and communication skills (Criss et al., 2003; Feldman et al., 1999; Harrist & Waugh, 2002; Im-Bolter et al., 2015; Kochanska et al., 2008; Suveg et al., 2016). However, low prefrontal synchrony is related to psychopathology risk (Quiñones-Camacho et al., 2021; Quiñones‐Camacho et al., 2019). One recent study (Smith et al., 2019) captured autonomic synchrony between caregivers and one-year-old throughout a typical day within the home. Anxious parents showed greater autonomic synchrony with their infants throughout the day than their less anxious counterparts (Smith et al., 2019). In particular, non-anxious parents showed elevated attunement with their infant only at moments of high arousal or need and then down-regulated in response to infant arousal. Anxious parents, in contrast, are reactive more often and at lower thresholds of infant arousal (Wass et al., 2020). Wass and colleagues suggest that anxious parents are ‘always on,’ while non-anxious parents take an approach of being there ‘when you need me.’
Additionally, studies with older children find greater behavioral synchrony among dyads with an anxious parent during table top play (Beebe et al., 2011; Granat et al., 2017), in contrast to lower synchrony with depressed (Granat et al., 2017) and maltreating (Lunkenheimer et al., 2018; Lunkenheimer et al., 2018) parents. Thus, while much of the literature has examined the positive effects of dyadic synchrony on child development (Feldman et al., 1999; Feldman et al., 2011; Lunkenheimer et al., 2018; Lunkenheimer et al., 2018; Lunkenheimer et al., 2011; Lunkenheimer et al., 2015; Lunkenheimer et al., 2018; Wass et al., 2020), there is growing evidence that dyadic synchrony may also act as a maladaptive influence (Beebe et al., 2011; Granat et al., 2017; Smith et al., 2019; Wass et al., 2020), particularly within the context of anxiety.
Emotion Modeling
Definition of Emotion Modeling
In the current review we are focused on two distinct forms of dyadic social dynamics: dyadic synchrony and emotion modeling. In the first, the dyad is linked by coupled moment-to-moment fluctuations that can be captured within multiple levels of analysis, including neural, physiological, and behavioral. In the second, we highlight a cascading dyadic relation that typically occurs across levels of analysis. That is, emotion modeling relies on the observation, and then internalization, of behavior, which is in turn reflected in the child’s cognitions, emotions, and behaviors (Denham, Bassett, & Wyatt, 2007). We suggest that emotion modeling is one functional mechanism underlying the relation between parental behavior and child socioemotional outcomes. It is possible that individual differences in dyadic synchrony may influence the strength and impact of emotion modeling (Feldman et al., 1999; Johnco et al., 2021; Pérez-Edgar, 2019). However, as we note below, this is one of many open questions in this literature.
The transgenerational continuity of stress reactivity (Hipwell et al., 2019) is partially driven by the inheritance of biological tendencies in reactivity profiles. The tendency to have an elevated stress response in both parent and child may be traced to an underlying shared biological predisposition (Bartels, Van den Berg, Sluyter, Boomsma, & de Geus, 2003). However, these tendencies can then be potentiated and triggered by the child directly observing the caregiver’s emotional and behavioral response to stress (Laurent, 2014; Silvers et al., 2020). Most learning takes place in social settings, as the brain dynamically adapts to the behavior of others (Wass et al., 2020). Modeling is evident from the first months of life as even infants will mimic emotional displays evident around them. For example, increased autonomic reactivity in a parent during an emotion exposure task is then reflected in increased negative affect in infants (Waters et al., 2014, 2017). This process of spreading or shared responses to the environment is often labeled an ‘emotional contagion’ effect that is evident even in early infancy (Geangu et al., 2010). Of note, this label may overstate the connection by implying the direct transmission of a shared emotional response in the moment (Ruffman et al., 2017).
However, the clear link between emotion and behavior in one individual and the subsequent shift in the emotion and behavior of another do point to the fact that we are attuned to perceptual and behavioral signals from others even before we have the ability to reflect more complex cognitive or affective responses. Indeed, this attunement appears to be an evolutionarily conserved trait as non-human primates exhibit referential gaze to caregivers and respond differentially to emotional messages based on both gaze behavior and avoidance of the novel objects (Russell et al., 1997). These summative effects then lead to observed variation in sociobehavioral profiles. In particular, observations create ‘ostensive cues’ that trigger a neural response, which, with repetition, may lead to dyadic attunement (Albert, Schwade, & Goldstein, 2018; Grossmann et al., 2008; Urakawa et al., 2015; Young et al., 2017), which is evident in the child’s reflection of modeled behavior. Building on associative and statistical learning, children come to create stable schematic expectations of their environment (e.g., threatening vs. safe) based on how the people around them react to daily life experiences. The child’s idiosyncratic socioemotional curriculum can facilitate fear or anxiety learning (Muris et al., 1996) and model how the child should respond.
Measurement of Emotion Modeling
Children likely take in instances of modeling through a number of channels. That is, caregivers and adults can convey information through verbalization, emotion expression, or direct action. The initial steps in studying this process is to capture both the presented signal and the subsequent capture by the child. On the first point, a number of emotion modeling studies have focused on behavioral observation of parental emotion expression (Camras, 2019). Behavioral coding schemes have focused on micro-analytic coding of facial affect (Chaplin et al., 2005) as well as broader more gestalt affective expressions, including body language (Lunkenheimer et al., 2020). Parental expressions often emerge spontaneously and are not necessarily directly aimed at the child. As such, their value as a modeling signal comes in the child’s equally spontaneous attention to the form and function of these expressions, linking the trigger, the expression, and the subsequent outcomes (Bayet & Nelson, 2019).
Often, the emotion modeling literature then looks to see if the captured parental signal of emotion is correlated with or predicts child emotional behavior (Burstein & Ginsburg, 2010; Eisenberg et al., 1998; Muris et al., 1996). This is a fundamental relation for establishing the impact of modeling. However, as in our discussion of dyadic synchrony, it skips over the functional mechanisms that could be linking parental modeling with child expression (Zhou et al., 2002). Here, we focus on two potential processes that might generate this link: selective attention to parental expression and the child’s biological response.
Interestingly, there are relatively less data specifically focused on selective attention to parental modeling (Aktar, Nimphy, Van Bockstaele, & Pérez-Edgar, in press). However, it may be that parents who frequently model negative emotion expression are providing the salient stimuli that feed into, and entrench, attention biases to threat (Aktar & Bögels, 2017). Indeed, there are emerging data suggesting a link between parental anxiety and attention biases in both infancy (Morales et al., 2017) and early childhood (Aktar et al., 2013; Aktar, Van Bockstaele, Pérez‐Edgar, Wiers, & Bögels, 2019).
The broader evidence base suggests the likely mechanistic pathways between parental modeling and child outcomes via attention. For example, cognitive theories of anxiety collectively suggest that anxiety is associated with attention bias toward threat (Van Bockstaele et al., 2014). Information processing perspectives (Crick & Dodge, 1994; Morales et al., 2016; Pérez-Edgar, Taber-Thomas, Auday, & Morales, 2014) further argue that attention bias to threat elicits a cascade of effects on downstream information processing and behavioral responses. These processes influence each other through a feedback loop, strengthening a biased mental database that perpetuates and reinforces anxious behaviors over time.
Involuntary orienting toward salient stimuli gradually tunes the visual system, resulting in biased attention, memory encoding, and behavioral enactment in response to previously perceived threats. Thus, attention bias to threat may be an automatic process produced from repeated pre-tuning of top-down attention selection (Todd et al., 2012). The cyclical relation between attention and socioemotional experiences, mediated through caregiving, may shape long-term anxiety trajectories. For example, temperamentally fearful children are most likely to exhibit maladaptive social withdrawal (Cole et al., 2016; Morales, et al., 2015; Morales et al., 2015; Pérez-Edgar et al., 2011; Pérez-Edgar et al., 2010; Pérez-Edgar et al., 2010) and anxiety (White et al., 2017) if they also exhibit an attention bias to threat.
As with dyadic synchrony, new technology has allowed for more ‘child-centered’ observation of the instances of modeling they experience. For example, recent work has used mobile eye-tracking (Pérez-Edgar et al., 2020) to move beyond the use of static, non-dynamic stimuli that do not fully reflect the rich information provided by one-on-one contingent social interactions (Fu & Pérez-Edgar, 2019). Mobile eye-tracking can provide the child’s point of view and supplement observed behavior captured by room-view cameras that may capture the major dynamics of an encounter but not the idiosyncratic input available to the child. One recent study used mobile eye-tracking to capture underlying tendencies in attention as adolescent girls present a speech in front of two judges, one positive and one critical (Woody et al., 2019). They found that the likelihood to visually focus on the critical judge correlated positively with depression symptoms. Patterns of visual attention were also associated with neural connectivity among regions associated with emotion regulation (Sequeira et al., 2021). Importantly, mobile eye-tracking can provide insight into the child’s processing of observed behavior even in the absence of any overt behavior on the part of the child. For example, in one laboratory task, six-year-olds rarely directly interacted with an unknown adult. However, mobile eye-tracking measures found that children who visually avoided the stranger, and had visual patterns less coupled with the stranger’s actions, demonstrated higher levels of internalizing problems (Gunther et al., in press).
Once attended to, the child’s initial response to parental emotion expressions may be evident in neural or physiological signals. One classic example can be seen in the visual cliff experiment (Sorce et al., 1985). In one variation, an infant is coaxed to cross the glass cliff by the mother, who is asked to present with either neutral, fearful, or encouraging affect. The infant is less likely to do so if the parent presents with a hesitant or fearful expression, and this hesitation is accompanied by changes in the infant’s cardiac response to the cliff (Campos et al., 1992; Klinnert, Campos, Sorce, Emde, & Svejda, 1983). It is important to point out that this initial physiological response need not mean that the infant is also experiencing an emotional response. That is, the infant can attend to the parent’s signal, internalize its meaning (e.g., the cliff may be dangerous), and act accordingly (LoBue & Adolph, 2019) without experiencing an emotional response. From this perspective, the physiological or behavioral response is not in and of itself evidence for emotion modeling. Rather, we would need downstream evidence of a fear response, potentially mediated by the cardiac response, to reflect our current conceptualization of emotion modeling.
Observational learning and modeling may also be reflected in a neural response. At a basic level, the motor cortex responds to an action performed by another person (Kerr et al., 2019). This response is part of a larger network, often incorporating the anterior cingulate cortex (ACC), anterior insula, and amygdala (Tramacere & Ferrari, 2016). Of relevance here, higher levels of maternal negative emotion have been linked to greater neural activation in brain regions implicated in detecting (e.g., amygdala, anterior insula) and regulating (e.g., ACC, vlPFC) emotions (Chaplin et al., 2019; Turpyn et al., 2018).
One drawback to imaging studies is that the emotion modeling stimuli are often static pictures or pre-recorded videos in order to account for testing requirements. However, recent work with fNIRS technology suggests that we can couple neural responses in active interactions, much as we have traditionally done with cardiac measures. Future work can use these new technologies to capture the three-needed components of our emotion modeling chain: (1) parental emotional expression, (2) a shift in the child’s attention and/or biological response, and (3) a subsequent shift in the child’s own emotional expression or behavior. This chain allows researchers to extend the general argument that parental expressions of emotion have a direct impact on a child’s emotional arousal and learning about emotions (Eisenberg et al., 1998) by examining parents’ active emotional signaling and the dynamic cascade of responses that may follow (Hajal & Paley, 2020).
Emotion Modeling in the Parent–Child Psychopathology Context
Within the emotion development literature, emotion modeling is often approached as a core component of the parental socialization of emotion, along with overt reactions to the child’s expressive behavior, explicit discussions of emotion, and managing children’s exposure to emotion-inducing situations (Camras, 2019; Camras, Shuster, & Fraumeni, 2014). Parental socialization encompasses a mix of behaviors that are at times overt or covert and can be deliberate or unconscious. Over time, they help shape individual patterns of expression, familial patterns of expressiveness, and contribute to social or cultural profiles of emotional experience and expression (Camras et al., 2014; Leerkes et al., 2020). For example, Holodynski and Friedlmeier (2006) suggest that caregivers shape an infants’ undifferentiated negative expressions (e.g., crying) to reflect the response deemed appropriate to the specific cue or trigger. That is, parents themselves will display the more specific discrete expression (e.g., anger or sadness) to model the expected familial or cultural response. One meta-analytic study (Halberstadt & Eaton, 2002) found that expressivity by family members was significantly related to children’s own emotional expressiveness.
Emotion modeling by parents spans a wide range of emotions, including anger (Dollar & Calkins, 2019), sadness (Denham & Kochanoff, 2002), fear (LoBue, Kim, & Delgado, 2019), disgust (Gerull & Rapee, 2002; Shutts et al., 2013), and happiness (Messinger et al., 2019). Through these behaviors, parents can communicate common behavioral tendencies associated with daily events, as well as the personal significance of an eliciting event (Denham, 2019). Importantly, this process influences and shapes the regulation of emotion and socioemotional competence (Eisenberg et al., 2001; Valiente et al., 2004). Intense and frequent expression of anger within parent–child interactions is associated with lowered abilities to appropriately regulate anger and aggressive behaviors (Rubin et al., 2003; Smeekens et al., 2007). However, exposure to well-regulated negative emotion is positively related to components of emotional competence (Garner et al., 1994).
In the specific case of anxiety, anxious parents tend to express higher levels of behavioral and linguistic stress markers in social situations or unfamiliar contexts (Aktar et al., 2013; Murray et al., 2008). In addition, anxious parents are more likely to exhibit interpretation biases that lead them to negatively frame daily events (Creswell, Cooper, & Murray, 2010; Creswell et al., 2011). Parental-negative interpretations are often not confined to their own environment, but also extended to their child’s environment and daily activities (Lester et al., 2009). As a result, they are also more likely to spontaneously communicate threat information to their children regarding potentially stressful situations (Murray et al., 2014; Percy et al., 2016). Children receiving threat information are, in turn, more likely to endorse that the targeted person or context is threatening when directly asked (Aktar et al., in press). This process has been observed naturalistically, but also through laboratory induction. One study (De Rosnay et al., 2006) trained mothers to interact with a stranger in either a normative or socially anxious manner. Infants displayed more hesitancy to interact with the stranger after observing the socially anxious interaction of their mother. The potency of this information can increase when combined with individual difference factors, like fearful temperament (LoBue et al., 2019).
Child observation of anxiety-tinged parenting behavior can lead to the subsequent experience of anxiety on the part of the child (Reynolds, Askew, & Field, 2018). For example, anxious child behavior in a modified speech task has been associated with the later emergence of social withdrawal in middle childhood and adolescence (Degnan et al., 2011). It may be that this anxious behavior reflects not only the child’s individual traits (e.g., temperament) but also their internalization of observed distress. Indeed, children who observed their caregiver display anxiety during an oral spelling test endorsed higher anxiety levels, anxious cognitions, and desired avoidance of the spelling test versus children who observed low levels of anxious behavior (Burstein & Ginsburg, 2010).
By the same token, repeated observations of non-anxious responses to mild stressors are linked to decreased anxiety in at-risk children. For example, behaviorally inhibited toddlers who experience non-parental child care are less likely to display shyness and social reticence as preschoolers, relative to children in familial care (N. A. Fox et al., 2001). Daily close interactions with an anxious individual, particularly in the absence of countervailing evidence may inadvertently convey a general expectation of negative social evaluations (Bögels et al., 2011; Ollendick & Hirshfeld-Becker, 2002). Compounding this effect, anxious parents may provide fewer social outlets for their children, reflecting their own concerns with social interactions (Ollendick & Hirshfeld-Becker, 2002; Spence & Rapee, 2016). The relatively strong countervailing effect of out-of-home care (Degnan et al., 2008; Phillips et al., 2011) also reflects clinical insights into the use of gradual, but repeated exposure, in cases of social anxiety (Kendall et al., 1997; Radtke, Strege, & Ollendick, 2020).
Outcome Measurement: Early Markers Preceding Symptom Onset
Our model (Fig. 1) proposes that researcher can capture the developmental progression to anxiety by examining intermediate markers that can precede an increase in overt or tractable symptoms. The developmental literature has carefully detailed endogenous factors that are associated with individual patterns of anxiety development (Fox et al., 2006; Hane et al., 2008), including EEG-based markers of regulation, ERP-based metrics of cognitive control, and patterns of attention bias to threat evident in eye-tracking measures (Briggs‐Gowan et al., 2015; Brooker et al., 2014; LoBue et al., 2014; Morales et al., 2015; Morales et al., 2015; Pérez-Edgar et al., 2013; Pérez-Edgar, et al., 2010; Pérez-Edgar et al., 2010). To date, however, we do not know how these factors intersect with emerging patterns of anxiety, as influenced by dyadic social dynamics. These biological and cognitive markers of risk in early childhood can be examined across RDoC units of analysis (Casey et al., 2014). Further, they can serve as leading indicators of anxiety progression, given that young children may not yet show elevated symptom levels and provide insight into associated functional mechanisms of risk.
First, the field has often employed neural measures, specifically using EEG, to mark anxiety risk. For example, delta–beta coupling is the moment-to-moment correlation between EEG power in the delta (1 to 3 Hz) and beta (13 to 20 Hz) bands, reflecting the cross-talk between subcortical and higher-order cortical brain networks (Knyazev et al., 2006). Top-down processes (marked by prefrontal cortex activation and beta power) are thought to dampen bottom-up stimuli (marked by limbic activity and delta power) (Knyazev et al., 2006; van Peer et al., 2008). Decoupling between delta and beta power may indicate regulation difficulties, while positive coupling reflects active regulatory processes. However, some studies have reported strong, positive delta–beta coupling in high-anxiety groups (Knyazev, 2011), suggesting that enhanced synchrony can be indicative of “overcontrol” processes induced by anxious states. Stronger delta–beta coupling has also been observed in children with dysregulated fearful temperament (Phelps et al., 2016).
Additionally, EEG signals time-locked to individual task trials can be aggregared to creat event-related potentials (ERPs), which can both capture current processing on the order of milliseconds and act as a predictive indicator of later socioemotional functioning (Hajcak et al., 2019). The N2 component, in particular, is a negative deflection that peaks between 200 and 350 ms after stimulus onset and reflects response inhibition, attention shifting and focusing, and conflict detection (Luck et al., 2000). Thus, the N2 may reflect the extent to which children monitor their environment, take in information, and shift subsequent processing. One study found that six-year-olds high in fearful temperament showed elevated levels of anxiety symptoms only if they also exhibited a small N2 during a flanker task (Morales, 2017).
Analytic Considerations
Our approach to examining moment-to-moment dyadic interaction, often in a longitudinal context, requires careful application of complex statistical methodology. Calculation of synchrony as a dyadic index should employ measures of similarity, such as Pearson product-moment correlations between two sets of scores, covariances, or intraclass correlations where larger values imply greater similarity (Kenny, Kashy, & Cook, 2006). Synchrony indexes are helpful when measuring similarity (i.e., high and low synchrony) across a set of behavioral, physiological, or neural measures to understand contingent relations between members of a dyad. Modeling moment-to-moment interactions that emerge into developmental patterns of risk require longitudinal data that are classified as stable processes or developmental processes (Jongerling et al., 2015; McNeish & Hamaker, 2019).
Emergent Patterns in Dyadic Synchrony
Stable processes allow for the capture of dyadic synchrony within moment-to-moment interactions. These intensive longitudinal data (ILD) structures typically include ten or more measurement occasions that are spaced closer together (e.g., seconds, hours, days) with a focus on instantaneous change (McNeish & Hamaker, 2019). Mean-level changes are typically ignored given that such processes are mean-reverting and stable within an observation window (e.g., Fig. 2). Rather, variability of the outcome (i.e., dyadic synchrony) becomes the focus of the analysis, which includes understanding the relative influence of covariates in explaining within-person variability. A candidate model to analyze stable processes is the two-level dynamic structural equation model (DSEM) appropriate for N > 1 analysis of ILD structures (Asparouhov & Muthén, 2020; Asparouhov et al., 2018; McNeish & Hamaker, 2019). Two-level DSEM integrates different modeling approaches (i.e., time-series analysis, multilevel modeling, and structural equation modeling) that specifies a within-dyad (fixed effects) level that meets the stationarity assumption (i.e., mean, variance, and autocorrelations do not systematically change over time) and a between-dyad (random effects) level.
Here, dyadic synchrony can be used as the repeated measures outcome assumed to be mean-reverting, where covariates such as child temperament can be included to examine how or why synchrony patterns might vary. Given that synchrony reflects an emergent property of the parent–child dyad’s shared experience, one way to model heterogeneity in these patterns is to employ finite mixture modeling. For example, DSEM mixture models allow estimation of a categorical latent class or continuous latent profile where dyadic synchrony patterns (within-dyad) and the relative influence of covariates (between-dyad) are now dependent on this latent classification or profile. These emergent classes or profiles can then be employed in subsequent examinations of developmental processes.
Long-Term Anxiety Trajectories of Dyadic Synchrony Patterns
Developmental processes, unlike stable processes that focus on variability around a mean, focus on the amount of change in the means of the outcome variable over time. Although still longitudinal in nature, these outcomes are measured in fewer occasions (e.g., 3–10) that occur across months or years. Whereas covariates in stable processes predict fluctuations in each moment-to-moment measurement, covariates in developmental processes are considered in how they explain between-dyad variability or change in growth curves. Therefore, emergent patterns (e.g., latent profiles) derived from the analysis of ILD structures can further be employed in the examination of developmental trajectories that influence child anxiety symptoms as a result of familial risk factors. For example, estimation of multiple DSEM mixture models from annual parent–child lab visits across ages 3, 4, 5, and 6 years can be examined using latent growth curve modeling. The shape (e.g., linear, quadratic, cubic) of such developmental processes can also be hypothesized based on existing literature and used to inform macro-level data collection occasions. Inclusion of auxiliary variables such as distal outcomes (e.g., child anxiety symptoms) can also be included and precited based on the resultant growth patterns.
Capturing Emotion Modeling in Parent–Child Dyads
Given the availability of child-centered data capture technology (e.g., mobile eye-tracking, heart rate) to measure intensive longitudinal data, differential equation modeling (DEM) could serve as a viable analytic tool. According to Ferrer and Steele (2012), DEMs are useful when considering members of a dyad as an interdependent system where changes result from individual level and/or system-level factors. Each interdependent system is modeled as two differential equations (see Ferrer & Steele, 2012, p. 114) that represent the rate of change in behaviors/emotion for members of a dyad (e.g., parent and child). Framed within the hypothesis that children’s phenotypic expression of anxiety results from direct modeling of caregiver response to stress, parameter estimates for DEMs can be modeled to show that changes in a child’s behavior/emotion over time are a function of individual factors (e.g., child-related goals) and system-level factors (e.g., caregiver stress response). Variations in the basic model can be employed, resulting in four different predicted patterns of dyadic functioning: approach (child behavior/emotion mirrors that of the caregiver), independent (parent and child behavior/emotion are separate from each other), contrarian (parent and child behavior/emotion repel each other), and unregulated (cyclical oscillations in child and parent behavior/emotion that reach equilibrium or instability).
Conclusions, Future Directions, and Treatment Implications
Our conceptual model of parent-to-child anxiety transmission (Fig. 1) is designed to develop innovative research questions and test tractable hypotheses underlying the mechanisms by which parent-to-child anxiety transmission occurs within early childhood. The current approach builds on decades of work characterizing the transactional nature of the parent–child relationship with respect to emergence of psychopathology over time (Sameroff, 1975; Yirmiya, Motsan, Kanat‐Maymon, & Feldman, 2021). As described above, we suggest that parent anxiety is behaviorally transmitted to the child by a process of dyadic social dynamics, leading to maladaptive cognitive and emotion processing. The approach of examining variation across dyadic social dynamics, using multi-modal assessments, especially when applied to a longitudinal context, will allow us to capture trajectories of behavior change over time linked to in-the-moment dynamics (Casey et al., 2014). Importantly, our model presents a flexible empirical strategy that can be applied to developmental and clinically relevant questions across multiple domains of interest. Understanding transmission through dyadic processes may extend to other disorders (e.g., depression, disruptive behavior) or other life stressors (e.g., poverty, family interpersonal stress), to help explain both how emotional dysfunction and adaptive emotional responses occur in familial relationships.
As is evident in the review, there are a number of open questions regarding the form and impact of dyadic social dynamics in the transmission of anxiety. There are, among others, two broad areas of work needed in order to advance this research. First, when and for whom does variation in the context and content of dyadic social dynamics impact anxiety trajectories? Second, once these relations are isolated, do direct perturbations to the systems shift patterns of anxiety? If yes, this would support the premise that dyadic social dynamics can serve as an active mechanism in intergenerational transmission. On the first question, there are a number of strong initial candidates for potential moderators (or mediators) of dyadic effects. Based on the broader anxiety literature, this includes child or parent sex (Bögels & Phares, 2008), match in parent and child sex, co-parent behavior, the specific form of parental psychopathology, patterns of co-morbidity, and individual differences in stress reactivity.
For example, it is unlikely that all children show the same levels of dyadic synchrony nor are as receptive to instances of emotion modeling. In particular, children sensitive to environmental input are more likely to carry the influence of dyadic process, for better and for worse. From the early literature, there is growing evidence that temperamental differences in socioemotional reactivity and regulation are associated with variation in synchrony and modeling. Fearful temperament is the strongest individual difference predictor of anxiety (Chronis-Tuscano et al., 2009; Clauss & Blackford, 2012; Fox et al., 2015; Fox, Snidman, et al., 2015; Hayward et al., 1998; Prior et al., 2000; Schwartz et al., 1999). Temperamentally at-risk children have a three- to fourfold increased likelihood for developing social anxiety (Clauss & Blackford, 2012). The majority of children with extreme temperament are not clinically anxious. Instead, they are more likely to display elevated levels of shyness and social withdrawal and may have difficulty establishing and maintaining social relationships with peers (Buss et al., 2013; Degnan & Fox, 2007; Fox et al., 2005, 2008).
Over-controlling and over-solicitous parenting behavior potentiate temperamental risk (Kiel & Kalomiris, 2015; Kiel et al., 2016), reflecting a complex relationship between parental behaviors and child anxiety risk (Kiel et al., 2021; Maag et al., 2021). These broad patterns of parental behavior are also evident in moment-to-moment interactions. For example, during a parent–child puzzle task, parents who had higher levels of anxiety spent more time in parent-focused/controlling behaviors with their children, which increased with levels of child fearful temperament (MacNeill et al., in press). Another study, in turn, found that parental anxiety is associated with higher levels of synchrony between parent and child in the same task, providing the underlying conduit for the observed relations (Quiñones-Camacho et al., 2021). Coupled with the fact that temperamentally fearful children display greater biological sensitivity to context (Hastings et al., 2019), attunement with parents may play an outsized role in shaping anxiety trajectories (Muris et al., 1996; Reynolds et al., 2018).
There is, of course, also the open question of whether variation in dyadic synchrony is associated with the impact of emotion modeling within the dyad. To date, we know of no empirical studies that have directly addressed this question. However, one could presume that higher levels of synchrony in parent–child dyads (e.g., neural or behavioral) may be associated with the child being particularly attuned to fluctuations in the parent’s expressed emotion, which could precipitate instances of direct modeling. This relation is unlikely to be simple or linear. For example, in an illustrative study with 10-year-olds, levels of behavioral synchrony between child–child dyads varied across structured and unstructured tasks, reflecting the specific in-the-moment goals for an interaction (Anaya, Vallorani, & Pérez-Edgar, 2021a). In addition, mismatches (e.g., one child positive and the other child negative) in delta–beta coupling, a neural marker for emotion regulation (Anaya et al., 2021b), impacted patterns of behavioral synchrony. In particular, negative coupling in the temperamentally fearful child was associated with less synchrony, perhaps reflecting underregulated cortical–subcortical interactions that may interfere with the ability to get in sync with an unfamiliar partner.
We emphasize that our developmental model has the potential to guide novel treatment approaches aimed at intervening on dyadically linked biobehavioral mechanisms before symptom onset. For example, interventions often target children after the emergence of functional impairment. The work described in this review highlights the translation of the parent–child dyad as an intervention point that can counteract evident (e.g., genetic, parenting)-risk markers. For example, future research pinpointing a deficit in dyadic social dynamics that develops along the trajectory to anxiety could be intervened upon using Parent–Child Interaction Therapy (PCIT). PCIT is the most widely used treatment to improve child mental health in early childhood (Herschell et al., 2002). Based on both Attachment Theory and Social Learning Theory, this intervention technique was designed to improve child disruptive behavior through increased attunement during parent–child interaction. PCIT has been adapted to treat anxiety (Puliafico et al., 2012) and depression (Lenze et al., 2011), among other disorders. For example, testing of our model could indicate an increase of dyadic synchrony during moments of increased tension with overt anxious behaviors from the parent (e.g., nervous movement, hovering), with or without a child moderator (e.g., fearful temperament), leading to a lagged response of anxious behaviors in the child (i.e., modeling). As a result, we might use PCIT to teach the parent how to limit their own anxious behaviors. This approach, of course, would only be possible through dyadic approaches to both research and treatment. Thus, in adapting our developmental model to potential treatment implications, we propose that PCIT, and other techniques focused on parent–child interaction, could be modified to treat children who are exposed to increased parental anxiety, especially those with highly anxious temperament, before the onset of their own symptoms. This would allow for early intervention and potential prevention prior to symptom onset along the psychopathology trajectory.
Change history
09 March 2022
A Correction to this paper has been published: https://doi.org/10.1007/s10567-022-00396-2
References
Aktar, E., & Bögels, S. M. (2017). Exposure to parents’ negative emotions as a developmental pathway to the family aggregation of depression and anxiety in the first year of life. Clinical Child and Family Psychology Review, 20(4), 369–390.
Aktar, E., Majdandžić, M., de Vente, W., & Bögels, S. M. (2013). The interplay between expressed parental anxiety and infant behavioural inhibition predicts infant avoidance in a social referencing paradigm. Journal of Child Psychology and Psychiatry, 54(2), 144–156.
Aktar, E., Nimphy, C. A., Van Bockstaele, B., & Pérez-Edgar, K. (in press). The social learning of threat and safety in the family: Parent-to-child transmission of stranger anxiety via verbal information. Developmental Psychobiology.
Aktar, E., Van Bockstaele, B., Pérez-Edgar, K., Wiers, R. W., & Bögels, S. M. (2019). Intergenerational transmission of attentional bias and anxiety. Developmental Science, 22(3), e12772.
Albert, R. R., Schwade, J. A., & Goldstein, M. H. (2018). The social functions of babbling: acoustic and contextual characteristics that facilitate maternal responsiveness. Developmental Science, 21(5), e12641.
Almas, A. N., Phillips, D. A., Henderson, H. A., Hane, A. A., Degnan, K. A., Moas, O. L., & Fox, N. A. (2011). The relations between infant negative reactivity, non-maternal childcare, and children’s interactions with familiar and unfamiliar peers. Social Development, 20, 718–740. https://doi.org/10.1111/j.1467-9507.2011.00605.x
Anaya, B., & Pérez-Edgar, K. (2019). Personality development in the context of temperament and parenting dynamics. New Ideas in Psychology, 53, 37–46.
Anaya, B., Vallorani, A., & Pérez-Edgar, K. (2021a). Dyadic behavioral synchrony between behaviorally inhibited and non-inhibited peers is associated with concordance in EEG frontal Alpha asymmetry and Delta-Beta coupling. Biological Psychology, 1, 108018.
Anaya, B., Vallorani, A., & Pérez-Edgar, K. (2021b). Individual dynamics of delta-beta synchrony: Using a multilevel framework to examine trait- and state-level differences in relation to Behavioral Inhibition and social anxiety. Journal of Child Psychology & Psychiatry. https://doi.org/10.1111/jcpp.13319
Askew, C., & Field, A. P. (2008). The vicarious learning pathway to fear 40 years on. Clinical Psychology Review, 28(7), 1249–1265.
Asparouhov, T., Hamaker, E. L., & Muthén, B. (2018). Dynamic structural equation models. Structural Equation Modeling, 25(3), 359–388.
Asparouhov, T., & Muthén, B. (2020). Comparison of models for the analysis of intensive longitudinal data. Structural Equation Modeling, 27(2), 275–297.
Azhari, A., Leck, W., Gabrieli, G., Bizzego, A., Rigo, P., Setoh, P., … & Esposito, G. (2019). Parenting stress undermines mother-child brain-to-brain synchrony: A hyperscanning study. Scientific Reports, 9(1), 1–9.
Bartels, M., Van den Berg, M., Sluyter, F., Boomsma, D., & de Geus, E. J. (2003). Heritability of cortisol levels: Review and simultaneous analysis of twin studies. Psychoneuroendocrinology, 28(2), 121–137.
Baumrind, D. (1971). Current patterns of parental authority. Developmental Psychology, 4(1), 1.
Bayet, L., & Nelson, C. A. (2019). The perception of facial emotion in typical and atypical development. In Handbook of emotional development (pp. 105–138): Springer.
Beebe, B., Steele, M., Jaffe, J., Buck, K. A., Chen, H., Cohen, P., … & Margolis, A. (2011). Maternal anxiety symptoms and mother–infant self-and interactive contingency. Infant Mental Health Journal, 32(2), 174–206.
Beesdo-Baum, K., & Knappe, S. (2012). Developmental epidemiology of anxiety disorders. Child and Adolescent Psychiatric Clinics, 21(3), 457–478.
Bell, R. Q. (1968). A reinterpretation of the direction of effects in studies of socialization. Psychological Review, 75, 81–95. https://doi.org/10.1037/h0025583
Belsky, J. (1984). The determinants of parenting: A process model. Child Development, 14, 83–96.
Belsky, J., Schlomer, G. L., & Ellis, B. J. (2012). Beyond cumulative risk: Distinguishing harshness and unpredictability as determinants of parenting and early life history strategy. Developmental Psychology, 48, 662–673.
Bevilacqua, D., Davidesco, I., Wan, L., Chaloner, K., Rowland, J., Ding, M., … & Dikker, S. (2019). Brain-to-brain synchrony and learning outcomes vary by student–teacher dynamics: evidence from a real-world classroom electroencephalography study. Journal of Cognitive Neuroscience, 31(3), 401–411. doi:https://doi.org/10.1162/jocn_a_01274
Biederman, J., Faraone, S. V., Hirshfeld-Becker, D. R., Friedman, D., Robin, J. A., & Rosenbaum, J. F. (2001). Patterns of psychopathology and dysfunction in high-risk children of parents with panic disorder and major depression. American Journal of Psychiatry, 158, 49–57. https://doi.org/10.1176/appi.ajp.158.1.49
Bögels, S., & Phares, V. (2008). Fathers’ role in the etiology, prevention and treatment of child anxiety: A review and new model. Clinical Psychology Review, 28(4), 539–558.
Bögels, S., Stevens, J., & Majdandžić, M. (2011). Parenting and social anxiety: Fathers’ versus mothers’ influence on their children’s anxiety in ambiguous social situations. Journal of Child Psychology and Psychiatry, 52(5), 599–606.
Borelli, J. L., Margolin, G., & Rasmussen, H. F. (2015). Parental overcontrol as a mechanism explaining the longitudinal association between parent and child anxiety. Journal of Child and Family Studies, 24(6), 1559–1574.
Bornstein, M. H. (2013). Mother-infant attunement: A multilevel approach via body, brain, and behavior. In The infant mind: Origins of the social brain (pp. 266–298).
Briggs-Gowan, M. J., Pollak, S. D., Grasso, D., Voss, J., Mian, N. D., Zobel, E.,… & Pine, D. S. (2015). Attention bias and anxiety in young children exposed to family violence. Journal of Child Psychology and Psychiatry, 56, 1194–1201.
Brooker, R. J., & Buss, K. A. (2014). Harsh parenting and fearfulness in toddlerhood interact to predict amplitudes of preschool error-related negativity. Developmental Cognitive Neuroscience, 9, 148–159.
Brooker, R. J., Neiderhiser, J. M., Ganiban, J. M., Leve, L. D., Shaw, D. S., & Reiss, D. (2014). Birth and adoptive parent anxiety symptoms moderate the link between infant attention control and internalizing problems in toddlerhood. Development and Psychopathology, 26(2), 347–359.
Brooker, R. J., Neiderhiser, J. M., Kiel, E. J., Leve, L. D., Shaw, D. S., & Reiss, D. (2011). The association between infants’ attention control and social inhibition is moderated by genetic and environmental risk for anxiety. Infancy, 16, 490–507.
Brooker, R. J., Neiderhiser, J. M., Leve, L. D., Shaw, D. S., Scaramella, L. V., & Reiss, D. (2015). Associations between infant negative affect and parent anxiety symptoms are bidirectional: Evidence from mothers and fathers. Frontiers in Psychology, 6, 1875.
Burstein, M., & Ginsburg, G. S. (2010). The effect of parental modeling of anxious behaviors and cognitions in school-aged children: An experimental pilot study. Behaviour Research and Therapy, 48(6), 506–515.
Buss, K. A., Davis, E. L., Kiel, E. J., Brooker, R. J., Beekman, C., & Early, M. C. (2013). Dysregulated fear predicts social wariness and social anxiety symptoms during kindergarten. Journal of Child Clinical and Adolescent Psychology, 42(5), 603–616. https://doi.org/10.1080/15374416.2013.769170
Buss, K. A., & Kiel, E. J. (2011). Do maternal protective behaviors alleviate toddlers’ fearful distress? International Journal of Behavioral Development, 35(2), 136–143.
Campos, J. J., Bertenthal, B. I., & Kermoian, R. (1992). Early experience and emotional development: The emergence of wariness of heights. Psychological Science, 3, 61–64.
Camras, L. A. (2019). Facial expressions across the life span. In Handbook of emotional development (pp. 83–103).
Camras, L. A., Shuster, M. M., & Fraumeni, B. R. (2014). Emotion socialization in the family with an emphasis on culture. In Children and emotion (Vol. 26, pp. 67–80): Karger Publishers.
Casey, B. J., Oliveri, M. E., & Insel, T. (2014). A neurodevelopmental perspective on the research domain criteria (RDoC) framework. Biological Psychiatry, 76(5), 350–353.
Chaplin, T. M., Cole, P. M., & Zahn-Waxler, C. (2005). Parental socialization of emotion expression: Gender differences and relations to child adjustment. Emotion, 5, 80.
Chaplin, T. M., Poon, J. A., Thompson, J. C., Hansen, A., Dziura, S. L., Turpyn, C. C., … & Ansell, E. B. (2019). Sex‐differentiated associations among negative parenting, emotion‐related brain function, and adolescent substance use and psychopathology symptoms. Social Development, 28(3), 637–656.
Chronis-Tuscano, A., Danko, C. M., Rubin, K. H., Coplan, R. J., & Novick, D. R. (2018). Future directions for research on early intervention for young children at risk for social anxiety. Journal of Clinical Child and Adolescent Psychology, 47, 655–667.
Chronis-Tuscano, A., Degnan, K. A., Pine, D. S., Pérez-Edgar, K., Henderson, H. A., Diaz, Y., … & Fox, N. A. (2009). Stable behavioral inhibition during infancy and early childhood predicts the development of anxiety disorders in adolescence. Journal of the American Academy of Child & Adolescent Psychiatry, 48, 928–935. doi:https://doi.org/10.1097/CHI.ob013e3181ae09df
Cioffi, C. C., Leve, L. D., Natsuaki, M. N., Shaw, D. S., Reiss, D., Ganiban, J. M., & Neiderhiser, J. M. (2021). Examining reciprocal associations between parent depressive symptoms and child internalizing symptoms on subsequent psychiatric disorders: An adoption study. Depression and Anxiety, 38, 1211–1224.
Clauss, J. A., & Blackford, J. U. (2012). Behavioral inhibition and risk for developing social anxiety disorder: A meta-analytic study. Journal of the American Academy of Child and Adolescent Psychiatry, 51, 1066–1075.
Cole, C., Zapp, D. J., Fettig, N. B., & Perez-Edgar, K. (2016). Impact of attention biases to threat and effortful control on individual variations in negative affect and social withdrawal in very young children. Journal of Experimental Child Psychology, 141, 210–221.
Cole, P. M., & Hollenstein, T. (2018). Emotion regulation: A matter of time. Routledge.
Cole, P. M., Lougheed, J. P., & Ram, N. (2018). The Development of Emotion Regulation in Early Childhood: A Matter of Multiple Time Scales. In Emotion regulation (pp. 70–87): Routledge.
Coplan, R. J., Arbeau, K. A., & Armer, M. (2008). Don’t fret, be supportive! Maternal characteristics linking child shyness to psychosocial and school adjustment in kindergarten. Journal of Abnormal Child Psychology, 36(3), 359–371.
Creaven, A. M., Skowron, E. A., Hughes, B. M., Howard, S., & Loken, E. (2014). Dyadic concordance in mother and preschooler resting cardiovascular function varies by risk status. Developmental Psychobiology, 56(1), 142–152.
Creswell, C., Cooper, P., & Murray, L. (2010). Intergenerational transmission of anxious information processing biases. In Information processing biases and anxiety: A developmental perspective (pp. 279–295).
Creswell, C., Shildrick, S., & Field, A. P. (2011). Interpretation of ambiguity in children: A prospective study of associations with anxiety and parental interpretations. Journal of Child and Family Studies, 20(2), 240–250.
Crick, N. R., & Dodge, K. A. (1994). A review and reformulation of social information-processing mechanisms in children’s social adjustment. Psychological Bulletin, 115, 74–101. https://doi.org/10.1037/0033-2909.115.1.74
Criss, M. M., Shaw, D. S., & Ingoldsby, E. M. (2003). Mother–son positive synchrony in middle childhood: Relation to antisocial behavior. Social Development, 12(3), 379–400.
Cui, X., Bryant, D. M., & Reiss, A. L. (2012). NIRS-based hyperscanning reveals increased interpersonal coherence in superior frontal cortex during cooperation. NeuroImage, 59(3), 2430–2437.
Danko, C. M., O’Brien, K. A., Rubin, K. H., & Chronis-Tuscano, A. (2018). the turtle program: PCIT for young children displaying behavioral inhibition. In L. N. Niec (Ed.), Handbook of parent-child interaction therapy: innovations and applications for research and practice (pp. 85–98). Springer International Publishing.
Davis, E. L., Brooker, R. J., & Kahle, S. (2020). Considering context in the developmental psychobiology of self-regulation. Developmental Psychobiology., 14, 125–589.
Davis, M., Bilms, J., & Suveg, C. (2017). In sync and in control: A meta-analysis of parent–child positive behavioral synchrony and youth self-regulation. Family Process, 56(4), 962–980.
Davis, M., West, K., Bilms, J., Morelen, D., & Suveg, C. (2018). A systematic review of parent–child synchrony: It is more than skin deep. Developmental Psychobiology, 60(6), 674–691.
De Rosnay, M., Cooper, P. J., Tsigaras, N., & Murray, L. (2006). Transmission of social anxiety from mother to infant: An experimental study using a social referencing paradigm. Behaviour Research and Therapy, 44(8), 1165–1175.
Degnan, K. A., & Fox, N. A. (2007). Behavioral inhibition and anxiety disorders: Multiple levels of a resilience process. Development and Psychopathology, 19, 729–746. https://doi.org/10.1017/S0954579407000363
Degnan, K. A., Hane, A. A., Henderson, H. A., Moas, O. L., Reeb-Sutherland, B. C., & Fox, N. A. (2011). Longitudinal stability of temperamental exuberance and social-emotional outcomes in early childhood. Developmental Psychology, 47, 765–780.
Degnan, K. A., Henderson, H. A., Fox, N. A., & Rubin, K. H. (2008). Predicting social wariness in middle childhood: The moderating roles of child care history, maternal personality, and maternal behavior. Social Development, 17, 471–487.
Del Giudice, M., Ellis, B. J., & Shirtcliff, E. A. (2011). The adaptive calibration model of stress responsivity. Neuroscience & Biobehavioral Reviews, 35(7), 1562–1592.
Denham, S. A. (2019). Emotional competence during childhood and adolescence. In Handbook of emotional development (pp. 493–541): Springer.
Denham, S. A., Bassett, H. H., & Wyatt, T. (2007). The socialization of emotional competence. In Handbook of Socialization: Theory and Research (pp. 614–637).
Denham, S. A., & Kochanoff, A. T. (2002). Parental contributions to preschoolers’ understanding of emotion. Marriage & Family Review, 34(3–4), 311–343.
DePasquale, C. E. (2020). A systematic review of caregiver–child physiological synchrony across systems: Associations with behavior and child functioning. Development and Psychopathology, 32(5), 1754–1777.
Dikker, S., Michalareas, G., Oostrik, M., Serafimaki, A., Kahraman, H. M., Struiksma, M. E., & Poeppel, D. (2021). Crowdsourcing neuroscience: inter-brain coupling during face-to-face interactions outside the laboratory. NeuroImage, 227, 117436.
Dikker, S., Wan, L., Davidesco, I., Kaggen, L., Oostrik, M., McClintock, J., … & Ding, M. (2017). Brain-to-brain synchrony tracks real-world dynamic group interactions in the classroom. Current Biology, 27(9), 1375–1380
Dollar, J. M., & Calkins, S. D. (2019). The development of anger. In Handbook of emotional development (pp. 199–225): Springer.
Dunne, G., & Askew, C. (2013). Vicarious learning and unlearning of fear in childhood via mother and stranger models. Emotion, 13(5), 974.
Eisenberg, N., Cumberland, A., Spinrad, T., Fabes, R. A., Shepard, S., Reiser, M., … & Guthrie, I. (2001). The relations of regulation and emotionality to children's externalizing and internalizing problem behavior. Child Development, 72, 1112–1134
Eisenberg, N., Cumberland, A., & Spinrad, T. L. (1998). Parental socialization of emotion. Psychological Inquiry, 9, 241–273.
Eley, T. C., McAdams, T. A., Rijsdijk, F. V., Lichtenstein, P., Narusyte, J., Reiss, D., … & Neiderhiser, J. M. (2015). The intergenerational transmission of anxiety: A children-of-twins study. American Journal of Psychiatry, 172(7), 630–637.
Ellis, B. J., & Boyce, W. T. (2008). Biological sensitivity to context. Current Directions in Psychological Science, 17(3), 183–187.
Ellis, B. J., Boyce, W. T., Belsky, J., Bakermans-Kranenburg, M. J., & Van IJzendoorn, M. H. (2011). Differential susceptibility to the environment: An evolutionary–neurodevelopmental theory. Development and Psychopathology, 23(1), 7–28.
Essau, C., Conradt, J., & Petermann, F. (1999). Frequency and comorbidity of social phobia and social fears in adolescents. Behaviour, Research, & Therapy, 37, 881–843.
Feldman, R. (2007). Parent–infant synchrony: Biological foundations and developmental outcomes. Current Directions in Psychological Science, 16(6), 340–345.
Feldman, R., & Greenbaum, C. W. (1997). Affect regulation and synchrony in mother—infant play as precursors to the development of symbolic competence. Infant Mental Health Journal, 18(1), 4–23.
Feldman, R., Greenbaum, C. W., & Yirmiya, N. (1999). Mother–infant affect synchrony as an antecedent of the emergence of self-control. Developmental Psychology, 35(1), 223.
Feldman, R., Greenbaum, C. W., Yirmiya, N., & Mayes, L. C. (1996). Relations between cyclicity and regulation in mother-infant interaction at 3 and 9 months and cognition at 2 years. Journal of Applied Developmental Psychology, 17(3), 347–365.
Feldman, R., Magori-Cohen, R., Galili, G., Singer, M., & Louzoun, Y. (2011). Mother and infant coordinate heart rhythms through episodes of interaction synchrony. Infant Behavior and Development, 34(4), 569–577.
Fergusson, D., Horwood, L., & Lynskey, M. (1993). Prevalence and comorbidity of DSM-III-R diagnoses in a birth cohort of 15 year olds. Journal of the American Academy of Child and Adolescent Psychiatry, 32, 1127–1134.
Ferrari, M., & Quaresima, V. (2012). A brief review on the history of human functional near-infrared spectroscopy (fNIRS) development and fields of application. NeuroImage, 63(2), 921–935.
Ferrer, E., & Steele, J. (2012). Dynamic systems analysis of affective processes in dyadic interactions using differential equations. In Advances in longitudinal methods in the social and behavioral sciences (pp. 111–134).
Field, T., Healy, B., & LeBlanc, W. G. (1989). Sharing and synchrony of behavior states and heart rate in nondepressed versus depressed mother-infant interactions. Infant Behavior and Development, 12(3), 357–376.
Fishburn, F. A., Murty, V. P., Hlutkowsky, C. O., MacGillivray, C. E., Bemis, L. M., Murphy, M. E., … & Perlman, S. B. (2018). Putting our heads together: interpersonal neural synchronization as a biological mechanism for shared intentionality. Social Cognitive and Affective Neuroscience, 13(8), 841–849
Fox, A. S., Oler, J. A., Birn, R. M., Shackman, A. J., Alexander, A. L., & Kalin, N. H. (2018). Functional connectivity within the primate extended amygdala is heritable and associated with early-life anxious temperament. The Journal of Neuroscience, 38(35), 7611–7621. https://doi.org/10.1523/jneurosci.0102-18.2018
Fox, A. S., Oler, J. A., Shackman, A. J., Shelton, S. E., Raveendran, M., McKay, D. R., … & Kalin, N. H. (2015). Intergenerational neural mediators of early-life anxious temperament. Proceedings of the National Academy of Sciences, 112(29), 9118–9122. https://doi.org/10.1073/pnas.1508593112
Fox, N. A., Hane, A. A., & Pérez-Edgar, K. (2006). Psychophysiological methods for the study of developmental psychopathology. In D. Cicchetti & D. J. Cohen (Eds.), Developmental Psychopathology (Vol. 2, pp. 381–426). Wiley.
Fox, N. A., Henderson, H. A., Marshall, P. J., Nichols, K. E., & Ghera, M. M. (2005). Behavioral inhibition: Linking biology and behavior within a developmental framework. Annual Review of Psychology, 56, 235–262.
Fox, N. A., Henderson, H. A., Pérez-Edgar, K., & White, L. (2008). The biology of temperament: An integrative approach. In C. Nelson & M. Luciana (Eds.), The handbook of developmental cognitive neuroscience (pp. 839–854). MIT Press.
Fox, N. A., Henderson, H. A., Rubin, K. H., Calkins, S. D., & Schmidt, L. A. (2001). Continuity and discontinuity of behavioral inhibition and exuberance: Psychophysiological and behavioral influences across the first four years of life. Child Development, 72, 1–21. https://doi.org/10.1111/1467-8624.00262
Fox, N. A., Snidman, N., Haas, S. A., Degnan, K. A., & Kagan, J. (2015). The relations between reactivity at 4 months and behavioral inhibition in the second year: Replication across three independent samples. Infancy, 20(1), 98–114.
Fu, X., & Pérez-Edgar, K. (2019). Threat-related attention bias in socioemotional development: A critical review and methodological considerations. Developmental Review, 51, 31–57.
Garner, P. W., Jones, D. C., & Miner, J. L. (1994). Social competence among low-income preschoolers: Emotion socialization practices and social cognitive correlates. Child Development, 65(2), 622–637.
Geangu, E., Benga, O., Stahl, D., & Striano, T. (2010). Contagious crying beyond the first days of life. Infant Behavior and Development, 33(3), 279–288.
Gerull, F. C., & Rapee, R. M. (2002). Mother knows best: Effects of maternal modelling on the acquisition of fear and avoidance behaviour in toddlers. Behaviour Research and Therapy, 40(3), 279–287.
Giuliano, R. J., Skowron, E. A., & Berkman, E. T. (2015). Growth models of dyadic synchrony and mother–child vagal tone in the context of parenting at-risk. Biological Psychology, 105, 29–36.
Grabow, A. P., Khurana, A., Natsuaki, M. N., Neiderhiser, J. M., Harold, G. T., Shaw, D. S., … & Leve, L. D. (2017). Using an adoption–biological family design to examine associations between maternal trauma, maternal depressive symptoms, and child internalizing and externalizing behaviors. Development and Psychopathology, 29(5), 1707–1720.
Granat, A., Gadassi, R., Gilboa-Schechtman, E., & Feldman, R. (2017). Maternal depression and anxiety, social synchrony, and infant regulation of negative and positive emotions. Emotion, 17(1), 11.
Gregory, A., & Eley, T. (2007). Genetic influences on anxiety in children: What we’ve learned and where we’re heading. Clinical Child and Family Psychology Review, 10, 199–212.
Grossmann, T., Johnson, M. H., Lloyd-Fox, S., Blasi, A., Deligianni, F., Elwell, C., & Csibra, G. (2008). Early cortical specialization for face-to-face communication in human infants. Proceedings of the Royal Society b: Biological Sciences, 275(1653), 2803–2811.
Gunther, K. E., Fu, X., MacNeill, L., Vallorani, A., Ermanni, B., & Pérez-Edgar, K. (in press). Profiles of naturalistic attentional trajectories moderate the relation between BI and internalizing symptoms: A mobile eye tracking study. Research in Child and Adolescent Psychopathology.
Guyer, A. E., Jarcho, J. M., Pérez-Edgar, K., Degnan, K. A., Pine, D. S., Fox, N. A., & Nelson, E. E. (2015). Temperament and parenting styles in early childhood differentially influence neural response to peer evaluation in adolescence. Journal of Abnormal Child Psychology, 43, 863–874.
Hails, K. A., Reuben, J. D., Shaw, D. S., Dishion, T. J., & Wilson, M. N. (2018). Transactional associations among maternal depression, parent–child coercion, and child conduct problems during early childhood. Journal of Clinical Child & Adolescent Psychology, 47(sup1), S291–S305.
Hajal, N. J., & Paley, B. (2020). Parental emotion and emotion regulation: A critical target of study for research and intervention to promote child emotion socialization. Developmental Psychology, 56(3), 403–417.
Hajcak, G., Klawohn, J., & Meyer, A. (2019). The utility of event-related potentials in clinical psychology. Annual Review of Clinical Psychology, 15, 71–95.
Halberstadt, A. G., & Eaton, K. L. (2002). A meta-analysis of family expressiveness and children’s emotion expressiveness and understanding. Marriage & Family Review, 34(1–2), 35–62.
Ham, J., & Tronick, E. (2009). Relational psychophysiology: Lessons from mother–infant physiology research on dyadically expanded states of consciousness. Psychotherapy Research, 19(6), 619–632.
Hane, A. A., Cheah, C. S. L., Rubin, K. H., & Fox, N. A. (2008). The role of maternal behavior in the relation between shyness and social withdrawal in early childhood and social withdrawal in middle childhood. Social Development, 17, 795–811.
Harrist, A., & Waugh, R. (2002). Dyadic synchrony: It’s function and structure in child development. Developmental Review, 22(4), 555–592.
Hasson, U., Ghazanfar, A. A., Galantucci, B., Garrod, S., & Keysers, C. (2012). Brain-to-brain coupling: A mechanism for creating and sharing a social world. Trends in Cognitive Sciences, 16(2), 114–121.
Hastings, P. D., Rubin, K. H., Smith, K. A., & Wagner, N. J. (2019). Parenting behaviorally inhibited and socially withdrawn children. In M. H. Bornstein (Ed.), Handbook of parenting (pp. 467–495). New York: Routledge. https://www.taylorfrancis.com/books/edit/10.4324/9780429440847/handbook-parenting-marc-bornstein?refId=97bae4f8-05fc-4b90-bdab-29867619ad5d&context=ubx
Hayward, C., Killen, J., Kraemer, H., & Taylor, C. (1998). Linking self-reported childhood behavioral inhibition to adolescent social phobia. Journal of the American Academy of Child and Adolescent Psychiatry, 37, 1308–1316.
Helm, J. L., Miller, J. G., Kahle, S., Troxel, N. R., & Hastings, P. D. (2018). On measuring and modeling physiological synchrony in dyads. Multivariate Behavioral Research, 53(4), 521–543.
Herschell, A. D., Calzada, E. J., Eyberg, S. M., & McNeil, C. B. (2002). Parent-child interaction therapy: New directions in research. Cognitive and Behavioral Practice, 9(1), 9–16.
Hettema, J. M., Neale, M. C., & Kendler, K. S. (2001). A review and meta-analysis of the genetic epidemiology of anxiety disorders. American Journal of Psychiatry, 158, 1568–1578.
Hipwell, A. E., Tung, I., Northrup, J., & Keenan, K. (2019). Transgenerational associations between maternal childhood stress exposure and profiles of infant emotional reactivity. Development and Psychopathology, 31(3), 887–898.
Hollenstein, T. (2013). State space grids: Depicting dynamics across development. Springer.
Holodynski, M., & Friedlmeier, W. (2006). Development of emotions and emotion regulation. Springer Science & Business Media.
Hoyniak, C. P., Quiñones-Camacho, L. E., Camacho, M. C., Chin, J. H., Williams, E. M., Wakschlag, L. S., & Perlman, S. B. (2021). Adversity is Linked with Decreased Parent-Child Behavioral and Neural Synchrony. Developmental Cognitive Neuroscience, 24, 100937.
Hudson, J. L., & Rapee, R. M. (2001). Parent–child interactions and anxiety disorders: An observational study. Behaviour Research and Therapy, 39(12), 1411–1427.
Im-Bolter, N., Anam, M., & Cohen, N. J. (2015). Mother–child synchrony and child problem behavior. Journal of Child and Family Studies, 24(7), 1876–1885.
Johnco, C. J., Magson, N. R., Fardouly, J., Oar, E. L., Forbes, M. K., Richardson, C., & Rapee, R. M. (2021). The role of parenting behaviors in the bidirectional and intergenerational transmission of depression and anxiety between parents and early adolescent youth. Depression and Anxiety, 38, 1256–1266.
Johnson, J. G., Cohen, P., Kasen, S., Smailes, E., & Brook, J. S. (2001). Association of maladaptive parental behavior with psychiatric disorder among parents and their offspring. Archives of General Psychiatry, 58(5), 453–460.
Jongerling, J., Laurenceau, J.-P., & Hamaker, E. L. (2015). A multilevel AR (1) model: Allowing for inter-individual differences in trait-scores, inertia, and innovation variance. Multivariate Behavioral Research, 50(3), 334–349.
Kellerman, A. M., Schwichtenberg, A., Abu-Zhaya, R., Miller, M., Young, G. S., & Ozonoff, S. (2020). Dyadic synchrony and responsiveness in the first year: Associations with autism risk. Autism Research, 13(12), 2190–2201.
Kendall, P. C., Panichelli-Mindel, S. M., Sugarman, A., & Callahan, S. A. (1997). Exposure to child anxiety: Theory, research, and practice. Clinical Psychology: Science and Practice, 4(1), 29.
Kenny, D. A., Kashy, D. A., & Cook, W. L. (2006). Dyadic data analysis. Guilford Press.
Kerr, K. L., Ratliff, E. L., Cosgrove, K. T., Bodurka, J., Morris, A. S., & Simmons, W. K. (2019). Parental influences on neural mechanisms underlying emotion regulation. Trends in Neuroscience and Education, 16, 100118.
Kiel, E. J., & Buss, K. A. (2011). Prospective relations among fearful temperament, protective parenting, and social withdrawal: The role of maternal accuracy in a moderated mediation framework. Journal of Abnormal Child Psychology, 39, 953–966.
Kiel, E. J., & Buss, K. A. (2012). Associations among context-specific maternal protective behavior, toddlers’ fearful temperament, and maternal accuracy and goals. Social Development, 21, 742–760.
Kiel, E. J., & Buss, K. A. (2014). Dysregulated fear in toddlerhood predicts kindergarten social withdrawal through protective parenting. Infant and Child Development, 23, 304–313.
Kiel, E. J., & Hummel, A. C. (2017). Contextual influences on concordance between maternal report and laboratory observation of toddler fear. Emotion, 17, 240–250.
Kiel, E. J., & Kalomiris, A. E. (2015). Current themes in understanding children’s emotion regulation as developing from within the parent–child relationship. Current Opinion in Psychology, 3, 11–16.
Kiel, E. J., Premo, J. E., & Buss, K. A. (2016). Maternal encouragement to approach novelty: A curvilinear relation to change in anxiety for inhibited toddlers. Journal of Abnormal Child Psychology, 44, 433–444.
Kiel, E. J., Price, N. N., & Buss, K. A. (2021). Maternal anxiety and toddler inhibited temperament predict maternal socialization of worry. Social Development, 30(1), 258–273.
Kiff, C. J., Lengua, L. J., & Zalewski, M. (2011). Nature and nurturing: Parenting in the context of child temperament. Clinical Child and Family Psychology Review, 14(3), 251.
Kinreich, S., Djalovski, A., Kraus, L., Louzoun, Y., & Feldman, R. (2017). Brain-to-brain synchrony during naturalistic social interactions. Scientific Reports, 7(1), 17060. https://doi.org/10.1038/s41598-017-17339-5
Klinnert, M. D., Campos, J. J., Sorce, J. F., Emde, R. N., & Svejda, M. (1983). Emotions as behavior regulators: Social referencing in infancy. In Emotions in early development (pp. 57–86): Elsevier.
Knyazev, G. G. (2011). Cross-frequency coupling of brain oscillations: An impact of state anxiety. International Journal of Psychophysiology, 80, 236–245.
Knyazev, G. G., Schutter, D. J., & van Honk, J. (2006). Anxious apprehension increases coupling of delta and beta oscillations. International Journal of Psychophysiology, 61(2), 283–287.
Kochanska, G., Aksan, N., Prisco, T. R., & Adams, E. E. (2008). Mother–child and father–child mutually responsive orientation in the first 2 years and children’s outcomes at preschool age: Mechanisms of influence. Child Development, 79(1), 30–44.
Kruppa, J. A., Reindl, V., Gerloff, C., Oberwelland Weiss, E., Prinz, J., Herpertz-Dahlmann, B., … & Schulte-Rüther, M. (2020). Brain and motor synchrony in children and adolescents with ASD—a fNIRS hyperscanning study. Social Cognitive and Affective Neuroscience, 16(1-2), 103–116. doi:https://doi.org/10.1093/scan/nsaa092
Laurent, H. K. (2014). Clarifying the contours of emotion regulation: Insights from parent-child stress research. Child Development Perspectives, 8(1), 30–35. https://doi.org/10.1111/cdep.12058
Laurent, H. K., Ablow, J. C., & Measelle, J. (2011). Risky shifts: How the timing and course of mothers’ depressive symptoms across the perinatal period shape their own and infant’s stress response profiles. Development and Psychopathology, 23(2), 521–538.
Laurent, H. K., Ablow, J. C., & Measelle, J. (2012). Taking stress response out of the box: Stability, discontinuity, and temperament effects on HPA and SNS across social stressors in mother–infant dyads. Developmental Psychology, 48(1), 35.
Laurent, H. K., Sbrilli, M., Dawson, D., Finnegan, M., & Ramdas-Neal, D. (2021). Disentangling levels of mother–infant neuroendocrine attunement and longitudinal relations with maternal risk and protective factors. Developmental Psychobiology, 63(1), 88–97.
Leerkes, E. M., Bailes, L. G., & Augustine, M. E. (2020). The intergenerational transmission of emotion socialization. Developmental Psychology, 56(3), 390.
Lenze, S. N., Pautsch, J., & Luby, J. (2011). Parent–child interaction therapy emotion development: A novel treatment for depression in preschool children. Depression and Anxiety, 28(2), 153–159.
Lester, K. J., Field, A. P., Oliver, S., & Cartwright-Hatton, S. (2009). Do anxious parents interpretive biases towards threat extend into their child’s environment? Behaviour Research and Therapy, 47(2), 170–174.
Lin, B., Kaliush, P. R., Conradt, E., Terrell, S., Neff, D., Allen, A. K., … Crowell, S. E. (2019). Intergenerational transmission of emotion dysregulation: Part I. Psychopathology, self-injury, and parasympathetic responsivity among pregnant women. Development and Psychopathology, 31(3), 817–831.
Lindenberger, U., Li, S.-C., Gruber, W., & Müller, V. (2009). Brains swinging in concert: Cortical phase synchronization while playing guitar. BMC Neuroscience, 10(1), 1–12.
Liu, T., Duan, L., Dai, R., Pelowski, M., & Zhu, C. (2021). Team-work, Team-brain: Exploring synchrony and team interdependence in a nine-person drumming task via multiparticipant hyperscanning and inter-brain network topology with fNIRS. NeuroImage, 237, 118147.
LoBue, V., & Adolph, K. E. (2019). Fear in infancy: Lessons from snakes, spiders, heights, and strangers. Developmental Psychology, 55(9), 1889–1907.
LoBue, V., Kim, E., & Delgado, M. R. (2019). Fear in development. In V. LoBue, K. Pérez-Edgar, & K. A. Buss (Eds.), Handbook of emotional development. Springer.
LoBue, V., Matthews, K., Harvey, T., & Stark, S. L. (2014). What accounts for the rapid detection of threat? Evidence for an advantage in perceptual and behavioral responding from eye movements. Emotion, 14, 816–823.
Luck, S., Woodman, G., & Vogel, E. (2000). Event-related potential studies of attention. Trends in Cognitive Sciences, 4, 432–440.
Lunkenheimer, E., Brown, K. M., & Fuchs, A. (2021). Differences in mother–child and father–child RSA synchrony: Moderation by child self-regulation and dyadic affect. Developmental Psychobiology, 15, 1478.
Lunkenheimer, E., Busuito, A., Brown, K. M., & Skowron, E. A. (2018). Mother–child coregulation of parasympathetic processes differs by child maltreatment severity and subtype. Child Maltreatment, 23(3), 211–220.
Lunkenheimer, E., Hamby, C. M., Lobo, F. M., Cole, P. M., & Olson, S. L. (2020). The role of dynamic, dyadic parent–child processes in parental socialization of emotion. Developmental Psychology, 56(3), 566.
Lunkenheimer, E., Olson, S. L., Hollenstein, T., Sameroff, A. J., & Winter, C. (2011). Dyadic flexibility and positive affect in parent–child coregulation and the development of child behavior problems. Development and Psychopathology, 23(2), 577–591.
Lunkenheimer, E., Ram, N., Skowron, E., & Yin, P. (2017). Harsh parenting, child behavior problems, and the dynamic coupling of parents’ and children’s positive behaviors. Journal of Family Psychology, 31, 689–698.
Lunkenheimer, E., Tiberio, S. S., Buss, K. A., Lucas-Thompson, R. G., Boker, S. M., & Timpe, Z. C. (2015). Coregulation of respiratory sinus arrhythmia between parents and preschoolers: Differences by children’s externalizing problems. Developmental Psychobiology, 57(8), 994–1003.
Lunkenheimer, E., Tiberio, S. S., Skoranski, A. M., Buss, K. A., & Cole, P. M. (2018). Parent-child coregulation of parasympathetic processes varies by social context and risk for psychopathology. Psychophysiology, 55(2), e12985.
Maag, B., Phelps, R. A., & Kiel, E. J. (2021). Do maternal parenting behaviors indirectly link toddler dysregulated fear and child anxiety symptoms? Child Psychiatry & Human Development, 52(2), 225–235.
MacNeill, L., Fu, X., Buss, K. A., & Pérez-Edgar, K. (in press). Do you see what I mean?: Using mobile eye-tracking to capture parent-child dynamics in the context of anxiety risk. Development and Psychopathology.
McGee, R., Feehan, M., Williams, S., Partridge, F., Silva, P., & Kelly, J. (1990). DSM-III disorders in a large sample of adolescents. Journal of the American Academy of Child and Adolescent Psychiatry, 29, 611–619.
McLeod, B. D., Wood, J. J., & Weisz, J. R. (2007). Examining the association between parenting and childhood anxiety: A meta-analysis. Clinical Psychology Review, 27(2), 155–172.
McNeish, D., & Hamaker, E. L. (2019). A primer on two-level dynamic structural equation models for intensive longitudinal data in Mplus. Psychological Methods., 25, 610.
Messinger, D., Mitsven, S. G., Ahn, Y. A., Prince, E. B., Sun, L., & Rivero-Fernández, C. (2019). Happiness and Joy. In Handbook of Emotional Development (pp. 171–198): Springer.
Moore, G. A., & Calkins, S. D. (2004). Infants’ vagal regulation in the still-face paradigm is related to dyadic coordination of mother-infant interaction. Developmental Psychology, 40(6), 1068.
Moore, G. A., Hill-Soderlund, A. L., Propper, C. B., Calkins, S. D., Mills-Koonce, W. R., & Cox, M. J. (2009). Mother–infant vagal regulation in the face-to-face still-face paradigm is moderated by maternal sensitivity. Child Development, 80(1), 209–223.
Moore, G. A., Powers, C. J., Bass, A. J., Cohn, J. F., Propper, C. B., Allen, N. B., & Lewinsohn, P. M. (2013). Dyadic interaction: Greater than the sum of its parts? Infancy, 18, 490–515.
Morales, S. (2017). Anxiety and Attention Bias Towards Threat: A Developmental and Multi-Method Approach. (PhD). The Pennsylvania State University
Morales, S., Beekman, C., Blandon, A. Y., Stifter, C. A., & Buss, K. A. (2015). Longitudinal associations between temperament and socioemotional outcomes in young children: The moderating role of RSA and gender. Developmental Psychobiology, 57, 105–119.
Morales, S., Brown, K. M., Taber-Thomas, B. C., LoBue, V., Buss, K. A., & Pérez-Edgar, K. (2017). Maternal anxiety predicts attentional bias towards threat in infancy. Emotion, 17, 874–883.
Morales, S., Fu, X., & Pérez-Edgar, K. (2016). A developmental neuroscience perspective on affect biased attention. Developmental Cognitive Neuroscience, 21, 26–41.
Morales, S., Pérez-Edgar, K., & Buss, K. A. (2015). Attention biases towards and away from threat mark the relation between early dysregulated fear and the later emergence of social withdrawal. Journal of Abnormal Child Psychology, 43, 1067–1078.
Müller, V., Sänger, J., & Lindenberger, U. (2013). Intra-and inter-brain synchronization during musical improvisation on the guitar. PLoS ONE, 8(9), 73852.
Muris, P., Steerneman, P., Merckelbach, H., & Meesters, C. (1996). The role of parental fearfulness and modeling in children’s fear. Behaviour Research and Therapy, 34(3), 265–268.
Murray, L., De Rosnay, M., Pearson, J., Bergeron, C., Schofield, E., Royal-Lawson, M., & Cooper, P. J. (2008). Intergenerational transmission of social anxiety: The role of social referencing processes in infancy. Child Development, 79(4), 1049–1064.
Murray, L., Pella, J. E., De Pascalis, L., Arteche, A., Pass, L., Percy, R., … & Cooper, P. J. (2014). Socially anxious mothers' narratives to their children and their relation to child representations and adjustment. Development and Psychopathology, 26(4pt2), 1531–1546.
Nguyen, T., Schleihauf, H., Kayhan, E., Matthes, D., Vrtička, P., & Hoehl, S. (2020). The effects of interaction quality on neural synchrony during mother-child problem solving. Cortex, 124, 235–249.
Ollendick, T. H., & Hirshfeld-Becker, D. R. (2002). The developmental psychopathology of social anxiety disorder. Biological Psychiatry, 51, 44–58.
Ostlund, B. D., Measelle, J. R., Laurent, H. K., Conradt, E., & Ablow, J. C. (2017). Shaping emotion regulation: Attunement, symptomatology, and stress recovery within mother–infant dyads. Developmental Psychobiology, 59(1), 15–25.
Ostlund, B. D., Vlisides-Henry, R. D., Crowell, S. E., Raby, K. L., Terrell, S., Brown, M. A., … & Shakib, J. H. (2019). Intergenerational transmission of emotion dysregulation: Part II. Developmental origins of newborn neurobehavior. Development and Psychopathology, 31(3), 833–846.
Pemberton, C. K., Neiderhiser, J. M., Leve, L. D., Natsuaki, M. N., Shaw, D. S., Reiss, D., & Ge, X. (2010). Influence of parental depressive symptoms on adopted toddler behaviors: An emerging developmental cascade of genetic and environmental effects. Development and Psychopathology, 22(4), 803–818.
Percy, R., Creswell, C., Garner, M., O’Brien, D., & Murray, L. (2016). Parents’ verbal communication and childhood anxiety: A systematic review. Clinical Child and Family Psychology Review, 19(1), 55–75.
Pérez-Edgar, K. (2019). Through the Looking Glass: Temperament and Emotion as Separate and Interwoven Constructs. In Handbook of Emotional Development (pp. 139–168): Springer.
Pérez-Edgar, K., Bar-Haim, Y., McDermott, J. M., Chronis-Tuscano, A., Pine, D. S., & Fox, N. A. (2010). Attention biases to threat and behavioral inhibition in early childhood shape adolescent social withdrawal. Emotion, 10, 349–357. https://doi.org/10.1037/a0018486
Pérez-Edgar, K., Kujawa, A., Nelson, S. K., Cole, C., & Zapp, D. J. (2013). The relation between electroencephalogram asymmetry and attention biases to threat at baseline and under stress. Brain and Cognition, 82(3), 337–343.
Pérez-Edgar, K., MacNeill, L., & Fu, X. (2020). Navigating through the experienced environment: Insights from mobile eye-tracking. Current Directions in Psychological Science, 29, 286–292.
Pérez-Edgar, K., McDermott, J. M., Korelitz, K., Degnan, K. A., Curby, T. W., Pine, D. S., & Fox, N. A. (2010). Patterns of sustained attention in infancy shape the developmental trajectory of social behavior from toddlerhood through adolescence. Developmental Psychology, 46, 1723–1730. https://doi.org/10.1037/a0021064
Pérez-Edgar, K., Reeb-Sutherland, B. C., McDermott, J. M., White, L. K., Henderson, H. A., Degnan, K. A., … & Fox, N. A. (2011). Attention biases to threat link behavioral inhibition to social withdrawal over time in very young children. Journal of Abnormal Child Psychology, 39, 885–895. https://doi.org/10.1007/s10802-011-9495-5
Pérez-Edgar, K., Taber-Thomas, B., Auday, E., & Morales, S. (2014). Temperament and attention as core mechanisms in the early emergence of anxiety. In K. Lagattuta (Ed.), Children and emotion: New Iisights into developmental affective science (Vol. 26, pp. 42–56): Karger Publishing
Phelps, R. A., Brooker, R. J., & Buss, K. A. (2016). Toddlers’ dysregulated fear predicts delta–beta coupling during preschool. Developmental Cognitive Neuroscience, 17, 28–34.
Phillips, D. A., Fox, N. A., & Gunnar, M. R. (2011). Same place, different experiences: Bringing individual differences to research in child care. Child Development Perspectives, 5, 44–49.
Piazza, E. A., Cohen, A., Trach, J., & Lew-Williams, C. (2021). Neural synchrony predicts children’s learning of novel words. Cognition, 214, 104752.
Pratt, M., Apter-Levi, Y., Vakart, A., Kanat-Maymon, Y., Zagoory-Sharon, O., & Feldman, R. (2017). Mother-child adrenocortical synchrony; Moderation by dyadic relational behavior. Hormones and Behavior, 89, 167–175.
Prior, M., Smart, D., Sanson, A., & Oberklaid, F. (2000). Does shy-inhibited temperament in childhood lead to anxiety problems in adolescence? Journal of the American Academy of Child & Adolescent Psychiatry, 39(4), 461–468.
Puliafico, A. C., Comer, J. S., & Pincus, D. B. (2012). Adapting parent-child interaction therapy to treat anxiety disorders in young children. Child and Adolescent Psychiatric Clinics, 21(3), 607–619.
Quiñones-Camacho, L. E., Fishburn, F. A., Belardi, K., Williams, D. L., Huppert, T. J., & Perlman, S. B. (2021). Dysfunction in interpersonal neural synchronization as a mechanism for social impairment in autism spectrum disorder. Autism Research. https://doi.org/10.1002/aur.2513
Quiñones-Camacho, L. E., Hoyniak, C. P., Wakschlag, L. S., & Perlman, S. B. (2021). Getting in synch: Unpacking the role of parent–child synchrony in the development of internalizing and externalizing behaviors. Development and Psychopathology, 24, 1–13.
Quiñones-Camacho, L. E., Fishburn, F. A., Camacho, M. C., Hlutkowsky, C. O., Huppert, T. J., Wakschlag, L. S., & Perlman, S. B. (2019). Parent–child neural synchrony: a novel approach to elucidating dyadic correlates of preschool irritability. Journal of Child Psychology and Psychiatry., 61, 1213–1233.
Radtke, S. R., Strege, M. V., & Ollendick, T. H. (2020). Exposure therapy for children and adolescents with social anxiety disorder. In Exposure Therapy for children with anxiety and OCD (pp. 193–219): Elsevier.
Rapee, R. M., Kennedy, S. J., Ingram, M., Edwards, S., & Sweeney, L. (2010). Altering the trajectory of anxiety in at-risk young children. American Journal of Psychiatry, 167, 1518–1525.
Reindl, V., Gerloff, C., Scharke, W., & Konrad, K. (2018). Brain-to-brain synchrony in parent-child dyads and the relationship with emotion regulation revealed by fNIRS-based hyperscanning. NeuroImage, 178, 493–502.
Reynolds, G., Askew, C., & Field, A. P. (2018). Behavioral Inhibition and the Associative Learning of Fear. In Behavioral inhibition (pp. 263–282): Springer.
Rosenbaum, J. F., Biederman, J., Bolduc, E., Hirshfeld, D., Faraone, S., & Kagan, J. (1992). Comorbidity of parental anxiety disorders as risk for childhood-onset anxiety in inhibited children. American Journal of Psychiatry, 149, 475–481.
Rosenbaum, J. F., Biederman, J., Gersten, M., Hirshfeld, D. R., Meminger, S. R., Herman, J. B., … & Snidman, N. (1988). Behavioral inhibition in children of parents with panic disorder and agoraphobia: A controlled study. Archives of General Psychiatry, 45, 463–470.
Rubin, K. H., Barstead, M. G., Smith, K. A., & Bowker, J. C. (2018). Peer relations and the behaviorally inhibited child. In K. Pérez-Edgar & N. A. Fox (Eds.), Behavioral inhibition (pp. 157–184). Springer.
Rubin, K. H., Burgess, K. B., Dwyer, K. M., & Hastings, P. D. (2003). Predicting preschoolers’ externalizing behaviors from toddler temperament, conflict, and maternal negativity. Developmental Psychology, 39(1), 164.
Rubin, K. H., Nelson, L. J., Hastings, P., & Asendorpf, J. (1999). The transaction between parents’ perceptions of their children’s shyness and their parenting styles. International Journal of Behavioral Development, 23(4), 937–957.
Rudasill, K. M., & Rimm-Kaufman, S. E. (2009). Teacher–child relationship quality: The roles of child temperament and teacher–child interactions. Early Childhood Research Quarterly, 24(2), 107–120.
Ruffman, T., Lorimer, B., & Scarf, D. (2017). Do infants really experience emotional contagion? Child Development Perspectives, 11(4), 270–274.
Russell, C. L., Bard, K. A., & Adamson, L. B. (1997). Social referencing by young chimpanzees (Pan troglodytes). Journal of Comparative Psychology, 111(2), 185.
Sameroff, A. (1975). Transactional models in early social relations. Human Development, 18(1–2), 65–79.
Sanislow, C. A., Pine, D. S., Quinn, K. J., Kozak, M. J., Garvey, M. A., Heinssen, R. K., … & Cuthbert, B. N. (2010). Developing constructs for psychopathology research: Research domain criteria. Journal of Abnormal Psychology, 119, 631–639.
Schwartz, C. E., Snidman, N., & Kagan, J. (1999). Adolescent social anxiety as an outcome of inhibited temperament in childhood. Journal of the American Academy of Child and Adolescent Psychiatry, 38, 1008–1015.
Sequeira, S. L., Rosen, D. K., Silk, J. S., Hutchinson, E., Allen, K. B., Jones, N. P., … & Ladouceur, C. D. (2021). “Don’t judge me!”: Links between in vivo attention bias toward a potentially critical judge and fronto-amygdala functional connectivity during rejection in adolescent girls. Developmental Cognitive Neuroscience, 49, 100960.
Shih, E. W., Quiñones-Camacho, L. E., Karan, A., & Davis, E. L. (2019). Physiological contagion in parent–child dyads during an emotional challenge. Social Development, 28(3), 620–636.
Shutts, K., Kinzler, K. D., & DeJesus, J. M. (2013). Understanding infants’ and children’s social learning about foods: Previous research and new prospects. Developmental Psychology, 49(3), 419.
Silverman, W. K., Marin, C. E., Rey, Y., Jaccard, J., & Pettit, J. W. (2021). Directional effects of parent and child anxiety 1 year following treatment of child anxiety, and the mediational role of parent psychological control. Depression and Anxiety, 38, 1289–1297.
Silvers, J. A., Callaghan, B. L., VanTieghem, M., Choy, T., O’Sullivan, K., Tottenham, N., & Silvers, J. A. (2020). An Exploration of Amygdala-Prefrontal Mechanisms in the Intergenerational Transmission of Learned Fear Abbreviated title: Intergenerational fear. Developmental Science, 14, e13056.
Smeekens, S., Riksen-Walraven, J. M., & van Bakel, H. J. (2007). Multiple determinants of externalizing behavior in 5-year-olds: A longitudinal model. Journal of Abnormal Child Psychology, 35(3), 347–361.
Smith, C., Jones, E. J., Charman, T., Clackson, K., Mirza, F., & Wass, S. (2019). Anxious parents show higher physiological synchrony with their infants. Psychological Medicine, 13, 1–11.
Sorce, J. F., Emde, R. N., Campos, J. J., & Klinnert, M. D. (1985). Maternal emotional signaling: Its effect on the visual cliff behavior of 1-year-olds. Developmental Psychology, 21(1), 195.
Spence, S. H., & Rapee, R. M. (2016). The etiology of social anxiety disorder: An evidence-based model. Behaviour Research and Therapy, 86, 50–67.
Su, W.-C., Culotta, M., Mueller, J., Tsuzuki, D., Pelphrey, K., & Bhat, A. (2020). Differences in cortical activation patterns during action observation, action execution, and interpersonal synchrony between children with or without autism spectrum disorder (ASD): An fNIRS pilot study. PLoS ONE, 15(10), e0240301.
Suveg, C., Shaffer, A., & Davis, M. (2016). Family stress moderates relations between physiological and behavioral synchrony and child self-regulation in mother–preschooler dyads. Developmental Psychobiology, 58(1), 83–97.
Todd, R. M., Cunningham, W. A., Anderson, A. K., & Thompson, E. (2012). Affect-biased attention as emotion regulation. Trends in Cognitive Sciences, 16, 365–372. https://doi.org/10.1016/j.tics.2012.06.003
Torpey, D. C., Hajcak, G., Kim, J., Kujawa, A. J., Dyson, M. W., Olino, T. M., & Klein, D. N. (2013). Error-related brain activity in young children: Associations with parental anxiety and child temperamental negative emotionality. Journal of Child Psychology and Psychiatry, 54(8), 854–862.
Tramacere, A., & Ferrari, P. F. (2016). Faces in the mirror, from the neuroscience of mimicry to the emergence of mentalizing. Journal of Anthropological Sciences, 94, 1–14.
Tronick, E. Z. (1989). Emotions and emotional communication in infants. American Psychologist, 44, 112.
Tronick, E. Z., & Cohn, J. F. (1989). Infant-mother face-to-face interaction: Age and gender differences in coordination and the occurrence of miscoordination. Child Development, 14, 85–92.
Turpyn, C. C., Poon, J. A., Ross, C. E., Thompson, J. C., & Chaplin, T. M. (2018). Associations between parent emotional arousal and regulation and adolescents’ affective brain response. Social Development, 27(1), 3–18.
Urakawa, S., Takamoto, K., Ishikawa, A., Ono, T., & Nishijo, H. (2015). Selective medial prefrontal cortex responses during live mutual gaze interactions in human infants: An fNIRS study. Brain Topography, 28(5), 691–701.
Valiente, C., Eisenberg, N., Fabes, R. A., Shepard, S. A., Cumberland, A., & Losoya, S. H. (2004). Prediction of children’s empathy-related responding from their effortful control and parents’ expressivity. Developmental Psychology, 40(6), 911.
Van Bockstaele, B., Verschuere, B., Tibboel, H., De Houwer, J., Crombez, G., & Koster, E. H. W. (2014). A review of current evidence for the causal impact of attentional bias on fear and anxiety. Psychological Bulletin, 140(3), 682–721.
Van Leeuwen, P., Geue, D., Thiel, M., Cysarz, D., Lange, S., Romano, M. C., … & Grönemeyer, D. (2009). Influence of paced maternal breathing on fetal–maternal heart rate coordination. Proceedings of the National Academy of Sciences, 106(33), 13661–13666.
van Peer, J. M., Roelofs, K., & Spinhoven, P. (2008). Cortisol administration enhances the coupling of midfrontal delta and beta oscillations. International Journal of Psychophysiology, 67(2), 144–150.
Wass, S. V., Clackson, K., & Leong, V. (2018). Increases in arousal are more long-lasting than decreases in arousal: On homeostatic failures during emotion regulation in infancy. Infancy, 23, 628–649.
Wass, S. V., Whitehorn, M., Phillips, E., Haresign, I. M., & Leong, V. (2020). Interpersonal neural synchrony and responsivity during early learning interactions. Trends in Cognitive Science, 24, 329–342.
Waters, S. F., West, T. V., Karnilowicz, H. R., & Mendes, W. B. (2017). Affect contagion between mothers and infants: Examining valence and touch. Journal of Experimental Psychology: General, 146(7), 1043–1051.
Waters, S. F., West, T. V., & Mendes, W. B. (2014). Stress contagion: Physiological covariation between mothers and infants. Psychological Science, 25(4), 934–942.
Wesseldijk, L. W., Dieleman, G. C., van Steensel, F. J., Bleijenberg, E. J., Bartels, M., Bögels, S. M., & Middeldorp, C. M. (2018). Do parental psychiatric symptoms predict outcome in children with psychiatric disorders? A naturalistic clinical study. Journal of the American Academy of Child & Adolescent Psychiatry, 57(9), 669–677.
White, L. K., Degnan, K. A., Henderson, H. A., Pérez-Edgar, K., Walker, O. L., Schechner, T., … Fox, N. A. (2017). Developmental relations between behavioral inhibition, anxiety, and attention biases to threat and positive information. Child Development, 88, 141–155
Williamson, D. E., Coleman, K., Bacanu, S.-A., Devlin, B. J., Rogers, J., Ryan, N. D., & Cameron, J. L. (2003). Heritability of fearful-anxious endophenotypes in infant rhesus macaques: A preliminary report. Biological Psychiatry, 53(4), 284–291.
Woody, M. L., Feurer, C., Sosoo, E. E., Hastings, P. D., & Gibb, B. E. (2016). Synchrony of physiological activity during mother–child interaction: Moderation by maternal history of major depressive disorder. Journal of Child Psychology and Psychiatry, 57(7), 843–850.
Woody, M. L., Rosen, D., Allen, K. B., Price, R. B., Hutchinson, E., Amole, M. C., & Silk, J. S. (2019). Looking for the negative: Depressive symptoms in adolescent girls are associated with sustained attention to a potentially critical judge during in vivo social evaluation. Journal of Experimental Child Psychology, 179, 90–102.
Yirmiya, K., Motsan, S., Kanat-Maymon, Y., & Feldman, R. (2021). From mothers to children and back: Bidirectional processes in the cross-generational transmission of anxiety from early childhood to early adolescence. Depression and Anxiety, 38, 1298–1312.
Young, K. S., Parsons, C. E., Stein, A., Vuust, P., Craske, M. G., & Kringelbach, M. L. (2017). The neural basis of responsive caregiving behaviour: Investigating temporal dynamics within the parental brain. Behavioural Brain Research, 325, 105–116.
Zhou, Q., Eisenberg, N., Losoya, S. H., Fabes, R. A., Reiser, M., Guthrie, I. K., … Shepard, S. A. (2002). The relations of parental warmth and positive expressiveness to children's empathy‐related responding and social functioning: A longitudinal study. Child Development, 73(3), 893–915.
Acknowledgements
This paper was supported by NIH Grants R56 MH126349 and R01 MH130007 to MPIs Pérez-Edgar and Perlman.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no disclosures.
Ethical approval
This manuscript is a review/theoretical paper and does not fall under IRB approval. No subjects were tested as part of this manuscript.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised: The author name 'Erika Lunkenheimer' was incorrectly written as 'Erica Lunkenheimer'.
Rights and permissions
About this article

Cite this article
Perlman, S.B., Lunkenheimer, E., Panlilio, C. et al. Parent-to-Child Anxiety Transmission Through Dyadic Social Dynamics: A Dynamic Developmental Model. Clin Child Fam Psychol Rev 25, 110–129 (2022). https://doi.org/10.1007/s10567-022-00391-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10567-022-00391-7