
Abstract
Cryopreservation of teeth before autotransplantation may create new possibilities in dentistry. The purpose of this study was to examine the effect of a standardised cryopreservation procedure on human periodontal ligament (PDL) cell cultures. Human PDL fibroblasts obtained from immature third molars of 11 patients were cultured and divided into two groups. The experimental group was cryopreserved and cultured after thawing. The control group was cultured without cryopreservation. A comparison was made between cryopreserved and control cells. To evaluate possible differences in the characteristics of the fibroblasts, the cells in both groups were tested for viability (membrane integrity), growth capacity and alkaline phosphatase (ALP) expression. The Wilcoxon test for paired comparison between cryopreserved and non-cryopreserved cells was performed for each characteristic. The results showed that membrane integrity of cells was not influenced by cryopreservation. There was no statistically significant difference in growth capacity between cryopreserved and control cells. Non-cryopreserved cells were slightly stronger positive for ALP, but the difference was not statistically significant. From these experiments it can be concluded that the observed parameters are not influenced by cryopreservation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Autotransplantation of teeth with partly formed roots seems to give better results than other treatment modalities for substituting missing teeth in a long-term perspective (Czochrowska et al. 2002; Zachrisson et al. 2004; Jonsson and Sigurdsson 2004).
Sometimes however, the ideal situation for autotransplantation is not always present: for example, when at the appropriate time of transplantation (root formation) the recipient site is too small and orthodontic treatment is required to gain space (Andreasen 1992). Another example is a clinical situation where extractions of premolars for orthodontic purposes are indicated in a patient with a traumatised incisor with an uncertain prognosis. In these cases cryopreservation of teeth may create new possibilities.
The aim of cryopreservation of living tissues is “controlled reversibility of the cessation of all biological functions” at an ultra low temperature. It is essential that no intracellular ice crystals are created. Shrinkage of cells during slow freezing is a result of egress of intracellular water, which is essential for the avoidance of cellular injury during the freezing process. Cryoprotective agents, such as dimethylsulfoxide (DMSO) or glycerol, are effective in counteracting cellular injury caused by freezing by reducing intracellular ice crystal formation and by reducing osmolarity stresses to cell membranes (Andreasen 1992).
Although several investigations concerning cryopreservation of teeth have been carried out in the past (Bartlett and Reade 1972; Price and Cserepfalvi 1972; Schwartz and Andreasen 1983; Schwartz et al. 1985; Schwartz and Rank 1986; Schwartz 1986; Politis et al. 1995; Laureys et al. 2001; Kawasaki et al. 2004), some questions remain unanswered concerning dental tissue reactions following cryopreservation. This is probably one of the reasons why transplantation of cryopreserved teeth is not widely accepted yet.
Periodontal healing is one of the most important factors determining success after autotransplantation (Kristerson 1985). It is generally known that the presence of a healthy and functional periodontal ligament (PDL) in immature teeth enhances the success rate after transplantation. The survival and function of the periodontal cells seem to be essential for the healing and prognosis of a tooth autograft (Melcher 1970; Line et al. 1974; Lindskog et al. 1983; Schwartz and Andreasen 1983; Schwartz et al. 1985; Kristerson 1985).
Cryopreservation does not seem to have a negative influence on the repair capacity of the PDL: in combination with a controlled freezing rate, the use of DMSO guarantees the survival of the majority of the periodontal cells (Kristerson et al. 1976; Politis et al. 1995; Schwartz et al. 2002; Oh et al. 2005). Therefore, the prognosis of autotransplanted cryopreserved teeth appears to be similar to that of directly transplanted and endodontically treated premolars and canines (Schwartz and Rank 1986; Andreasen 1992). This was confirmed by Kawasaki et al. (2004) in a study on rats: although the repair process proceeded slower, periodontal regeneration of cryopreserved teeth was substantially similar to that of the immediately transplanted teeth. There was no progressive root resorption, indicating that these cryopreserved teeth could be applicable for clinical use.
To date, the effect of cryopreservation on established cell cultures of PDL fibroblasts has not been studied yet.
The purpose of this research was to examine the effect of a standard cryopreservation procedure on human PDL cell cultures. Three variables were tested (membrane integrity (as an indicator of viability), growth capacity and alkaline phosphatase expression) in order to compare cryopreserved and control cells.
Materials and methods
Cell isolation and culture (Fig. 1)
Fibroblasts were obtained from the PDL of healthy extracted third molars of 11 subjects (between 15–30 years of age). Prior to extraction, informed consent was obtained. After extraction the teeth were stored in a carrier medium (DMEM 450 ml, Foetal Calf Serum (FCS) 19 ml, Raid solution (penicillin/streptomycin 10,000 U/ml, fungizone 5 mg/ml, and gentamycin 40 mg/ml) 1 ml) at 4°C for a maximum period of 24 h. The PDL attached to the middle third of the roots was cut from the root surface using a scalpel knife to avoid incorporation of gingival cells. After washing several times in phosphate buffered saline (PBS), the fragments of the PDL of each tooth were placed as explants in a 24-well culture plate and incubated with 1.5 ml culture medium (Optimem I supplemented with Ultroser G 2%, Glutamin 1%, penicillin/streptomycin 1%, FCS 5%) in a humidified atmosphere of 10% CO2 in air at 37°C. The different wells (two per patient) were considered as separate cell lines and all were treated individually. One cell line was lost due to bacterial contamination, resulting in a total of 21 cell lines. The culture medium was not changed until outgrowth of cells was seen. When the cells surrounding the tissue explants had become confluent (after a culture period of several weeks) they were trypsinised using 0.25% trypsine and 0.08% ethylenediamine tetraacetic acid (EDTA), transferred to a Petri dish (90 mm diameter), and designated as passage 1 (P1). The cells of P1 showed a typical fibroblastic shape. When confluence was reached the fibroblasts of each Petri dish were trypsinised and divided in two groups A and B (P2). Half of the cells (group A) was brought into FCS containing 10% DMSO (Schwartz et al. 1985), frozen at a slow freezing rate of 1°C/min from room temperature to −80°C followed by plunging in liquid nitrogen at −196°C. After storage in liquid nitrogen for one day the fibroblasts were rapidly thawed in a 37°C water bath, washed twice in Optimem and grown in Petri dishes. A quarter of the cells (group B) was immediately passaged to P2, without freezing. In order to investigate whether the cells were not merely gingival fibroblasts, the other quarter of cells was tested for alkaline phosphatase (ALP). ALP is an enzyme responsible for hard tissue formation present in most periodontal fibroblasts (Groeneveld et al. 1995), indicating that these cells are able to differentiate into osteoblasts and cementoblasts (Basdra and Komposch 1997). As most of the cells were strongly positive for ALP, it was assumed that the cells were indeed periodontal fibroblasts (Somerman 1988; Arceo et al. 1991; Giannopoulou and Cimasoni 1996; Gao et al. 1999). In order to justify the used technique, the gingival tissue of three patients was tested for ALP as a negative control in order to check the exclusive nature of the harvested periodontal tissue (Fig. 2).

Flow chart of the experimental set-up. (ALP = alkaline phosphatase; P0–P3 = cell culture passages)

Alkaline phosphatase (ALP) expression in fibroblasts of PDL (A) and gingiva (B). Gingival fibroblasts cultured from gingival tissue of three of the patients, served as a negative control. There is a strong contrast in ALP staining between PDL and gingival tissue. (Original magnification: ×160)
The cells in groups A and B reached confluence after 4–5 days of growth. At confluence, they were trypsinised and used for the experiments.
By using established cell cultures to start with for the different test settings, problems dealing with failing initiation of the cell cultures could be excluded. Hence, it could be anticipated that all the effects observed with the established cell cultures in the test conditions could be attributed to the cryopreservation procedure.
Experiments
Three variables were tested on the fibroblasts in both the cryopreserved group and the control group: viability (membrane integrity), growth capacity and ALP expression. For each variable a comparison was made between cryopreserved and non-cryopreserved cells to find out whether cryopreservation influenced the quality of human PDL cells. Cultured fibroblasts (P2) were trypsinised and the cell suspension was used for the experiments.
Viability/membrane integrity
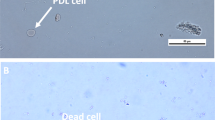
Exclusion of trypan blue is a measure of membrane integrity. An intact membrane is compatible with but does not guarantee viability of the cell. On the other hand, injury of the membrane and hence absence of membrane integrity is not compatible with the normal functions of a cell, except in the rare situation of recovery of membrane integrity. Therefore exclusion of trypan blue has often been associated with cell viability. The technique has been proposed by Bartlett and Reade (1972) to evaluate the viability of cryopreserved teeth. These authors described the trypan blue exclusion test as a simple and cost-effective procedure, which allows the detection of viable cells for subcultivation. Therefore, the trypan blue exclusion test was used in this study to isolate a certain amount of viable cells needed to evaluate growth capacity in the second part of the study.
A sample of the cell suspension was mixed 1:1 with trypan blue and the number of viable (no colouring) and non-viable (blue colouring) cells was counted using a Bürker chamber (Brand). The percentage of viable cells was used to compare groups A and B for each cell line.
Growth capacity
To define the most reliable initial amount of cells in the volume, a pilot test was carried out with cultures with an initial density of 104 and 105 cells. The cells in both groups were fixated after a certain period of time. Cell density assessment showed that the Petri dishes with an initial concentration of 105 cells gave the most reliable results after the chosen culture period. Therefore, this initial concentration was used for comparison between groups A and B.
A volume of cell suspension containing 105 viable cells was cultured in Petri dishes, for group A as well as for group B. After an equal period of time, when cells were subconfluent, the cryopreserved and non-cryopreserved fibroblasts (Fig. 3) were fixated using crystal violet (crystal violet 0.5%, formaldehyde 4%, absolute ethanol 30%, NaCl 0.17%). The Petri dishes were scanned and after background subtraction, a computerised cell density assessment was performed. Based on the number of pixels (as an indication for the number of cells) in the Petri dishes of groups A and B, the growth capacity of the cryopreserved and the control cells was evaluated (Fig. 4).

Periodontal ligament cells at subconfluence fixed with crystal violet. (Original magnification: ×160)

The Petri dishes were scanned and after background subtraction a computerised cell density assessment was performed. Based on the number of pixels (as an indication for the number of cells) present in the Petri dishes of groups A and B, growth capacity of the cryopreserved and the non-cryopreserved cells was evaluated. (U1 = fixated cryopreserved PDL cells; U2 = fixated non-cryopreserved PDL cells; U3 = empty Petri dish used as reference (B1 = background subtraction))
Alkaline phosphatase expression
Periodontal fibroblasts were scattered on a microscopic slide and Gomori alkaline phosphatase stain was used in order to quantify the ALP activity. The cells in groups A and B were scored twice by two independent and blinded persons. The scores ranged from 0 (no colouring) to 5 (very strong colouring). The scores in groups A and B were compared in order to find a possible influence of cryopreservation on the ALP activity, which could indicate an effect of cryopreservation on the differentiation capacity of the PDL cells.
Statistical analysis
Wilcoxon matched-pairs signed-ranks test was used for paired comparison between cells in groups A and B for each variable; association between both groups was expressed with the Spearman correlation coefficient. The error of the method was established using inter- and intra-observer weighted kappa (K) and intraclass correlation coefficient (ICC).
Results
Before dividing the confluent periodontal cells of P1 in groups A and B, they were tested for ALP, in order to check their true nature. In three patients, gingival fibroblasts had been tested for ALP as well and served as control. ALP scores for the gingival and periodontal fibroblasts were compared within the same patient. Only in one gingival sample, a very weak ALP score (score 1) was observed; whereas in the other two, the score was 0. ALP in periodontal fibroblasts always reached scores 4 or 5.
Viability (membrane integrity) of cells (Fig. 5)
The cells that had taken up trypan blue and the cells which excluded trypan blue were counted in order to define the percentage of viable cells. The mean of three counts (expressed in percentage) performed by one observer was used for the comparison between the cryopreserved and the control group. There was a weak correlation between both groups (Spearman correlation coefficient rS = 0.46, P = 0.04). The Wilcoxon test showed that there was no statistically significant difference in the percentage of viable cells (cells which excluded trypan blue) in the cryopreserved compared with the control group (P = 0.53).

Scatter diagram illustrating the correlation between the percentage of viable cells after culturing with and without cryopreservation
Growth capacity (Fig. 6)
The computerised cell density assessments were repeated in three differently defined areas within the same Petri dish. The ICC between the assessments was very high (ICC = 0.99). The paired Wilcoxon test revealed that there was no statistically significant difference in growth capacity between cryopreserved and control cells (P = 0.96). The Spearman correlation coefficient between the measurements of A and B was rS = 0.57 (P = 0.01).

Scatter diagram illustrating the correlation between the growth capacity after culturing with and without cryopreservation
Alkaline phosphatase expression (Fig. 7)
ALP staining was scored twice by two different observers. The inter and intraclass correlation coefficients and the weighted K (Table 1) according to Landis and Koch (1977) and Fleiss (1981) were assessed. The weighted K values ranged between 0.659 and 0.714 whereas the ICC varied between 0.877 and 0.935. There was a strong correlation between cryopreserved and control cells (rS = 0.70, P = 0.001). Although non-cryopreserved cells were slightly more positive for ALP, this difference was not statistically significant (P = 0.76).

Scatter diagram illustrating the correlation between the alkaline phosphatase (ALP) expression after culturing with and without cryopreservation. Point bins are used for the coinciding points by the adaptation of the diameter of the symbol
Discussion
The regeneration process after transplantation is complex, so it is difficult to draw conclusions about the influence of cryopreservation on the cell type involved. Schwartz et al. (1985) stated that the in vivo replantation technique precludes further extensive investigations into the optimisation of the cryoprofile of mature teeth. They concluded that it would be desirable to develop a simpler and more sensitive in vitro test system which would allow a more detailed study of cooling and freezing rates and the effect of cryoprotective agents on the cells involved. Therefore, the aim of this investigation was to examine the effect of a standardised cryopreservation procedure on periodontal fibroblasts in vitro.
Concerning viability, no statistically significant difference was found in the percentage of cells, which excluded trypan blue, in the cryopreserved group compared to the control group. Assessment of PDL cell viability after cryopreservation, using trypan blue, was already performed by Bartlett and Reade (1972). According to these authors it appeared that cryopreservation carried out under controlled conditions left the intricate intracytoplasmatic functions of the cells either undamaged or amenable to recovery. Kristerson et al. (1976) proved cell viability on the root surface of teeth by demonstrating that the cells could be detached from the teeth using trypsin after cryopreservation and a cultivation period. These findings also indicated that cryopreservation has no negative effect on the viability of PDL cells. Oh et al. (2005) came to the same conclusion in a study where MTT and Tunel assay were performed to examine the viability and apoptotic death of PDL cells on frozen and control PDL tissue. These authors also remarked that some cellular functions may remain transiently active in damaged cells (Stevenson et al. 2004) and consequently, the presence of viable cells does not ensure that cells are not damaged at all. In fact, membrane integrity testing of cells reveals the “status presens” of PDL cells at a certain moment of time. It does not offer an insight in the potential longitudinal properties of these cells. In order to study the effects of cryopreservation on PDL cells on the long term, it is more appropriate to assess the growth capacity of these cells.
To evaluate growth capacity, the technique used in this study was based on the technique to measure colony forming capacity (CFC). A fixed number of cells of a biopsy or at a certain passage is cultured using the current culture conditions. Colonies are fixed approximately 10 days later, stained with crystal violet and scored under a microscope. CFC values are expressed as the ratio of the number of colonies to the number of inoculated cells. In this study, colonies were not formed but cells were equally spread over the culture dish. Therefore it was decided to scan the plates and measure the density by means of the software program Quantity One (Bio-Rad).
Results show that there was no statistically significant difference in cell density between cryopreserved and control cells. These findings indicate that the procedure of freezing and thawing has no negative influence on the growth capacity of periodontal cells.
According to Basdra and Komposch (1997), cultured human PDL fibroblasts show an intense staining for ALP. This suggests that these cells have the potential to differentiate into osteoblasts and/or cementoblasts. In most of the investigations on PDL cultures in vitro, periodontal fibroblasts seemed to express ALP activity at a much higher level compared with gingival fibroblasts (Somerman et al. 1988; Arceo et al. 1991; Ogata et al. 1995; Giannopoulou and Cimasoni 1996; Gao et al. 1999; Kawasaki et al. 2004). However, some authors have found that gingival fibroblasts can express ALP activity, although to a lesser extent than PDL fibroblasts (Groeneveld et al. 1995; Carnes et al. 1997; Lallier et al. 2005). In the present study before comparing ALP staining between cryopreserved and non-cryopreserved cells (see materials and methods), gingival tissue was used as a negative control for ALP expression and compared to PDL in the same patients. The gingival fibroblasts in the present study were negative for ALP, except in one patient where ALP staining remained very weak compared to the PDL fibroblasts. This contrast in ALP scores between the cells obtained from both tissues, gave an indication that there was no incorporation of gingival cells in the cultures.
Quantification of the ALP staining was carried out in cryopreserved and control cells and evaluated by an intraclass correlation coefficient and a weighted K (Table 1). According to Landis and Koch (1977) values of K between 0.61 and 0.80 can be interpreted as substantial. According to Fleiss (1981) values of K between 0.41 and 0.75 can be interpreted as fair to good. In this study, the agreement between and within the observers can be expressed as good. The Wilcoxon test showed that non-cryopreserved cells were slightly stronger positive for alkaline phosphatase, but the difference was not statistically significant.
Oh et al. (2005) performed immunohistochemical staining for ALP to detect the differentiation capability of PDL cells after freezing and thawing. They showed ALP-positive cells in the PDL of control and cryopreserved teeth, without establishing a quantification of ALP activity in both groups. Kawasaki et al. (2004) observed, in their study on rats, that ALP activity in the PDL of cryopreserved teeth was still weak 2 weeks after transplantation. In this study, no statistically significant differences in ALP staining could be shown between cryopreserved and non-cryopreserved cells. It can be concluded that ALP activity of periodontal cells is not influenced by cryopreservation.
Every cryoprofile may be different to some extent. In a study on monkeys, Schwartz and colleagues (1985) showed that the best cryoprotectant was DMSO at a concentration of 10%, which was also used in the present study. For all the variables (membrane integrity, growth capacity and ALP expression) investigated, there was no difference between cryopreserved and control cells. The animal experiment of Kawasaki et al. (2004), and the in vitro investigation of Oh et al. (2005), using slightly different cryoprofiles, came to the same conclusion. Therefore, it seems that under controlled conditions, some small variations in the procedure of cryopreservation do not give different results. There seems to be no need to optimise the cryoprofile that has been used in this study.
Conclusion
According to the findings of this study there seems to be no difference in membrane integrity, growth capacity and ALP expression between cryopreserved and control periodontal cells. It can be concluded that cryopreservation under controlled conditions does not influence the viability nor the proliferation and differentiation capacity of human PDL cells in vitro. The standardised procedure used for cryopreservation meets the expectations and needs no further optimisation. Further studies are needed to find out whether or not other cell types are influenced by the cryopreservation procedure.
References
Andreasen JO (1992) Atlas of replantation and transplantation of teeth. Mediglobe SA, Fribourg (Switserland)
Arceo N, Sauk JJ, Moehring J, Foster RA, Somerman MJ (1991) Human periodontal cells initiate mineral-like nodules in vitro. J Periodontol 62:499–503
Bartlett PF, Reade PC (1972) Cryopreservation of developing teeth. Cryobiology 9:205–211
Basdra EK, Komposch G (1997) Osteoblast-like properties of human periodontal ligament cells:an in vitro analysis. Eur J Orthod 19:615–621
Carnes DL, Maeder CL, Graves DT (1997) Cells with osteoblastic phenotypes can be explanted from human gingiva and periodontal ligament. J Periodontol 68:701–707
Czochrowska EM, Stenvik A, Bjercke B, Zachrisson BU (2002) Outcome of tooth transplantation: survival and success rates 17–41 years posttreatment. Am J Orthod Dentofacial Orthop 121:110–119 quiz 193
Fleiss JL (1981) Statistical methods for rates and proportions. J Wiley & Sons
Gao J, Symons AL, Haase H, Bartold PM (1999) Should cementoblasts express alkaline phosphatase activity? Preliminary study of rat cementoblasts in vitro. J Periodontol 70:951–959
Giannopoulou C, Cimasoni G (1996) Functional characteristics of gingival and periodontal ligament fibroblasts. J Dent Res 75:895–902
Groeneveld MC, Everts V, Beertsen W (1995) Alkaline phosphatase activity in the periodontal ligament and gingiva of the rat molar: its relation to cementum formation. J Dent Res 74:1374–1381
Jonsson T, Sigurdsson TJ (2004) Autotransplantation of premolars to premolar sites. A long-term follow-up study of 40 consecutive patients. Am J Orthod Dentofacial Orthop 125:668–675
Kawasaki N, Hamamoto Y, Nakajima T, Irie K, Ozawa H (2004) Periodontal regeneration of transplanted rat molars after cryopreservation. Arch Oral Biol 49:59–69
Kristerson L (1985) Autotransplantation of human premolars. A clinical and radiographic study of 100 teeth. Int J Oral Surg 14:200–213
Kristerson L, Soder PO, Otteskog P (1976) Transport and storage of human teeth in vitro for autotransplantation and replantation. J Oral Surg 34:13–18
Lallier TE, Spencer A, Fowler MM (2005) Transcript profiling of periodontal fibroblasts and osteoblasts. J Periodontol 76:1044–1055
Landis JR, Koch GG (1977)The measurement of observer agreement for categorical data. Biometrics 33:159–174
Laureys W, Beele H, Cornelissen R, Dermaut L (2001) Revascularization after cryopreservation and autotransplantation of immature and mature apicoectomized teeth. Am J Orthod Dentofacial Orthop 119:346–352
Lindskog S, Blomlof L, Hammarstrom L (1983) Repair of periodontal tissues in vivo and in vitro. J Clin Periodontol 10:188–205
Line SE, Polson AM, Zander HA (1974) Relationship between periodontal injury, selective cell repopulation and ankylosis. J Periodontol 45:725–730
Melcher AH (1970) Repair of wounds in the periodontium of the rat. Influence of periodontal ligament on osteogenesis. Arch Oral Biol 15:1183–1204
Ogata Y, Niisato N, Sakurai T, Furuyama S, Sugiya H (1995) Comparison of the characteristics of human gingival fibroblasts and periodontal ligament cells. J Periodontol 66:1025–1031
Oh YH, Che ZM, Hong JC, Lee EJ, Lee SJ, Kim J (2005) Cryopreservation of human teeth for future organization of a tooth bank-A preliminary study. Cryobiology 51:322–329
Politis C, Vrielinck L, Schepers S, Lambrichts I (1995) Cryopreservation of teeth. Organizational aspects of a tissue bank for tooth tissues. Acta Stomatol Belg 92:149–154
Price PJ, Cserepfalvi M (1972) Pulp viability and the homotransplantation of frozen teeth. J Dent Res 51:39–43
Schwartz O (1986) Cryopreservation as long-term storage of teeth for transplantation or replantation. Int J Oral Maxillofac Surg 15:30–32
Schwartz O, Andreasen JO (1983) Cryopreservation of mature teeth before replantation in monkeys (I). Effect of different cryoprotective agents and freezing devices. Int J Oral Surg 12:425–436
Schwartz O, Andreasen FM, Andreasen JO (2002) Effects of temperature, storage time and media on periodontal and pulpal healing after replantation of incisors in monkeys. Dent Traumatol 18:190–195
Schwartz O, Andreasen JO, Greve T (1985) Cryopreservation before replantation of mature teeth in monkeys. (II). Effect of preincubation, different freezing and equilibration rates and endodontic treatment upon periodontal healing. Int J Oral Surg 14:350–361
Schwartz O, Rank CP (1986) Autotransplantation of cryopreserved tooth in connection with orthodontic treatment. Am J Orthod Dentofacial Orthop 90:67–72
Somerman MJ, Archer SY, Imm GR, Foster RA (1988) A comparative study of human periodontal ligament cells and gingival fibroblasts in vitro. J Dent Res 67:66–70
Stevenson DJ, Morgan C, Goldie E, Connel G, Grant MH (2004) Cryopreservation of viable hepatocyte monolayers in cryoprotectant media with high serum content: metabolism of testosterone and kaempherol post-cryopreservation. Cryobiology 49:97–113
Zachrisson BU, Stenvik A, Haanaes HR (2004) Management of missing maxillary anterior teeth with emphasis on autotransplantation. Am J Orthod Dentofacial Orthop 126:284–288
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Temmerman, L., Dermaut, L.R., De Mil, M. et al. Influence of cryopreservation on human periodontal ligament cells in vitro. Cell Tissue Banking 9, 11–18 (2008). https://doi.org/10.1007/s10561-007-9047-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10561-007-9047-y