
Abstract
The development of a primary tumor as such is not the main cause of death, but is rather the spreading of metastases, which causes over 90% of deaths in cancer patients. This largely depends on the ability of tumor cells to migrate away from the tumor and relocate at other areas of the body. Cell migration is known to be regulated by various extracellular signal substances such as neurotransmitters. However, before single tumor cells can start to invade into distant tissue, they have to dissociate from the primary tumor. This requires the disruption of cell-cell contacts, which are provided by a plethora of adhesion molecules like the family of cadherins. Using our well, established three-dimensional collagen-based cell migration assay, we show that engagement of N-cadherin results in a significant decrease of the spontaneous and the norepinephrine-induced migration of MDA-MB-468 breast carcinoma cells, which was due to an increase in the average break length. Moreover, this N-cadherin driven influence on the migratory activity is intracellularly integrated via multiple signaling pathways. Our results show that the impact of N-cadherin on the locomotion of MDA cells involves the activation of the adenylyl cyclase and the phosphatidylinositol-3-kinase (PI3K), but is independent of the protein kinase C (PKC) alpha. In summary, we provide evidence that the engagement of N-cadherin provides a stop signal for breast carcinoma cell migration, and accordingly the use of anti-N-cadherin antibodies or soluble ligands might be a tool to inhibit metastasis formation in E-cadherin negative but N-cadherin positive tumors.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Although it is well accepted that the formation of metastases is one of the most pernicious steps in the course of a cancer disease, still surprisingly few is known about the molecular mechanisms of the metastasis cascade. As part of this cascade, migration is one of the essential abilities, which a tumor cell has to acquire in order to be able to emigrate from the primary tumor and invade into distant tissues. We have reported previously in this journal that the migratory activity of MDA-MB-468 breast carcinoma cells is tightly regulated by a bunch of neurotransmitters, which are ligands to G protein-coupled receptors (GPCRs) [1]. Especially those neurotransmitters, which are released in stress reactions, i.e. norepinephrine, dopamine, and substance P, are potent initiators for tumor cell migration. With regard to the migratory activity and dynamics we can distinguish two types of migration: a slow, spontaneous type of migration is initiated by the extracellular matrix [2], and a fast type of locomotion is induced by extracellular substances such as ligands to GPCRs [1]. The intracellular signal transduction differs between these two types of migration. For example, only the GPCR-induced but not matrix-induced tumor cell locomotion depends on the activity of the phosphatidylinositol-3-kinase (PI3K) and type II myosin [3, 4].
With regard to the very beginning of the metastasis cascade the remaining question is, how the metastasizing tumor cells, which have gained migratory activity by GPCR-engagement, manage their disruption from the primary tumor. Cell-cell contacts are provided by a plethora of adhesion molecules. For the integrity of tissues, especially in epithelial tissues and correlating carcinomas, cadherins seem to be the most important of these adhesion molecules. Cadherins are a superfamily of homophilic adhesion molecules that mediate the cell–cell contacts in all solid tissues of the organism [5]. Among the cadherins, there are three types of the so-called classical cadherins, which are known to be important for normal and malignant breast tissue: the E (epithelial)-, P (placental)- and N (neural)-cadherin. E- and P-cadherin play a role in the architecture and function of the normal and mammary gland [6]. In tumors, downregulation of E-cadherin is correlated with a high migratory activity [7, 8], and consequently, in terms of the clinical impact, an aberrant expression of E-cadherin is associated with the development of metastases in patients with breast cancer [9]. In contrast, misexpression of P- and N-cadherin seems to increase the metastatic potential of breast tumors [6, 10]. On the intracellular side, these three classical cadherins are coupled to a protein complex that initiates several signal transduction pathways. So far, cadherins were reported to bind to β-catenin, which is bound to α-catenin, which in turn is associated with filamentous (f)-actin [5]. Recent findings discuss that α-catenin does not simultaneously interact with the E-cadherin-β-catenin complex and actin filaments [11, 12]. However, N-cadherin signaling activates the non-receptor tyrosine kinase Fer, which phosphorylates protein tyrosine phosphatase 1B (PTP1B) and the p120 catenin [13, 14]. PTP1B activity reduces the protein kinase C (PKC)δ activity, whereas it is not clear whether this is caused by a direct dephosphorylation of the PKCδ or the dephosphorylation of the phospholipase C (PLC)γ, which is the upstream activator for the PKCδ. In addition, homophilic cadherin ligation stimulates the activation of the PI3K [15]. However, the exact molecular mechanism through which the impact of N-cadherin signaling on the migration of tumor cells is intracellularly transduced and converted in force generation remains illusive.
In this current work we investigated the effect of N-cadherin on the migratory behavior of human MDA-MB-468 breast carcinoma cells and, more significantly, addressed potential signal transduction pathways underlying the N-cadherin-driven influence on the migration. Our experiments revealed that N-cadherin has a strong impact on the migratory activity of MDA-MB-468 breast carcinoma cells, and this impact is intracellularly integrated via multiple signaling pathways. These results can explain the ability of the metastasizing tumor cells to disrupt from the primary tumor, and deliver new access points to prevent this decisive step in the metastasis cascade.
Materials and methods
Cell culture
The human breast carcinoma cell line MDA-MB-468 (American Type Culture Collection, Rockville, MA, USA) was cultured in Leibovitz`s L-15 culture medium (PAA, Linz, Austria) containing 10% fetal calf serum (PAA) at 37°C humidified atmosphere.
Cell migration and data analysis
The migratory activity of the cells within three-dimensional collagen matrices was recorded by time-lapse videomicroscopy and analyzed by computer-assisted cell tracking [16]. Briefly, 9 × 104 tumor cells were mixed with 150 μl buffered liquid collagen (1.63 mg/ml collagen type I, pH 7.4; Nutacon, Leimuiden, Netherlands) containing minimal essential Eagle`s medium (Sigma, Deisenhofen, Germany) as well as the investigated substances. This mixture was filled into self-constructed migration chambers [4], and allowed to polymerize for 30 min at 37°C. After polymerization of the collagen, the chambers were sealed and the migration of the cells was then recorded for 10 h.
Migratory activity was induced by 10 μM norepinephrine, which is the strongest stimulus of carcinoma cell migration in our hands [1, 17, 18]. For the induction of N-cadherin signaling we used the well characterized monoclonal anti-N-cadherin antibody GC-4 (150 μg/ml; Sigma). This antibody blocks the homotypic binding of N-cadherin [19, 20], and induces intracellular signaling [21, 22]. As a negative control we used the monoclonal isotype control IgG1 clone 11,711.11 (R&D Systems, Wiesbaden, Germany). Furthermore, we used the PKC activator phorbol-12-myristate-13-acetate (PMA; 5 ng/ml; Sigma), the PI3K inhibitor wortmannin (250 nM), the PKCα inhibitor Gö6976 (60 nM), the PKCδ inhibitor rottlerin (15 μM), the PLCγ inhibitor U73122 (250 ng/ml), the adenylyl cyclase activator forskolin (10 μM), and the protein kinase A inhibitor H-89 (500 nM; all substances were derived from Calbiochem-Merck, Bad Soden, Germany). None of the substances or antibodies affected the viability of the cells at the concentrations used, as was analyzed by propidium iodide staining and flow cytometry.
For analysis of the migratory activity, 30 cells were randomly selected and the paths were digitized in 15 min intervals by computer-assisted cell-tracking. The percentage of locomoting cells was calculated for every interval as the moving part of the thirty analyzed cells. Furthermore, migrating tumor cells migrate not continuously, but they perform stop periods. From the individual path analysis, we calculated the frequency and duration of these breaks between locomotory active phases [2]. Statistical analyses were performed using the Student`s t-test.
Flow cytometry
The expression of cadherins was determined using a FacsCalibur flow cytometer (Becton Dickinson, Heidelberg, Germany). After fixation with 4% formaldehyde, we incubated 3 × 105 cells for 10 min at room temperature with 10 μg/ml of the primary antibody (67A4 for E-cadherin; Coulter-Immunotech, Hamburg, Germany; the anti-P-cadherin antibody was derived from Zymed, San Francisco, CA; the anti-N-cadherin antibody GC-4 was derived from Sigma as described above). After washing we incubated the cells with 10 μg/ml of fluorescein isothiocyanate (FITC)-conjugated anti-mouse antibody (Dianova, Hamburg, Germany). Non-specific binding was determined by isotypic control mouse antibody (Coulter-Immunotech).
Adhesion assay
For investigation of cell adhesion, a 24-well plate (TPP, Trasadingen, Switzerland) was coated with 250 μl collagen solution/well. Following polymerization for 1 h at 37°C, medium containing 2.5 × 105 tumor cells, and the investigated substances were applied on the collagen lattices and allowed to adhere for 4 h at 37°C in a humidified atmosphere. Subsequently, the non-adherent cells were collected by washing the plate three times with 500 μl PBS, centrifuged and resuspended in 300 μl PBS. The adherent cells were assembled after digestion of the collagen gels with collagenase for 1 h at 37°C and one washing step with PBS. For analysis cells were counted for a 20 s interval using flow cytometry. Cell adhesion was enumerated as the ratio of cells bound to the collagen lattice to unbound cells in the supernatant and then evaluated relative to control. Each experiment was carried out in duplicate and repeated three times.
Results
N-cadherin expression and its function in MDA-MB-468 breast carcinoma cell migration
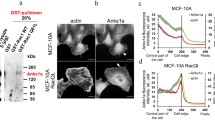
We have investigated the expression of E-, P-, and N-cadherin on the surface of MDA-MB-468 breast carcinoma cells by flow cytometry. All three cadherins are expressed on these cells as confirmed by western blot analysis (data not shown), whereas the expression of N-cadherin was most prominent (Fig. 1a). After incorporation into a three-dimensional collagen matrix, 45.5 ± 8.4% of the MDA cell were spontaneously locomotory active without any further stimulus (Fig. 1b). After addition of 10 μM norepinephrine, the locomotory activity significantly (P = 0.01) increased to 68.2 ± 5.7% of the cells. Treatment with 150 μg/ml of the monoclonal anti-N-cadherin antibody GC-4, which recognizes the extracellular domain of N-cadherin, significantly reduced this norepinephrine-induced migration to 33.6 ± 17.2% locomoting cells (P = 0.03; Fig. 1b). In addition, treatment with CG4 resulted in a significant decrease of the spontaneous migratory activity to 33.6 ± 14.2% migrating cells (P = 0.02) compared with the IgG-treated control (Fig. 1b).

a Expression of various cadherins on MDA-MB-468 breast carcinoma cells. The FITC-fluorescence of the cadherin-specific antibodies (gray area) was compared to an unspecific isotype control (black line). MFI, mean fluorescence intensity of specific and isotypic (Iso) binding. b The migration of the cells was induced by 10 μM norepinephrine and significantly inhibited by GC4-N-cadherin-antibody (150 μg/ml; anti-N-cad; P = 0.02). As a negative control cells were treated with IgG1 isotype control antibody (150 μg/ml, IgG). The graph shows mean values of four independent experiments (120 cells were analyzed per sample). c Treatment of the cells with anti-N-cadherin-antibody significantly increased their average break length during migration in the collagen matrix (P ≤ 0.021). An overview on additional migratory parameters and the number of analyzed cells is summarized in Table 1
Further analysis of the migration behavior showed that these effects of the GC-4 antibody were largely due to a significant increase of the average break length (Fig. 1c). As described in the Materials and methods section, tumor cells migrate not continuously, but they perform stop periods in a characteristic manner [2]. After N-cadherin engagement with the GC-4 antibody, the break length of spontaneously migrating cells significantly (P = 0.016) increased from 72.9 ± 13.0 to 102.0 ± 16.9 min, and of the norepinephrine-induced migration increased from 50.6 ± 14.4 to 101.5 ± 39.6 min (P = 0.021; Fig. 1c), whereas the frequency of breaks was only marginally lower as summarized in Table 1.
The signal transduction of N-cadherin with respect to migration
We have shown previously that changes in the migratory pattern are also accompanied by differences in the underlying intracellular signaling pathways [17, 23, 24]. Our results show that cells treated with an anti-N-cadherin antibody decreased the norepinephrine-stimulated migration as well as the spontaneous locomotion of MDA cells. These two types of carcinoma cell migration depend on the activity of the PLCγ by its tyrosine phosphorylation [17, 25]. Accordingly, treatment of the MDA cells with the PLCγ inhibitor U73122 (250 ng/ml) reduced the migratory activity to the same extent as N-cadherin engagement, from 51.3 ± 10.5% migrating cells of the control to 41.1 ± 10.7% migrating cells after treatment with the N-cadherin antibody and to 39.1 ± 9.6% migrating cells after treatment with U73122 (Fig. 2a). Strikingly, U73122 did not aggravate the effect of the N-cadherin antibody (46.0 ± 5.8% migrating cells).

Involvement of the PKC and the PLCγ activity in the N-cadherin mediated reduction of MDA-MB-468 cell migration. The PLCγ was inhibited by adding 250 ng/ml U73122 (a) and the PKC was activated by 5 ng/ml PMA (b; P = 0.03). The PKCα was inhibited using the PKCα-specific inhibitor Gö6976 (60 nM) and did not have any effect on MDA cell migration (c). The PKCδ was inhibited by rottlerin (15 μM) and resulted in a significant decrease of the spontaneous (P < 0.0001) and the N-cadherin mediated tumor cell locomotion (d; P = 0.002). The graphs show mean values of three independent experiments (90 cells were analyzed per sample). None of the substances reduced the viability of the cells as was assessed by propidium-iodide staining and flow cytometry
The PLCγ produces two second messenger molecules, diacylglycerol and inositol-triphosphate (IP3). Diacylglycerol is an activator for classical and novel PKC isotypes. We substituted diacylglycerol by its analog PMA, which led to a strong increase of migratory activity in these cells from 59.2 ± 0.5% migrating cell of the control to 67.5 ± 9.5% migrating cells after treatment with 5 ng/ml PMA (P = 0.03; Fig. 2b). This effect was not influenced by anti-N-cadherin antibody treatment, suggesting that a direct activation of PKCs can overcome their inactivation by a decrease of the diacylglycerol production, which is caused by the N-cadherin-mediated inactivation of the PLCγ. Which isotype is in these cells responsible for the regulation of migratory activity? From our own previous investigations and inquests on this topic we know that both the PKCα and the PKCδ are described to be involved in tumor cell migration [8], whereas the PKCδ negatively regulates the PKCα-activity via phosphorylation of the adapter-molecule syndecan-4 [26, 27]. Both of the PKC isotypes are expressed in MDA-MB-468 cells, as was analyzed by immunoblotting (data not shown). However, inhibition of the PKCα with 60 nM Gö6976 did not have any effect neither on the spontaneous, matrix-induced migration nor on the already decreased locomotion initiated by N-cadherin-antibody treatment (Fig. 2c). In contrast, inhibition of the PKCδ with 15 μM rottlerin significantly (P < 0.001) reduced the spontaneous migratory activity from 49.3 ± 19.3% migrating cell to 21.3 ± 2.7% migrating cells, and even further decreased the locomotor activity of cells incubated with the anti-N-cadherin antibody from 36.5 ± 3.7% migrating cells to 8.6 ± 5.6 after the combined incubation with the antibody and rottlerin (Fig. 2d); P = 0.002). These results suggest a differential involvement of these two PKC isotypes in the migration of MDA-MB-468 breast carcinoma cells: PKCα activity is not involved in the N-cadherin-antibody mediated decrease of locomotion, whereas PKCδ activity is inherently necessary for the spontaneous, matrix-induced migration.
Another signaling pathway known to be involved in the regulation of cell migration is transduced via the adenylyl cyclase, an enzyme which is usually activated during GPCR signaling [25]. The adenylyl cyclase generates cyclic AMP (cAMP), which in turn is an activator for the protein kinase A (PKA). The PKA plays a crucial role in cytoskeletal regulation by inducing actin filament assembly or activation of Rac, Cdc42 or Rho [28]. Herein, activation of the adenylyl cyclase using 10 μM forskolin resulted in a decrease of the spontaneous migration to the same extent as N-cadherin engagement from 55.85 ± 10.53% locomoting cells of the control to 37.06 ± 9.05% migrating cells after incubation with the anti-N-cadherin antibody (P = 0.033), and 38.48 ± 11.4% locomotory active MDA cells after treatment with forskolin (Fig. 3a). Stimulation of the adenylyl cyclase did not further augment the decrease of the migratory activity induced by antibody treatment. Incubation of the cells with GC-4 antibody was accompanied by an increase of the cytosolic cAMP concentration (data not shown).

Role of the adenylyl cyclase and the PKA in the N-cadherin mediated decrease of MDA cell migration. a The activity of the adenylyl cyclase was stimulated by adding 10 μM forskolin. b The PKA was inhibited by treatment of the cells with 500 nM of the specific inhibitor H-89. The graphs show mean values of four independent experiments (120 cells were analyzed per sample)
Inhibition of the PKA using 500 nM H-89 significantly reduced the spontaneous migratory activity of MDA cells from 59.8 ± 5.49% locomotory active cells to 45.3 ± 1.69% migrating active cells after incubation with the specific PKA inhibitor (P = 0.002; Fig. 3b). Moreover, the N-cadherin mediated decrease of tumor cell migration remained almost unchanged after additional treatment with H-89 (from 39.4 ± 9.76% locomoting after incubation with anti-N-cadherin antibody to 46.5 ± 6.9% migrating cells after incubation with H-89 in combination with the GC-4 antibody). Thus, we assume that N-cadherin signaling involves an activation of the adenylyl cyclase, but is independent from PKA activity.
As we have investigated previously, only the norepinephrine-induced, but not the spontaneous migration of MDA-MB-468 cells depends on the activity of the PI3K [3]. Here, inhibition of the PI3K with 250 nM wortmannin abolished the significant (P = 0.043) reduction caused by the anti-N-cadherin antibody GC-4 (Fig. 4): as compared to the control (60.7 ± 10.7% locomoting cells), 41.3 ± 8.5% of the cells migrated after GC-4 antibody treatment, but 61.2 ± 6.3% of the cells migrated after treatment with the antibody and wortmannin in combination (P = 0.02). Wortmannin alone had no effect (55.7 ± 4.0% locomoting cells). In conclusion, the PI3K is activated by N-cadherin signaling.

Involvement of the PI3K in the N-cadherin mediated reduction of the spontaneous locomotory activity of MDA cells. The PI3K was inhibited with 250 nM wortmannin. The graph shows mean values of three independent experiments (90 cells were analyzed per sample)
N-cadherin engagement causes an increase of adhesion, which is not mediated by integrins
There are several lines of evidence that PKCs regulate the integrin-mediated adhesion, and the PKCα and δ are associated with focal adhesions [29]. Furthermore, the PKCα regulates β1–integrin dependent motility [30]. In turn, β1–integrin signaling is a regulator for the phosphorylation and thereby activity of the PKCδ [31]. Especially β1–integrins are important mediators of cells adhesion to extracellular matrix proteins such as collagen and are essential for the migration of tumor cells [32]. In order to investigate what the dominant reduction of migratory activity by N-cadherin engagement might further be based on, we analyzed the adhesion of the MDA cells to collagen (Fig. 5). MDA cells treated with GC-4 antibody had a significantly increased ability to adhere to the collagen matrix (P = 0.02; Fig. 5). However, this enhanced adhesion was not mediated by integrins, because incubation of the cells with anti-N-cadherin antibody did not have any effect on the phosphorylation status of β1–integrins (data not shown).

Influence of various substances on the adhesion of MDA-MB-468 cells to the collagen matrix. The graph show mean values of three independent experiments. P ≤ 0.02 as indicated by an asterisk
Discussion
N-cadherin plays a role in MDA-MB-468 tumor cell migration
The process of metastasis formation consists of a long series of sequential, interrelated steps. Before single tumor cells can start to actively migrate to new sites within the body, they have to dissociate from the primary tumor. This requires the disruption of cell-cell contacts, which are provided by a plethora of adhesion molecules such as the family of cadherins. Downregulation of E-cadherin was found to correlate with a higher metastatic potential [8, 33], and is often correlated with an up-regulation of N-cadherin, an invasion promoter molecule [34]. Moreover, several studies discuss a functional cooperation between N-cadherin and members of the fibroblast growth factor receptor family, resulting in increased cellular motility and invasion [13]. We have shown previously that norepinephrine is a strong stimulator for the chemokinetic and chemotactic locomotion of MDA-MB-468 breast carcinoma cells [1]. However, with regard to the very beginning of the metastasis cascade, the present study establishes a key role of N-cadherin in the regulation of MDA breast carcinoma cell migration. Treatment of the cells with an anti-N-cadherin blocking antibody resulted in a significant decrease of the spontaneous as well as the norepinephrine-induced migration. This inhibitory effect on the locomotory activity of MDA cells was due to an increase in the average length of breaks. In adhesion assays the GC-4 antibody engagement significantly increased the adhesion of MDA cells to the collagen matrix. In this context, previous studies of the migratory pattern have shown that the norepinephrine-stimulated decrease in break length during tumor cell migration was attributed to the fact that the initial steps in cell migration, namely pseudopod development and matrix adhesion, take place faster [2]. Therefore, the herein observed increase in the length of breaks might originate from an elevated adhesion ability of the cells to the surrounding matrix, thus resulting in an enhanced retention period of the tumor cells in place because of a decelerated dissociation from the matrix. This might be the result of a decrease in the enzymatic activity of the proteins regulating the actin cytoskeleton, as we will discuss in the following.
Several pathways are involved in N-cadherin signaling
The signal pathways activated by cadherins have been under great deal of study [34, 35], whereas the molecular mechanisms, which intracellularly transduce the inhibitory effect of N-cadherin signaling on cell migration, are not entirely clear. With regard to the stimuli of cell migration and to the migratory dynamics, we can distinguish between a slow type of locomotion, which is initiated by the matrix, and a faster, more active type of migration, which is induced by soluble substances. Both types of migration are differentially regulated and various signaling molecules have been detected with a known function during the locomotory process [25]. The activity of the PLCγ is a decisive feature for both types of locomotion [17, 25]. In our study, inhibition of the PLCγ reduced the migratory activity to the same extent as N-cadherin engagement, and did not aggravate the effect of the N-cadherin antibody. Thus it is likely, that the signal pathway transducing the N-cadherin mediated inhibition of cell migration involves an inactivation of the PLCγ.
In general, the PLCγ catalyzes the hydrolysis of phosphatidyl-inositol-bisphosphate (PIP2) to DAG, an activator of classical and novel PKC isotypes, and inositol-triphosphate a second messenger, which opens intracellular calcium channels [17]. The PKC has been shown to regulate a plethora of target molecules essential for tumor cell migration, including the actin-binding proteins MARCKS and gelsolin [3] and the beta-chain of integrins [27]. Stimulating the cells with the DAG analog PMA strongly enhanced the locomotory activity of MDA cells, and this promigratory effect was not influenced by N-cadherin engagement. Therefore we suggest that a direct activation of the PKCs is sufficient to compensate their inactivation by a decrease of DAG as a result of N-cadherin mediated PLCγ dephosphorylation. Furthermore, N-cadherin signaling has been described to stimulate the activity of PTP1B [13]. PTP1B in turn has been shown to reduce the PKCδ activity [36], whereas it is not clear whether this is caused by a direct dephosphorylation of the PKCδ or the dephosphorylation of the PLCγ. However, our interpretation that an inactivation of the PLCγ and the PKCδ is involved in the N-cadherin mediated decrease of the migratory activity is further supported by migration experiments with PKC isotype specific inhibitors. Whereas treatment of the cells with the PKCα specific inhibitor Gö6976 did not have any effect on the locomotory behavior of the cells, inhibition of the PKCδ activity significantly decreased the spontaneous migration and further aggravated the decreased locomotion of MDAs after N-cadherin engagement.
A second distinct pathway, which is of special importance for tumor cell migration is transduced via the adenylyl cyclase, an enzyme which is usually activated during GPCR signaling [23, 25]. Activation of the adenylyl cyclase leads to an increase of cAMP, which mediates its action through the PKA and the exchange protein directly activated by cAMP (Epac). The PKA phosphorylates a plethora of downstream targets involved in the regulation of migration. For example, the PKA participates in the regulation of the cytosolic calcium concentration via phospholamban, an inhibitory protein of the sarcoplasmatic/endoplasmatic calcium ATPase, and actin assembly via action on Ena/VASP proteins and profilin [37, 38]. Epac is also involved in the control of cell migration via an activation of Rap1 GTPase [39]. Thus the observed decrease of migration after treatment of the cells with GC-4 antibody is dependent on the adenylyl cyclase, but independent of the PKA. Instead, cAMP seems to mediate its influence on the migratory activity via Epac and Rap1. Furthermore treatment of MDA cells with GC-4 antibody also resulted in an activation of the PI3K because the N-cadherin mediated decrease of MDA cell migration was abrogated by using wortmannin, a specific inhibitor of the PI3K. This is consistent with previous findings demonstrating that the migratory activity of MDA-MB-468 breast carcinoma cells was impaired by the inhibition of PI3K [3].
To sum up, our data show that the adenylyl cyclase as well as the PI3K are both involved in the migration of MDA-MB-468 cells and the N-cadherin mediated signaling, whereas the exact molecular regulation is not entirely clear.
In conclusion, we herein deliver evidence that tumor cells are not in all respects resistant to inhibitory cell-cell contact signals. Although it is clear that the proliferation is not sensitive to such contact signals, we show that the migration is still potentially sensitive and that these signals are dominant stop signals for migration. However, the question still is how tumor cells can emigrate from a primary tumor despite these inhibitory signals. It might well be, that the homotypic binding of N-cadherin alone is not strong enough to deliver a sufficient stop signal, which is able to override the migration inducing signal from neurotransmitters and chemokines. The antibody used in this study has a much higher affinity to N-cadherin and thereby provides a sufficient signal. Thus, the use of anti-N-cadherin antibodies or soluble ligands might be a tool to inhibit metastasis formation in E-cadherin negative but N-cadherin positive tumors.
References
Drell TL, Joseph J, Lang K, Niggemann B, Zaenker KS, Entschladen F (2003) Effects of neurotransmitters on the chemokinesis and chemotaxis of MDA-MB-468 human breast carcinoma cells. Breast Cancer Res Treat 80:63–70
Niggemann B, Drell TL, Joseph J, Weidt C, Lang K, Zaenker KS, Entschladen F (2004) Tumor cell locomotion: differential dynamics of spontaneous and induced migration in a 3D collagen matrix. Exp Cell Res 298:178–187
Bastian P, Posch B, Lang K, Niggemann B, Zaenker KS, Hatt H, Entschladen F (2006) Phosphatidylinositol 3-kinase in the G protein-coupled receptor-induced chemokinesis and chemotaxis of MDA-MB-468 breast carcinoma cells: a comparison with leukocytes. Mol Cancer Res 4:411–421
Bastian P, Lang K, Niggemann B, Zaenker KS, Entschladen F (2005) Myosin regulation in the migration of tumor cells and leukocytes within a three-dimensional collagen matrix. Cell Mol Life Sci 62:65–76
Ivanov DB, Philippova MP, Tkachuk VA (2001) Structure and functions of classical cadherins. Biochemistry (Mosc) 66:1174–1186
Knudsen KA, Wheelock MJ (2005) Cadherins and the mammary gland. J Cell Biochem 95:488–496
Behrens J (1993) The role of cell adhesion molecules in cancer invasion and metastasis. Breast Cancer Res Treat 24:175–184
Masur K, Lang K, Niggemann B, Zanker KS, Entschladen F (2001) High PKC alpha and low E-cadherin expression contribute to high migratory activity of colon carcinoma cells. Mol Biol Cell 12:1973–1982
Kowalski PJ, Rubin MA, Kleer CG (2003) E-cadherin expression in primary carcinomas of the breast and its distant metastases. Breast Cancer Res 5:R217–R222
Nagi C, Guttman M, Jaffer S, Qiao R, Keren R, Triana A, Li M, Godbold J, Bleiweiss IJ, Hazan RB (2005) N-cadherin expression in breast cancer: correlation with an aggressive histologic variant??? invasive micropapillary carcinoma. Breast Cancer Res Treat 94:225–235
Drees F, Pokutta S, Yamada S, Nelson WJ, Weis WI (2005) Alpha-catenin is a molecular switch that binds E-cadherin-beta-catenin and regulates actin-filament assembly. Cell 123:903–915
Yamada S, Pokutta S, Drees F, Weis WI, Nelson WJ (2005) Deconstructing the cadherin-catenin-actin complex. Cell 123:889–901
Cavallaro U, Christofori G (2004) Cell adhesion and signalling by cadherins and Ig-CAMs in cancer. Nat Rev Cancer 4:118–132
Lilien J, Balsamo J (2005) The regulation of cadherin-mediated adhesion by tyrosine phosphorylation/dephosphorylation of beta-catenin. Curr Opin Cell Biol 17:459–465
Kovacs EM, Ali RG, McCormack AJ, Yap AS (2002) E-cadherin homophilic ligation directly signals through Rac and phosphatidylinositol 3-kinase to regulate adhesive contacts. J Biol Chem 277:6708–6718
Entschladen F, Drell TL, Lang K, Masur K, Palm D, Bastian P, Niggemann B, Zaenker KS (2005) Analysis methods of human cell migration. Exp Cell Res 307:418–426
Masur K, Niggemann B, Zanker KS, Entschladen F (2001) Norepinephrine-induced migration of SW 480 colon carcinoma cells is inhibited by beta-blockers. Cancer Res 61:2866–2869
Lang K, Drell TL, Lindecke A, Niggemann B, Kaltschmidt C, Zaenker KS, Entschladen F (2004) Induction of a metastatogenic tumor cell type by neurotransmitters and its pharmacological inhibition by established drugs. Int J Cancer 112:231–238
Goncharova EJ, Kam Z, Geiger B (1992) The involvement of adherens junction components in myofibrillogenesis in cultured cardiac myocytes. Development 114:173–183
Puch S, Armeanu S, Kibler C, Johnson KR, Muller CA, Wheelock MJ, Klein G (2001) N-cadherin is developmentally regulated and functionally involved in early hematopoietic cell differentiation. J Cell Sci 114:1567–1577
Matsumura T, Wolff K, Petzelbauer P (1997) Endothelial cell tube formation depends on cadherin 5 and CD31 interactions with filamentous actin. J Immunol 158:3408–3416
Makrigiannakis A, Coukos G, Christofidou-Solomidou M, Gour BJ, Radice GL, Blaschuk O, Coutifaris C (1999) N-cadherin-mediated human granulosa cell adhesion prevents apoptosis: a role in follicular atresia and luteolysis? Am J Pathol 154:1391–1406
Joseph J, Niggemann B, Zaenker KS, Entschladen F (2002) The neurotransmitter gamma-aminobutyric acid is an inhibitory regulator for the migration of SW 480 colon carcinoma cells. Cancer Res 62:6467–6469
Lang K, Niggemann B, Zanker KS, Entschladen F (2002) Signal processing in migrating T24 human bladder carcinoma cells: role of the autocrine interleukin-8 loop. Int J Cancer 99:673–680
Palm D, Lang K, Brandt B, Zaenker KS, Entschladen F (2005) In vitro and in vivo imaging of cell migration: two interdepending methods to unravel metastasis formation. Semin Cancer Biol 15:396–404
Horowitz A, Tkachenko E, Simons M (2002) Fibroblast growth factor-specific modulation of cellular response by syndecan-4. J Cell Biol 157:715–725
Murakami M, Horowitz A, Tang S, Ware JA, Simons M (2002) Protein kinase C (PKC) delta regulates PKCalpha activity in a Syndecan-4-dependent manner. J Biol Chem 277:20367–20371
Howe AK (2004) Regulation of actin-based cell migration by cAMP/PKA. Biochim Biophys Acta 1692:159–174
Miranti CK, Ohno S, Brugge JS (1999) Protein kinase C regulates integrin-induced activation of the extracellular regulated kinase pathway upstream of Shc. J Biol Chem 274:10571–10581
Ng T, Shima D, Squire A, Bastiaens PI, Gschmeissner S, Humphries MJ, Parker PJ (1999) PKCalpha regulates beta1 integrin-dependent cell motility through association and control of integrin traffic. Embo J 18:3909–3923
Parekh DB, Katso RM, Leslie NR, Downes CP, Procyk KJ, Waterfield MD, Parker PJ (2000) Beta1-integrin and PTEN control the phosphorylation of protein kinase C. Biochem J 352(Pt 2):425–433
Maaser K, Wolf K, Klein CE, Niggemann B, Zanker KS, Brocker EB, Friedl P (1999) Functional hierarchy of simultaneously expressed adhesion receptors: integrin alpha2beta1 but not CD44 mediates MV3 melanoma cell migration and matrix reorganization within three-dimensional hyaluronan-containing collagen matrices. Mol Biol Cell 10:3067–3079
Mareel M, Vleminckx K, Vermeulen S, Bracke M, Van Roy F (1992) E-cadherin expression: a counterbalance for cancer cell invasion. Bull Cancer 79:347–355
Hazan RB, Qiao R, Keren R, Badano I, Suyama K (2004) Cadherin switch in tumor progression. Ann N Y Acad Sci 1014:155–163
Fricke E, Keller G, Becker I, Rosivatz E, Schott C, Plaschke S, Rudelius M, Hermannstadter C, Busch R, Hofler H, Becker KF, Luber B (2003) Relationship between E-cadherin gene mutation and p53 gene mutation, p53 accumulation, Bcl-2 expression and Ki-67 staining in diffuse-type gastric carcinoma. Int J Cancer 104:60–65
Benes C, Soltoff SP (2001) Modulation of PKCdelta tyrosine phosphorylation and activity in salivary and PC-12 cells by Src kinases. Am J Physiol Cell Physiol 280:C1498–C1510
Jockusch BM, Bubeck P, Giehl K, Kroemker M, Moschner J, Rothkegel M, Rudiger M, Schluter K, Stanke G, Winkler J (1995) The molecular architecture of focal adhesions. Annu Rev Cell Dev Biol 11:379–416
Kwiatkowski AV, Gertler FB, Loureiro JJ (2003) Function and regulation of Ena/VASP proteins. Trends Cell Biol 13:386–392
Yarwood SJ (2005) Microtubule-associated proteins (MAPs) regulate cAMP signalling through exchange protein directly activated by cAMP (EPAC). Biochem Soc Trans 33:1327–1329
Acknowledgments
We thank Gaby Troost for excellent technical assistance. This work was supported by the Fritz Bender Foundation (Munich, Germany), and the German Cancer Aid (Deutsche Krebshilfe, Bonn, Germany).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Potthoff, S., Entschladen, F., Niggemann, B. et al. N-cadherin engagement provides a dominant stop signal for the migration of MDA-MB-468 breast carcinoma cells. Breast Cancer Res Treat 105, 287–295 (2007). https://doi.org/10.1007/s10549-006-9458-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10549-006-9458-7