
Abstract
What changes when an evolutionary transition in individuality takes place? Many different answers have been given, in respect of different cases of actual transition, but some have suggested a general answer: that a major transition is a change in the extent to which selection acts at one hierarchical level rather than another. The current paper evaluates some different ways to develop this general answer as a way to characterise the property ‘evolutionary individuality’; and offers a justification of the option taken in Clarke (J Philos 110(8):413–435, 2013)—to define evolutionary individuality in terms of an object’s capacity to undergo selection at its own level. In addition, I suggest a method by which the property can be measured and argue that a problem which is often considered to be fatal to that method—the problem of ‘cross-level by-products’—can be avoided.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Philosophers have become much interested in the question of what sorts of biological things have the property ‘individuality’ (Hull 1978; Wilson 1999; De Sousa 2005; Wilson and Barker 2013; Clarke 2010; Martens 2010; Bouchard and Huneman 2013; Guay and Pradeu 2015). In addition to the long-standing debate about whether species should be thought of as particulars or as classes (Ghiselin 1974; Hull 1978), more recent attention has focused on the individuality of units at a lower compositional level. Bigger than organs, but smaller than populations, ‘biological individuals’ are in some ways the most obvious of biological particulars. Various biological processes have been suggested as picking out important kinds of biological particulars (for example, immunogenicity (Pradeu 2010) and metabolism (Dupré and O’Malley 2009)). And special attention has been focused on the ontological status of various, non-standard, candidate individuals, such as fungal hyphae (Booth 2014); insect colonies (Haber 2013); and bacterial biofilms (Ereshefsky and Pedroso 2015).
One important strand of debate, in both biology and philosophy, aims to say something general about which biological objects are treated as individuals by the process of natural selection (Lewontin 1970; Janzen 1977; Santelices 1999; Gould and Lloyd 1999; Queller 2000; Bouchard 2008; Pepper and Herron 2008; Queller and Strassmann 2009; Gardner and Grafen 2009; Godfrey-Smith 2009; Folse and Roughgarden 2010; Clarke 2013). This debate developed out of the controversy about group selection and the Major Transitions in Evolution. In the 1960s the consensus was that natural selection acts on individual organisms, rather than at higher, group, levels (Williams 1966). But according to a classical view, the properties necessary for evolution by natural selection can occur at any hierarchical level, at least as a matter of logic (Lewontin 1970). The key insight of the Major Transitions literature was that individual organisms, such as humans, are themselves higher-level individuals: groups of cells that somehow transitioned to being true individuals in their own right (Margulis 1970; Bonner 1974; Buss 1987; Maynard Smith and Szathmary 1997; Okasha 2006). Given this, Wilson and Sober argued that it makes no sense to resist the notion of group selection—we know that groups of cells can be selected, in the human case. The remaining question is just what other sorts of groups can be selected (Wilson and Sober 1989). Those who try to define the ‘evolutionary individual’ may be thought of as trying to say just what properties any sort of group needs to have so that it, too, can be selected, in the same way that humans are selected.
The quest to define this ‘evolutionary individual’ has some practical consequences for evolutionary theory, because the evolutionary individual is the bearer of fitness: the unit of currency, as it were, in which evolutionary change is routinely calculated. We talk of the relative frequencies of wrinkly versus smooth types of pea, for example, where one type is more frequent if there are a greater number of individuals—plants—that express that type. Or we talk of gene frequencies where, again, we mean that one allele occurs at a greater frequency than another if it is carried by a greater number of individual organisms. The simplest versions of evolutionary theory describe evolutionary individuals even more directly, as the things whose fitness roughly corresponds to their expected number of babies. If we define the evolutionary individual incorrectly—or rather, if we identify a real case as an evolutionary individual incorrectly—then we stand in danger of making false fitness measurements and, ultimately, getting the evolutionary book keeping wrong (Clarke 2012; In review).
The aim of this paper is to construct a measure of evolutionary individuality by focusing on the way natural selection acts at different hierarchical levels. Section two introduces the idea that Major Transitions can be understood as events during which a crucial underlying variable—the amount of natural selection which acts at one hierarchical level, rather than another—changes. I suggest that we can understand this variable as an empirical correlate of evolutionary individuality, and I present an explicit Pricean measure of the variable as one possible method for measuring evolutionary individuality. In section three I probe what I call ‘levels of selection’ accounts of individuality, to explore some different attitudes which have been adopted in respect of the connection between individuality and levels of selection. I argue that existing approaches can be understood as taking three distinct perspectives, as they focus on either actual selection, or on a history of selection, or on a capacity for selection, at the focal level. In section four I outline Clarke’s definition, which develops the idea that it is the capacity for selection which matters (Clarke 2013). I explain how we can understand what I call ‘individuating mechanisms’ as providing evidence of a unit’s capacity for participating in selection. Finally, section five acknowledges and responds to some objections that may be marshalled at the account I propose: the problem of cross-level by-products and the problem of trait-specificity.
A parameter underlying evolutionary transitions in individuality
‘Evolutionary transitions in individuality’ (ETISs) are evolutionary events during which independently reproducing units come to be mere parts in new higher-level wholes (Michod 1999). There is consensus in the literature that at least four kinds of ETI have taken place: independent genes combined to form chromosomes; independent prokaryotic cells combined to form eukaryotes; independent eukaryotic cells combined to become multicellular organisms; and multicellular organisms combined to form colonial organisms (Okasha 2006; Bourke 2011; West et al. 2015). The Transitions literature assumes that individuality emerges, in the sense that some lineage of individuals comes to acquire the property of evolutionary individuality, over evolutionary time scales, at a new hierarchical level (Clarke 2014). For example, around 800 million years ago all eukaryotes were single celled. Then a lineage of choanoflagellates—unicellular organisms which are morphologically similar to one of the cell types within sponges—began a process of transition.Footnote 1 Now, at the present time, one of the descendent lineages contains horses—unambiguous multicellular organisms. Choanoflagellates divide by fission, have only simple forms of cellular adhesion and a maximum of five different cell types (Fairclough 2015). Horses, by contrast, have eyes, hearts and other specialist organs, bilaterally symmetrical specialised limbs, a centralised nervous system including a brain, and a sophisticated immune system; they reproduce sexually and host myriad symbiotes. Choanoflagellates are standardly considered unicellular, horses are definitely multicellular, yet they are connected by an unbroken chain of intermediate life forms. Clearly a transition occurred somewhere in that chain.Footnote 2 But where? And in virtue of which changes? Which of the many traits that changed along the way are the ones that underwrite a horse’s status as multicellular?
Traits that are often associated with multicellularity in animals include axial symmetry, a separate germ layer, gastrulation and body plans (Ruiz-Trillo and Nedelcu 2015). However, non-animal multicellulars sometimes fail to share these features. Some biologists have tried to identify lineage-general correlates of multicellularity. Genome expansion was initially touted but failed to find empirical confirmation. Complexity of gene regulatory networks (Szathmáry et al. 2001) is beset by the problem that strikes all accounts which depend upon complexity—no one can agree on how to measure it (McShea 2000; Herron and Nedelcu 2015). Likewise, many authors use the number of cell types, but there is a fatal lack of consensus about what should qualify as a cell type (Lang and Rensing 2015). More ambitious still, some authors seek an account of the variable that is general, not only to different lineages, but to different kinds of transition event. In other words, they aim to identify an underlying variable that is common to the emergence of multicellularity, and to the emergence of coloniality, and to all the other consensus ETIs. This rules out the vast majority of candidate traits. Insect colonies didn’t evolve by intercellular adhesion, for example.
The present paper defends a conception of the underlying property in terms of natural selection itself—we say that the new individual appears at a new, higher, level of selection. Before I show how that works, it remains to be asked what value there is in such a general account. What function is served by bundling up the multitude of different correlates into a single general parameter? Why not prefer an account in which all of the correlated details are pulled apart and analysed separately? Surely this will sometimes be the right approach. For example, in respect of the evolution of multicellularity there is value in teasing apart the genetic and phylogenetic stories that explain the independent origins of different mechanisms for gluing cells to one another, and, independently, in disentangling those same stories about the origins of different systems for intercellular communication.
However, there are circumstances in which it is helpful to be able to condense all of the independent parameters within a single metric. To compare which system for gluing cells together brought about the biggest gains in multicellularity, for example. Or to make comparisons of the differential challenges that had to be overcome in transitioning to multicellularity, on the one hand, and eukaryocity, on the other. Whenever we want to do comparative analyses, we need to be able to step back from the causal-mechanical details about actual mechanisms for transition and about lineage-specific adaptations.
Furthermore, a single measure offers to remove some of the ambiguity generated by different authors utilising different parameters and presupposing different thresholds for those parameters. For example, one sometimes has to dig rather hard to discover what parameters underlie statements about which lineages have evolved complex as opposed to simple multicellularity. Some researchers only call a lineage multicellular if it exhibits gene regulatory networks (Valentine and Marshall 2015). Cock and Collén insist that a multicellular individual has to have at least eight different cell types (Cock and Collén 2015). At the other extreme, Solé and Duran-Nebreda accept as multicellular any aggregation that exhibits physical attachment—glues—between cells (Solé and Duran-Nebreda 2015). Not surprisingly, these researchers arrive at very different conclusions about when and how many transitions to multicellularity have taken place—numbers range from 7 to 25 separate events (Ruiz-Trillo and Nedelcu 2015). Use of a universal metric would aid communication in such contexts.
Proposal: a quantitative measure of individuality
Multilevel selectionists assume that the total natural selection acting on a system can be decomposed into distinct partitions which each measure the selection acting at different hierarchical levels (Wilson 1975; Damuth and Heisler 1988; Wilson and Sober 1994; Keller 1999; Goodnight et al. 1992; Okasha 2001, 2006). Sober and Wilson name the two components ‘within-group selection’ (lower-level selection) and between-group selection (higher-level selection) (Sober and Wilson 1998). Put simply, how well some trait does overall is given by the sum of how well it tends to do within groups, and how well the groups it is in tend to do. As an ETI proceeds, we expect the within-group component to diminish, and the between-group component to increase, until there is only the between-group, higher-level component left.
A simple way to capture the extent to which natural selection has shifted up to the higher level, then, is to calculate the relative strength of selection at the higher level, or the proportion of the total selection which acts between-groups, rather than within them.
Definition 1
Higher-level individuality: \({\text{Proportion of selection at the higher level}} = \frac{{{\text{Between}} - {\text{group selection}}}}{{{\text{Within}} - {\text{group selection}} + {\text{Between}} - {\text{group selection}}}}.\)
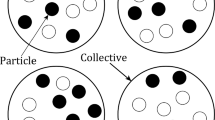
We can imagine a continuum of possible states of a population of particles nested within collectives, from one extreme in which selection occurs exclusively at the lower level, to the opposite extreme in which selection occurs exclusively at the higher level. As we move from one extreme to the other, the proportion of the overall selection that acts at the higher level increases from 0 to 1.
One complication is that selection is directional, and the distinct levels may be under selection in the same direction as one another, or in opposing directions. To accommodate this, we need to use the absolute values of the between-group and within-group terms. In fact, the two selective levels may perfectly cancel each other out, so that there is no overall change in trait frequencies at all. Using the absolute values allows us to retain the idea, in such cases, that there are two levels of selection at work.
So interpreted, we can use this variable to locate living systems on a continuum. We choose a focal unit, and a focal trait, and then peg the units to the left or right according to the proportion of selection at different levels.Footnote 3 Can such a value really be calculated? There is consensus amongst ‘levels of selection’ views that multilevel selection is possible (Damuth and Heisler 1988; Goodnight et al. 1992; Reeve and Keller 1999; Sober and Wilson 1998; Michod 1999; Okasha 2006; Gardner and Grafen 2009; Sober 2011; Gardner 2015). Consensus ceases in regard to the question of how to quantify the action of selection at different hierarchical levels, although many authors agree that the amount of selection occurring at one level rather than another, can be measured. There is, in other words, an objective numerical amount of selection at each level.Footnote 4
One way we might measure this quantity is to replace the terms ‘between-group selection’ and ‘within-group selection’ with the partitions of the multilevel version of Price’s Equation (Price 1970, 1972; Okasha 2006, 2016). The numerator of the ratio in definition 1 would measure the covariance between group fitness and the group character value, while the denominator would sum the latter with the average of the within-group covariances between particle fitnesses and particle trait values. We would need, once again, to use the absolute values of these terms. Assuming that reproduction of particles is clonal, generations are non-overlapping and group values are simply the averages of the particle values, the two terms in the denominator will sum to give the total expected change by natural selection. The whole ratio will give the proportion of the total change that is driven by selection at the higher level.
The multilevel Price equation has been subject to fierce criticism and many people will not accept it as a suitable tool for measuring the amount of selection that occurs at a focal level. Some of these critics will accept an alternative measure and I invite them to substitute such a measure for the Price equation. It will be interesting to investigate what changes such a substitution would precipitate for a levels-of-selection view of evolutionary individuality—whether different verdicts are generated, and so on. Alternatives include a contextualist measure of group versus individual selection (Heisler and Damuth 1987; Goodnight et al. 1992; Goodnight 2013); an analysis of fitness varianceFootnote 5; or a comparison of genetic variance at the different hierarchical levels.Footnote 6 Another alternative would be to use an inclusive fitness framework, and try to quantify the separate direct and indirect components of what Reeve and Keller call ‘the absolute inclusive fitness ‘force’’ (Reeve and Keller 1999, 8). What is essential is that there is some acceptable measure of the extent to which selection acts at one level rather than another—or that decomposes selection into more and less local components, for those who dislike ‘levels’ talk. If there is no such measure then we cannot describe the emergence of evolutionary individuality in terms of an increase in the amount of higher-level selection.
Note that one reason why a group selection measure might fail to work is if the context is one in which there is insufficient group structure. For example, the population may consist of individuals who interact socially with their neighbours, who interact socially with their neighbours, in such a way that there are no discrete interaction groups. In such cases a multilevel selection framework is inapplicable (Godfrey-Smith 2008). Cases like this are often marshalled as a reason to prefer kin selection approaches to group selection approaches. However, in the context of the problem of evolutionary individuality these cases do not undermine group selection approaches—they are simply cases in which evolutionary individuality does not appear at the level of groups.Footnote 7
Another problem with a Pricean measure is that it is usually applied to populations of conspecifics, but the ideal measure will accommodate collectives whose members come from distinct species, as occurs in symbioses. We do not usually consider fitness to be commensurate across diverse species. In these cases we require an alternative measure of the extent to which the members of collectives are competing with one another. One possibility may be to adapt Frank’s measure of ‘codispersal’ which measures the extent to which symbiotic partners are in reproductive synchrony (Frank 1997). ‘Fitness alignment’ similarly measures the extent to which the fitnesses of the partners are correlated (Friesen 2012).
Three alternative levels-of-selection approaches to individuality
I will use the name ‘levels of selection account of individuality’ for any approach which explicates the variable underlying ETIs in terms of the amount of natural selection acting at a focal hierarchical level. Such approaches assume, in other words, that to be an individual, in the evolutionary sense, is to exist at a specific level of a compositional hierarchy—the level at which natural selection acts. We can distinguish three distinct sorts of ‘levels of selection' approach.Footnote 8
Michod’s (1999) account suggests a single parameter underlying a system’s progress through an ETI, in so far as it characterises the parts of a transitioning entity (for example, a volvocine alga’s cells) as experiencing a decline in fitness, so that by the completion of the transition their fitness is zero (Michod 1999, 2006).Footnote 9 Godfrey-Smith uses three dimensions to chart a system’s progress through a process of transition, and explicitly considers this as measuring the system’s degree of evolutionary individuality. He builds upon Lewontin’s definition of a ‘unit of selection’ to locate living systems inside a three-dimensional space, according to their possession of properties that make them more readily evolvable by natural selection. Those with higher values for the properties are deemed to have more individuality, or to be closer to ‘paradigm’ status (Godfrey-Smith 2009). Several authors associate a group’s status as an individual with the amount of within-group conflict (Dawkins 1982; Reeve and Hölldobler 2007; Gardner and Grafen 2009). Queller and Strassmann, finally, peg living systems onto a two-dimensional space, in which individuality increases as one variable—cooperation—increases and another—competition—decreases (Queller and Strassmann 2009).
I call all these approaches ‘levels of selection approaches’ because they each assume that the variable which changes as an ETI occurs—the property evolutionary individuality—can be spelt out in terms of a change in the strength of natural selection at some compositional level. However, there are subtly different attitudes that can be adopted in respect of the relation between individuality and selection. One possibility is that we take an object’s individuality to be determined by its actual participation in selection. In other words, we measure the proportion of selection which occurs at the focal level, as above, and take this value as telling us the actual extent to which the objects at that level are evolutionary individuals. A second possibility is that we treat evolutionary individuality as dependent on a history of selection at the focal level. Finally, a third possible approach takes evolutionary individuality as equivalent to a capacity for selection at the focal level in the future. I evaluate each of these perspectives in turn.
Define individuality in terms of actual selection at the focal level
The first, simplest, possibility is to take a living object’s degree of individuality as measured by the extent to which it experiences actual, current selection. Reeve and Holldobler say that their measurement of intergroup conflict “precisely measures a society’s position along a ‘superorganism continuum’” (Reeve and Hölldobler 2007, 9739). We might also understand Lewontin’s position in this way—he argued that an object is a unit of selection if it exhibits heritable variance in fitness (Lewontin 1970). It is implied that if all the individuals in the relevant generation happen, for one reason or another, to have the same number of offspring, then, because there is no selection in that generation, the population contains no units of selection. This is undesirable for two reasons.
Firstly, if an object’s degree of evolutionary individuality is simply identical to its measured value for the proportion of selection at its level, then it will be a property which holds only at the temporal scale of one generation. It will neither obtain at an instant, nor will it be likely to remain constant across different generations.
Secondly, individuality ought to be intrinsic to the unit in question. A definition of the evolutionary individual in terms of the actual proportion of selection at the focal level is weak, because it makes the property hostage to facts which are nothing to do with the unit in question—facts about population size and about the environment, as well as sheer luck.
So while we might take the actual proportion of selection at the focal level as a valuable empirical correlate of individuality, we had better not say it constitutes evolutionary individuality.
Define individuality using evidence of a history of selection at the focal level
The second possibility is to treat individuality as obtaining only when there is evidence of a history of selection at the focal level. Approaches which take this perspective define the evolutionary individual by its possession of features which can be expected only in objects which have historically experienced a particular amount of selection at their level. Many authors focus, for example, on traits that can only be maintained in a system where lower-level conflict is low. Altruistic traits are a prime example because, by definition, they are undermined by lower-level (within-group) selection (Sober and Wilson 1998). If an altruistic trait is present, therefore, it can act as robust evidence that higher-level (between-group) selection has been dominant in the recent history of the system. Likewise, it is often assumed that very complex or delicately integrated traits can only survive if lower-level selection is absent (Williams 1966). Lloyd, for example, argues that genuine individuals can be identified by their possession of adaptations (Lloyd 1995). Adaptations are, by definition, products of selection processes (Sober 1994), so their existence can serve as evidence of a response to a prior selection process. If we can see that an object is adapted, we know that its ancestors have responded to selection.
Another account which might be characterised as taking this approach is Queller and Strassmann’s, though they don’t present it this way. They define evolutionary individuality as obtaining to the extent that an object’s parts cooperate with one another, and aren’t in conflict (Queller and Strassmann 2009; Strassmann and Queller 2010). But if we seek a definition of cooperation, we see that a cooperative trait is standardly defined as a trait which has evolved because it generated a benefit for some beneficiary (West et al. 2015). In other words, two objects are treated as cooperating only if there has been a particular selective history between them.
A definition according to which a unit qualifies as an evolutionary individual only if it has had the right kind of selective history, rather than in virtue of what it happens to be doing right now, avoids the problem associated with organisms which, for extraneous reasons, are not currently undergoing selection. It also yields a property which applies at all instants of an organism’s lifetime. A historical definition, furthermore, is able to accommodate many properties which are popularly associated with organismality—for example, organisational complexity, functional integration, division of labour—in so far as complex adaptations are prioritised as evidence for historic higher-level selection.
However, while a historical definition of the evolutionary individual will be useful in contexts in which we seek to give an explanation of selective dynamics that have already happened, biologists sometimes need a concept which can do more. Evolutionary modellers generally want a concept which can support generalisations and predictions about the future. For these purposes a historical, backwards-looking definition is of limited use. Just as a trait’s status as an adaptation is separable from its status as adaptive (Sober 1994), so a unit may have been selected in the past, without continuing to be selectable in its own right in the future. A backwards-looking definition is descriptive, but not modal—it cannot support counterfactual inferences. Can we find, instead, a forwards-looking definition of an evolutionary individual?
Define individuality using evidence of a capacity for selection at the focal level
A last possibility is to make future participation in selection essential to being an evolutionary individual. A forwards-looking definition of the evolutionary individual considers a unit to qualify in virtue of facts about how the unit will respond to selection in the future.Footnote 10 Such a concept can be used by an evolutionary biologist in making predictions about future selective dynamics. But how can we accommodate facts that are essentially inaccessible to empirical reach? How can we arbitrate individuality on the basis of things that might happen: on the future?
Gardner and Grafen state that it is not actual selection which determines a unit’s status as an organism, but potential selection. Their concept is thus intended to have modal force. But Gardner and Grafen fall short of securing a forward-looking concept of the organism. They try to secure the impossibility of future selective conflict by making a stipulation about how much genetic variance exists at the focal level—they assume that if a group is clonal, then there is no potential for within—group selection (Gardner and Grafen 2009). This is alongside an assumption that the presence of policing mechanisms can suffice to eliminate the possibility of within-group selection, with which I concur. But there are two problems with the assumption about clonality. One is that genes are not the only source of heritable variance in fitness in nature. Epigenetic differences, or possession of different symbionts, can be heritable and can affect fitness, for example. The other is that a unit’s status as clonal is a fact about its current, actual state—not about its potential or possible states. And given everything we know about rates of mutation and gene transfer, it is not a state that we can reasonably expect any macroscopic group to remain in for very long. Clonality is a transient and fragile state, not the sort of property which will support inferences about the future.
In order to secure a definition of the evolutionary individual that has modal force, we need to make a specification about what is and is not possible for evolutionary individuals, and we can do this by referencing mechanisms which rule some possibilities out. Many such mechanisms are identified in the literature on evolutionary individuality. For example, developmental bottlenecks are thought to be important because they reduce the potential for lower-level selection by sieving out genetic variation (Dawkins 1982; Maynard Smith and Szathmary 1997; Godfrey-Smith 2009). Sexual reproduction, on the other hand, makes higher-level selection more powerful, by generating genetic novelties (Janzen 1977; Harper 1977). Egg-eating behaviours in worker social insects mean that even if workers would like to compete against their sister workers by raising offspring of their own, it is not possible (Ratnieks and Visscher 1989).
Clarke names as ‘Individuating mechanisms’Footnote 11 any such properties or mechanisms that have the effect of determining a collection of objects’ capacity to evolve by natural selection (Clarke 2013 Footnote 12). Different lineages use different mechanisms, but they all function by influencing the extent to which objects are able to exhibit heritable variance in fitness. Other examples of ‘individuating mechanisms’ include germ separation, immune regulation and physical boundaries. Individuating mechanisms can achieve their effect by affecting genetic variance, by affecting the extent to which genetic variation is heritable, by affecting the extent to which genetic variation has fitness effects, or by affecting other, non-genetic, sources of heritable variance in fitness. For example, transposon silencing mechanisms (siRNAs) prevent conflict by eliminating the fitness effects of genetic variants, and so achieve a suppression of the evolutionary individuality of the transposons (Ågren 2014).
Individuating mechanisms act together to determine the potential of any object to participate in evolution by natural selection. They fix the extent to which any lineage may act as a unit of selection—not just now, but in the immediate future. Those accounts of evolutionary individuality which make the possession of individuating mechanisms essential therefore achieve a definition with modal force.
Godfrey-Smith offers a definition which enjoys modal force—he says a population contains evolutionary individuals in so far as it has the capacity to evolve by natural selection. He develops Lewontin’s conditions to describe what fixes the relevant capacity. In the context of ‘collective’ individuals, Godfrey-Smith makes the possession of particular properties necessary—two policing mechanisms plus a third criterion ‘integration’. Clarke (2013) expands the list of sufficient individuating mechanisms by defining them functionally, so that the possible realisers of the individuating role are unlimited.
Clarke’s definition of the evolutionary individual achieves its forwards-looking, modal, status by making the possession of individuating mechanisms essential to being an evolutionary individual.
Definition 2
An evolutionary individual = a collection of living parts which has some capacity for responding to selection at the between-collection level, because of the action of individuating mechanisms.
Reference to individuating mechanisms makes the definition more empirically applicable than it would be if it was given purely in terms of a capacity for evolution by natural selection. A capacity is not the sort of property that can be readily identified or measured, unless it is currently realised. But we can use the presence of individuating mechanisms to infer whether or not the objects at a hierarchical level have the capacity, even if the capacity is not currently being realised.Footnote 13 For example, if germ separation is present we can infer that the cells of a system lack heritable variance in fitness—lineages of such cells are not able to evolve independently of the other cells in the system. Individuating mechanisms determine the possibility of a response to selection, regardless of whether any selection is actually occurring.
In section five I describe another advantage of incorporating individuating mechanisms into the definition of the evolutionary individual: it allows us to avoid the so-called ‘problem of cross-level by-products’ (Okasha 2006, 99).
Note that Clarke’s concept is not categorical—different objects will exhibit different degrees of evolutionary individuality, because they will have a greater or lesser capacity. If and when the capacity for natural selection is realised, the proportion of selection at the focal level will be non-zero. Most levels-of-selection theorists have defended one view or another about exactly how much selection ought to act at the focal level before the objects at that level are considered evolutionary individuals. For some the halfway point on the continuum is significant, because only when higher-level selection is dominant over lower-level selection are altruistic traits robust against decay (Dawkins 1982; Sober and Wilson 1998; Bowles et al. 2003; Frank 2012). Others will only consider a unit an evolutionary individual if all or nearly all of the total selection occurs at the focal level (Wilson and Sober 1989; Queller 2000; Hölldobler and Wilson 2009; Gardner and Grafen 2009). This view rules out the possibility of finding systems at intermediate positions on an individuality continuum—only paradigm individuals exist. I side with those authors who prefer a strictly continuous view (Reeve and Hölldobler 2007; Godfrey-Smith 2009; Queller and Strassmann 2009; Clarke 2013) although I can see the value of drawing attention to both the halfway and the maximal threshold in particular contexts.
An important question is how definitions one and two above relate to one another. Definition two tells us if an object is an evolutionary individual—to some degree or other. It doesn’t tell us how far along a transition continuum the object is. We know that it has some capacity to undergo natural selection, but we need to know how big a proportion of the total selective force the object can experience. Measuring the actual selection experienced won’t tell us this—for all the reasons mentioned above. There can be extraneous factors leading the actual amount of selection to differ from what would be expected according to the object’s intrinsic capacity for participation in selection. Nonetheless, the actual proportion of selection acting at a level will often be a useful proxy for an object’s degree of evolutionary individuality, in exactly the same way as actual reproductive output is a useful proxy for fitness. It is far from perfect, because all sorts of real world phenomena can cause an organism to be lucky or unlucky and fail to have the number of offspring we would predict, given its intrinsic properties. But it is the best we’ve got because capacities just aren’t directly measurable. We can strengthen the reliability of the cue by performing multiple measures of the proxy, in different organisms of a type, to converge on a number that gives the degree of individuality that is typical for that type. In the end, it is definition two that has priority. Empirical measures of a correlation between a trait and fitness should only be taken as revealing of an intrinsic capacity for selection—of evolutionary individuality—if there are individuating mechanisms in place. Without this proviso, the measure in definition 1 is liable to incorrectly classify flukes and statistical artefacts as higher-level selection, as I’ll argue in section four.
Unfortunately, we can be led astray if we rely on identifying familiar individuating mechanisms too. A mule is a case of a living object which possesses all of the paradigmatic mechanisms which function to individuate other vertebrates—a developmental bottleneck, germ soma separation, a complex immune system. Mules also seek out and are capable of having sex. But they almost never sire offspring.Footnote 14 Mule individuating mechanisms simply aren’t succeeding to ground a capacity for participation in evolution by natural selection. The capacity itself is necessary to being an individual, and only when grounded in individuating mechanisms is it sufficient.
Two objections
The problem of cross-level by-products
What is the difference between a fleet herd of deer and a herd of fleet deer? It sounds like the opening of a joke, but was intended to motivate a distinction between a group whose members are individually selected, and a target of genuine group selection (Williams 1966). The problem can be seen clearly in a model in which we impose groups by definition.
Assume there is a population of giraffes which exhibit one of two phenotypes: tall or short. Tall giraffes always have a higher fitness than short ones, just because they are able to reach and eat a greater number of acacia leaves. Let us suppose that the giraffes are well mixed; nonetheless we may define two different groups. Group one is composed of all the tall giraffes, while group two is composed of all the short giraffes. Now we can apply the multilevel Price equation to the population of giraffes to find out what degree of individuality is possessed by the giraffes themselves, on the one hand, and the made-up groups, on the other. Disaster strikes: the multilevel covariance analysis yields the answer that the groups exhibit the highest degree of individuality, while the giraffes are mere parts. The reason is that because we arranged the giraffes into groups by height, each group contains zero character variance, so there is no selection within groups. All the variance is between groups—so all the selection is identified as taking place at the level of the groups. According to the Price analysis the giraffe groups are exclusive units of selection: paradigm evolutionary individuals.
But this is highly counterintuitive. Intuitively there is no group selection—the giraffe groups are not individuals. We made them up after all. There are no emergent group properties—group fitness and group phenotype are artificial constructs, just the averages of the giraffe’s fitnesses and phenotypes. The moral of the story is that higher-level covariance does not always indicate higher-level selection: it could be a mere statistical artefact of lower-level selection (Okasha 2006). In fact, higher-level covariance can be generated whenever there is lower-level covariance, by appropriate construction of higher-level groups. All that is required is to guarantee some assortment of types into the higher-level groups (Fletcher and Doebeli 2009).
There is nothing in the Price analysis itself that offers any guidance here (Okasha 2006, 97). Many people have pointed out that the Price approach is only applicable when groups are biologically real, so it is necessary to supplement the equations with some criteria restricting what qualifies as a group.Footnote 15 Sober and Wilson argue that only collections whose members engage in fitness-affecting interactions with one another may be considered suitable targets for Price’s analysis (Wilson 1975; Wilson and Sober 1989; Sober and Wilson 1998; Sober 2011). Giraffes would form a group with respect to height just in case short giraffes have their fitness raised by being in a group with lots of tall giraffes. This could be the case if for example, predators tended to pick groups to attack on the basis of their average height. Then it seems plausible that height really is selected (partly) at the group level. If, on the other hand, short giraffes in tall groups are just as likely to be preyed upon as those in short groups, then the interactionist definition says there are no trait groups with respect to height, and selection acts only on giraffes .
Sober and Wilson’s definition means that groups are trait-specific—a group is the set of particles that interact with respect to a particular trait. For example, to understand selection for altruistic predator warning calls the trait group will be defined by who is within ear shot, but for resource use it depends instead on who competes for resources. Some people have found the trait-specificity of the groups defined in this way odd, because groups defined for different traits need not coincide with one another. Another worry is that there is a sort of Sorites problem. Interaction is a continuous term, but how much is necessary? Doesn’t everything in the universe interact with everything else, in some sense?
Finally, and perhaps most seriously, the trait-group definition makes some inappropriate inclusions, because it doesn’t specify that fitness-affecting interactions must be group-structured. Suppose that short giraffes are less likely to be attacked by a predator while they are standing close to a tall giraffe. Then there is a fitness-effect for height. But giraffes wander about in such a fashion that tall B stands next to short C 1 day, while the next day C stands close to tall D while B huddles close to short A. Their interactions are neighbour-structured, but not group-structured, because the interaction is not transitive (Godfrey-Smith 2008; Birch forthcoming). Sober and Wilson’s definition would imply that a distinct group exists for every single giraffe plus its own interaction partners, but while such groups overlap, they fail to coincide. A group may therefore meet Sober and Wilson’s criteria for group-hood, even though a multilevel selection analysis is inappropriate, and a kin selection analysis would better capture the dynamics of the relevant social interaction. The trait-group definition therefore fails to identify ‘biologically real’ groups in this case, and still leads to a situation where Price’s analysis will generate the intuitively wrong answer about whether or not group selection is at work. Restricting the application of Price’s analysis to cases where the relevant trait is ‘social’ meets the same problem (Okasha 2016).
I propose a modification of Sober and Wilson’s definition that avoids these problems. We simply define groups by their possession of individuating mechanisms. A policing mechanism, by definition, inhibits the expression of heritable variance in fitness amongst the members of a collection. So fitness-affecting interactions may qualify as policing mechanisms, in so far as they tie the fitnesses of members of a group together. However, while fitness-affecting interaction achieves this affect by direct causation—one member causally affects the fitness of the other member—a policing mechanism can achieve the same effect without any direct causation between the two. A policing mechanism can act as a common cause on the fitness of both members. And in the end, it doesn’t matter, as far as future selective dynamics are concerned, why the fitness of two units is correlated, only that the correlation is not a temporary fluke.
Furthermore, policing mechanisms can be defined as tying the fitnesses of all of the members of a group, so that piecemeal, neighbour-structured interactions do not qualify. Germ soma separation, for example, is a policing mechanism whose action ranges over all the germ and soma cells in an organism, regardless of the extent to which cells engage in direct interactions with one another. Similarly, a worker bee doesn’t need to actually eat the eggs of a fellow in order to constitute a part in a higher-level individual along with it. It is enough that egg-eating takes place, so that if any worker in the colony lays an egg then it will be eaten, by someone.
The giraffe herds qualify as individuals, on this view, only if there are mechanisms enforcing the between-group variance and the within-group homogeneity for height. What sort of mechanism would fit the bill here? There would need to be something which forced the tall giraffes of a group to remain in sufficient proximity to the short members that the fitness of all the giraffes is affected by the group’s average height. Some mechanism of adhesion would do the job, perhaps a hormonal driver of behaviour. The adhesion mechanism would thereby force the members of a giraffe group to interact with one another, in respect of the predator-mediated fitness-effect of height. The group would be delimited by the hormones. Tall giraffes wouldn’t be able to escape the fitness-drag of being stuck with short group-mates. Under such conditions, I see no objection to interpreting the group-level covariance between height and fitness as higher-level selection—in viewing the giraffes, in other words, as subject to group selection.
We escape the problem of cross-level by-products, in Price’s analysis, by dictating that group-level covariance between traits and fitness can only be interpreted as higher-level selection when that covariance is maintained by individuating mechanisms. We still keep the advantage that “the groups are decided by the biology of the organism, not the whim of the biologist.” (Wilson 2010, 16) (17).
Is individuality trait-specific?
One problem we cannot completely escape is the implied trait-specificity of evolutionary individuality. Price analysis picks out levels of selection with respect to specific phenotypic traits, but there is something odd about a trait-relative concept of the individual. As Wilson puts it, “the concept conflicts with the image of an organism as a unit that is adaptive with respect to many traits. After all, an individual organism like a bird eats as a unit, flies as a unit, fights as a unit, and so on.” However, another reason to make individuating mechanisms essential to individuality is that they go some way to ameliorating this problem. A mechanism which prevents fitness differences between the parts of an object in respect of one trait will often prevent differences in respect of other traits at the same time. Giraffes that huddle together will affect each other in many ways that aren’t to do with height. Germ separation inhibits all fitness differences between cells, without differentiating separate causes.
However, the group delimited by one individuating mechanism may not coincide with the group delimited by a different individuating mechanism. For example, the vertebrate immune system may facilitate group-structured cooperative interactions between humans and their gut bacteria. If those bacteria are passed horizontally from a parent, then the human+bacteria unit may act as an evolutionary individual, in respect of certain traits. For example, there is some evidence that bacteria acquired from the mother during birth are not only accepted by the immune system, but are important for the immune system’s optimal development (Macpherson and Harris 2004).The mechanism of the developmental bottleneck excludes those same bacteria, however, because they didn’t develop from the germ cell. I would favour a permissive view here, so that any object qualifies as a part of an evolutionary individual if at least one mechanism is successful in ensuring that the part has some capacity to be selected along with the rest.
Conclusions
This paper develops the idea that we can understand the parameter underlying evolutionary transitions in individuality in terms of natural selection. I propose that we treat the ratio of between-group selection to the sum of between-group selection and within-group selection as a measurable empirical correlate of the degree of evolutionary individuality possessed by groups.
I explained why we shouldn’t consider the ratio as defining a collective’s degree of individuality more directly, and why, furthermore, we should incorporate the possession of ‘Individuating mechanisms’ into the definition in order to achieve a concept of the evolutionary individuality which supports predictions and other modal inferences about evolutionary dynamics.
The resulting levels-of-selection account defines a living object’s degree of evolutionary individuality (the property that moves to a higher-level as an ETI proceeds) in terms of the capacity/potential of the compositional units at the different hierarchical levels to undergo evolution by natural selection.
The account provides a species-neutral, transition-neutral, quantitative measure of evolutionary individuality which can be used in making comparisons across species and across levels. Unlike other levels-of-selection accounts it secures a forwards-looking, modal concept, but without sacrificing generality. By referencing individuating mechanisms the definition also avoids problems of trait-specificity and of cross-level by-products.
Notes
The phylogeny of multicellularity is very hard to unpick, but a popular theory is that metazoans evolved by heterochrony from an ancestor that was closely related to a sponge, descended from a choanoflagellate, around 780 million years ago (Valentine and Marshall 2015).
Note that transitions are not inevitable or unidirectional. For example, various fungal lineages are thought to have gained multicellularity and then later transitioned back to unicellularity (Sharpe et al. 2015, 9).
I will argue in section five that, thanks to the action of individuating mechanisms, much of the time we will get the same result regardless of which trait we choose.
Even those authors who dissent will concur that there is a fact of the matter about which of two hierarchical levels is dominant, in any case (Sober 2011).
ANOVA of fitness would fail in respect of cases, such as germ separated cases, in which some parts of the individual exhibit much higher fitness than others.
Another alternative would be to simply compare the levels of genetic variance at the different hierarchical levels. However, genetic variance is neither necessary nor sufficient for evolution by natural selection. It is not necessary because there can be non-genetic sources of heritable variance in fitness, such as differential vertically transmitted symbionts. It is not sufficient because genetic variants can be prevented from passing their traits onto offspring, as in the case of sterile worker insects.
Although, as Birch points out, the extent to which a population is group-structured versus network-structured may itself be continuous, so that groups may have an intermediate level of groupishness (Birch Forthcoming).
This characterisation of evolutionary individuals is far from universal. For example, when Hull discusses the individuality of species he is concerned with their particularity, rather than with whether selection acts at the level of species (Hull 1978).
It is unlikely, however that this variable can be empirically measured. Shelton and Michod introduce a notion of ‘counterfactual fitness’ in which we try to make informed judgments about how a unit would fare if it was removed from its social setting (Shelton and Michod 2014).
More precisely, facts about how a lineage of the unit in question will respond to selection in the future.
Which include but are not limited to ‘policing mechanisms’ (Reeve and Keller 1999), and ‘conflict modifiers’ (Michod and Roze 2001).’ Individuating mechanism’ forms a broader class, because it includes what I call ‘demarcation mechanisms’, which enhance focal-level selection, in addition to policing mechanisms, which suppress lower-level selection.
The full definition which is defended in Clarke (2013) says that an individuating mechanism is a mechanism that either limits an object’s capacity to undergo within-object selection, by decreasing the availability of within-object heritable variance in fitness (Policing kind), or increases its capacity to participate in a between-object selection process, by increasing the availability of object-level heritable variance in fitness (Demarcation kind).
To avoid circularity, we will need to appeal to cases in which there is actual selection at the focal level to justify consideration of a particular mechanism as an individuating mechanisms—as grounding the capacity, in other words.
Rare exceptions have been known (Rong et al 1988).
Another is to abandon the Price analysis in favour of the contextual approach. This technique of regression analysis avoids the problem of cross level by products, but it has problems of its own. In particular, it yields the counterintuitive result that group selection can occur even in the absence of variation between groups (Okasha 2006).
References
Ågren JA (2014) Evolutionary transitions in individuality: insights from transposable elements. Trends Ecol Evol 29(2):90–96
Birch J (Forthcoming) The philosophy of social evolution
Bonner JT (1974) On development: the biology of form. Harvard Uni Press, Cambridge
Booth A (2014) Populations and individuals in heterokaryotic fungi: a multilevel perspective. Philos Sci 81(4):612–632
Bouchard F (2008) Causal processes, fitness, and the differential persistence of lineages. Philos Sci 75(5):560–570
Bouchard F, Huneman P (2013) From groups to individuals: evolution and emerging individuality. MIT Press, Cambridge
Bourke AF (2011) Principles of social evolution. OUP, Oxford
Bowles S, Fehr E, Gintis H (2003) Strong reciprocity may evolve with or without group selection. Theor Primatol Proj Newslett 1:12
Buss LW (1987) The evolution of individuality. Princeton University Press, Princeton
Clarke E (In review) How to count organisms
Clarke E (2010) The problem of biological individuality. Biol Theory 5(4):312–325
Clarke E (2012) Plant individuality: a solution to the demographer’s dilemma. Biol Philos 27(3):321–361
Clarke E (2013) The multiple realizability of biological individuals. J Philos 110(8):413–435
Clarke E (2014) Origins of evolutionary transitions. J Biosci 39(2):303–317
Cock JM, Collén J (2015). Independent emergence of complex multicellularity in the brown and red algae. In: Evolutionary transitions to multicellular life. Springer Netherlands, pp 335–361
Damuth J, Heisler IL (1988) Alternative formulations of multilevel selection. Biol Philos 3:407–430
Dawkins R (1982) The extended phenotype. Oxford University Press, Oxford
De Sousa R (2005) Biological individuality. Croat J Philos 14:195–218
Dupré J, O’Malley MA (2009) Varieties of living things: life at the intersection of lineage and metabolism. Philos Theory Biol 1:1–25
Ereshefsky M, Pedroso M (2015) Rethinking evolutionary individuality. Proc Natl Acad Sci 112(33):10126–10132
Fairclough SR (2015) Choanoflagellates: perspective on the origin of animal multicellularity. In: Evolutionary transitions to multicellular life. Springer Netherlands, pp 99–116
Fletcher JA, Doebeli M (2009) A simple and general explanation for the evolution of altruism. Proc R Soc Lond B Biol Sci 276(1654):13–19
Folse HJ III, Roughgarden J (2010) ‘What is an individual organism? A multilevel perspective. Q Rev Biol 85:447–472
Frank SA (1997) Models of symbiosis. Am Nat 150(S1):S80–S99
Frank SA (2012) Natural selection. III. Selection versus transmission and the levels of selection. J Evol Biol 25(2):227–243
Friesen ML (2012) Widespread fitness alignment in the legume–rhizobium symbiosis. New Phytol 194(4):1096–1111
Gardner A (2015) The genetical theory of multilevel selection. J Evol Biol 28(2):305–319
Gardner A, Grafen A (2009) Capturing the superorganism: a formal theory of group adaptation. J Evol Biol 22(4):659–671
Ghiselin M (1974) A radical solution to the species problem. Syst Biol 23(4):536–544
Godfrey-Smith P (2008) Varieties of population structure and the levels of selection. Br J Philos Sci 59(1):25–50
Godfrey-Smith P (2009) Darwinian populations and natural selection. OUP, Oxford
Goodnight C (2013) On multilevel selection and kin selection: contextual analysis meets direct fitness. Evolution 67(6):1539–1548
Goodnight CJ, Schwartz JM, Stevens L (1992) Contextual analysis of models of group selection, soft selection, hard selection, and the evolution of altruism. Am Nat 140:743–761
Gould SJ, Lloyd EA (1999) Individuality and adaptation across levels of selection: how shall we name and generalize the unit of Darwinism? Proc Natl Acad Sci 96(21):11904–11909
Guay A, Pradeu T (eds) (2015) Individuals across the sciences. Oxford University Press, Oxford
Haber M (2013) Colonies are individuals: revisiting the superorganism revival. In: From groups to individuals: evolution and emerging individuality, pp 195–217
Harper JL (1977) Plant population biology. Academic, London
Heisler IL, Damuth J (1987) A method for analyzing selection in hierarchically structured populations. Am Nat 130(4):582–602
Herron MD, Nedelcu AM (2015) Volvocine algae: from simple to complex multicellularity. In: Ruiz-Trillo IR, Nedelcu AM (eds) Evolutionary transitions to multicellular life: principle and mechanisms. Springer, New York
Hölldobler B, Wilson EO (2009) The superorganism: the beauty, elegance, and strangeness of insect societies. WW Norton & Company, New York
Hull D (1978) A matter of individuality. Philos Sci 45:335–360
Janzen DH (1977) What are dandelions and aphids? Am Nat 111(979):586–589
Keller L (ed) (1999) Levels of selection in evolution. Princeton University Press, Princeton
Lang D, Rensing SA (2015) The evolution of transcriptional regulation in the Viridiplantae and its correlation with morphological complexity. In: Evolutionary transitions to multicellular life. Springer Netherlands, pp 301–333
Lewontin RC (1970) The units of selection. Annu Rev Ecol Syst 1:1–18
Lloyd E (1995) Units and levels of selection. In: Zalta EN (ed) The Stanford encyclopedia of philosophy (Fall 2005 Edition). http://plato.stanford.edu/archives/fall2005/entries/selection-units/
Macpherson AJ, Harris NL (2004) Interactions between commensal intestinal bacteria and the immune system. Nat Rev Immunol 4:478–485
Margulis L (1970) Origin of eukaryotic cells: evidence and research implications for a theory of the origin and evolution of microbial, plant, and animal cells on the Precambrian earth. Yale University Press, New Haven
Martens J (2010) Organisms in evolution. Hist Philos Life Sci 32(2–3):373–400
Maynard Smith J, Szathmary E (1997) The major transitions in evolution. Oxford University Press, Oxford
McShea DW (2000) Functional complexity in organisms: parts as proxies. Biol Philos 15(5):641–668
Michod RE (1999) Darwinian dynamics. Evolutionary transitions in fitness and individuality. Princeton University Press, Princeton
Michod RE (2006) The group covariance effect and fitness trade-offs during evolutionary transitions in individuality. Proc Natl Acad Sci 103(24):9113–9117
Michod RE, Roze D (2001) Cooperation and conflict in the evolution of multicellularity. Heredity 86(1):1–7
Okasha S (2001) Why won’t the group selection controversy go away? Br J Philos Sci 52(1):25–50
Okasha S (2006) Evolution and the levels of selection. Oxford University Press, Oxford
Okasha S (2016) The relation between kin and multilevel selection: an approach using causal graphs. Br J Philos Sci 67(2):435–470
Pepper JW, Herron MD (2008) Does biology need an organism concept? Biol Rev 83(4):621–627
Pradeu T (2010) What is an organism? An immunological answer. Hist Philos Life Sci 32:247–267
Price GR (1970) Selection and covariance. Nature 227:520–521
Price GR (1972) Extension of covariance selection mathematics. Ann Hum Genet 35(4):485–490
Queller DC (2000) Relatedness and the fraternal major transitions. Philos Trans R Soc Lond B Biol Sci 355(1403):1647–1655
Queller DC, Strassmann JE (2009) Beyond society: the evolution of organismality. Philos Trans R Soc Lond B Biol Sci 364(1533):3143–3155
Ratnieks FL, Visscher PK (1989) Worker policing in the honeybee. Nature 342(6251):796–797
Reeve HK, Hölldobler B (2007) The emergence of a superorganism through intergroup competition. Proc Natl Acad Sci 104(23):9736–9740
Reeve HK, Keller L (1999) Levels of selection: burying the units-of-selection debate and unearthing the crucial new issues. In: Levels of selection in evolution, pp 3–14
Rong R, Chandley AC, Song J, McBeath S, Tan PP, Bai Q, Speed RM (1988) A fertile mule and hinny in China. Cytogenet Genome Res 47(3):134–139
Ruiz-Trillo IR, Nedelcu AM (eds) (2015) Evolutionary transitions to multicellular life: principle and mechanisms. Springer, New York
Santelices B (1999) How many kinds of individual are there? Trends Ecol Evol 14(4):152–155
Sharpe SC, Eme L, Brown MW, Roger AJ (2015) Timing the origins of multicellular eukaryotes through phylogenomics and relaxed molecular clock analyses. In: Evolutionary transitions to multicellular life. Springer Netherlands, pp 3–29
Shelton DE, Michod RE (2014) Group selection and group adaptation during a major evolutionary transition: insights from the evolution of multicellularity in the volvocine algae. Biol Theory 9(4):452–469
Sober E (1994) Conceptual issues in evolutionary biology. Mit Press, Cambridge
Sober E (2011) Realism, conventionalism, and causal decomposition in units of selection: reflections on Samir Okasha’s evolution and the levels of selection. Philos Phenomenol Res 82(1):221–231
Sober E, Wilson DS (1998). Unto others: the evolution and psychology of unselfish behavior. Cembridge (Massachusetts), pp 34–36
Solé RV, Duran-Nebreda S (2015) In silico transitions to multicellularity. In: Evolutionary transitions to multicellular life. Springer Netherlands, pp 245–266
Strassmann JE, Queller DC (2010) The social organism: congresses, parties, and committees. Evolution 64(3):605–616
Szathmáry E, Jordán F, Pál C (2001) Can genes explain biological complexity? Science 292(5520):1315–1316
Valentine JW, Marshall CR (2015) Fossil and transcriptomic perspectives on the origins and success of metazoan multicellularity. In: Evolutionary transitions to multicellular life. Springer Netherlands, pp 31–46
West SA, Fisher RM, Gardner A, Kiers ET (2015) Major evolutionary transitions in individuality. Proc Natl Acad Sci 112(33):10112–10119
Williams GC (1966) Adaptation and natural selection. Princeton University Press, Princeton
Wilson DS (1975) A theory of group selection. Proc Natl Acad Sci 72(1):143–146
Wilson J (1999) Biological individuality: the identity and persistence of living entities. Cambridge University Press, Cambridge
Wilson DS (2010) Darwin’s cathedral: evolution, religion, and the nature of society. University of Chicago Press, Chicago
Wilson RA, Barker M (2013) The biological notion of individual. In: Stanford encyclopedia of philosophy. Stanford University, Stanford
Wilson DS, Sober E (1989) Reviving the superorganism. J Theor Biol 136:337–356
Wilson DS, Sober E (1994) Reintroducing group selection to the human behavioral sciences. Behav Brain Sci 17(04):585–608
Acknowledgments
With many thanks to Samuel Alizon, Pierrick Bourrat, Matthew Herron, Samir Okasha, Thomas Pradeu, Paul Ryan and two anonymous referees.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Clarke, E. A levels-of-selection approach to evolutionary individuality. Biol Philos 31, 893–911 (2016). https://doi.org/10.1007/s10539-016-9540-4
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10539-016-9540-4