
Abstract
Soil organic carbon (SOC) storage is a critical component of the overall sustainability of bioenergy cropping systems. Predicting the influence of cropping systems on SOC under diverse scenarios requires a mechanistic understanding of the underlying processes driving SOC accumulation and loss. We used a density fractionation technique to isolate three SOC fractions that are conceptualized to vary in SOC protection from decomposition. The free light fraction (FLF) is particulate SOC that is present in the inter-aggregate soil matrix, the occluded light fraction (OLF) is contained within aggregates, and the heavy fraction (HF) is associated with minerals. We evaluated surface (0 to 10 cm depth) SOC fraction changes from baseline conditions 5 years after biofuel cropping system establishment at two temperate sites with contrasting soil textures. The biofuel cropping systems included no-till maize, switchgrass, prairie, and hybrid poplar. The FLF concentration (g fraction C g bulk soil−1) did not change significantly from baseline levels under any of the cropping systems at either site after 5 years. Except for poplar, OLF concentrations were reduced in all systems at the site with coarse-textured soils and maintained at the site with fine-textured soils. In poplar systems, OLF concentrations were maintained on coarse-textured soils and increased on fine-textured soils. The HF concentrations also increased under poplar on the coarse-textured soil. A structural equation model indicated that OLF concentrations increased with lower litter C:N, and HF concentrations increased with greater litter quantity and lower litter C:N mass ratios. C:N increased over time within all SOC fractions, suggesting that all pools are sensitive to land-use change on sub-decadal timescales. In agreement with modern SOC theory, our empirical results indicate that increasing litter input quantity and promoting plant species with low C:N litter may improve SOC storage in aggregate and mineral-associated soil fractions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Bioenergy cropping systems are expected to play a significant role in meeting the future demand for sustainable energy (Robertson et al. 2017). In addition to providing a renewable energy feedstock, sustainable bioenergy cropping systems must also provide biogeochemical benefits including ecosystem carbon (C) storage (Robertson et al. 2011). Considering the large historical losses of soil organic carbon (SOC) induced by row crop agriculture (Sanderman et al. 2017), mitigating additional SOC losses and sequestering C in soils is a critical component of the biofuel sustainability equation (Robertston et al. 2008). Thus, a refined knowledge of SOC changes under bioenergy cropping systems is required to facilitate management decisions that optimize the potential for atmospheric CO2 stabilization (Robertson et al. 2017).
Specific vegetation types, management practices, and characteristics of soils may affect the rate or direction of SOC change (Wiesmeier et al. 2019). For example, crop type may influence SOC stocks, as perennial biofuel cropping systems are generally thought to enhance SOC sequestration over annual systems through greater belowground C allocation (Qin et al. 2016). Other management decisions, such as the proportion of stover harvested from maize bioenergy cropping systems, can affect the rate of SOC change (Anderson-Teixeira et al. 2009) with the potential for SOC losses under high stover removal or SOC gains under low stover removal (Qin et al. 2016). Differences in soil texture may also influence SOC stocks through the effects of soil aggregation, with greater aggregate formation and subsequent SOC accrual occurring in finer-textured soils (Baer et al. 2010). While these generalizations are qualitatively useful, a better mechanistic understanding of the underlying SOC processes is necessary to improve quantitative predictions across a wider range of cropping systems, management practices, and soil types (Luo et al. 2017; Abramoff et al. 2018; Blankinship et al. 2018).
The soil density physical fractionation approach (Golchin et al. 1994) provides a theoretical, mechanistic framework for isolating measurable SOC pools that have been shown to be influenced by soil properties, plant inputs, and other land management factors (Fig. 1). In the model framework, plant inputs enter the SOC pool as the free-light fraction (FLF), which is particulate organic matter of identifiable plant origin present in the soil matrix pore space exterior to soil aggregates. Given its location, FLF is not physically protected from microbial contact and hence is available for microbial decomposition. The occluded light fraction (OLF) is particulate SOC that becomes physically encapsulated within aggregates through the processes of comminution, microbial turnover, and aggregation, and thus is partially protected from decomposition (Oades 1988). Disturbance events, such as soil tillage, can physically disrupt soil aggregates, thereby promoting SOC destabilization and subsequent availability to decomposers (Golchin et al. 1994). Finally, the dense or heavy fraction (HF) consists primarily of microbial-processed SOC, which is thought to be more protected from further decomposition through associations with mineral surfaces (Kögel-Knabner et al. 2008; Miltner et al. 2012). Still, the mineral-associated HF is not a permanent C stock, as rhizosphere processes may cause desorption of C and facilitate its degradation (Jilling et al. 2018).

A conceptual diagram illustrating the proposed mechanisms of soil organic carbon (SOC) protection during soil aggregate stabilization and destabilization in relation to soil fractions isolated from a density fractionation approach. The unprotected free light fraction (FLF) is formed when partially decomposed plant materials enter inter-aggregate soil pore spaces. Through the processes of comminution, microbial turnover, and aggregation, the SOC becomes physically contained within aggregates, forming the partially protected occluded light fraction (OLF). The heavy fraction (HF) is formed when SOC becomes adsorbed to mineral particles through various bonding mechanisms. Physical and biochemical processes can lead to SOC destabilization via disaggregation and desorption, respectively
Because each SOC fraction is theoretically distinct, the mechanisms controlling the dynamics of each fraction should vary. For example, the total quantity of SOC stored within the FLF fraction may decline when plant litter inputs are reduced (Lajtha et al. 2014) but may not change as a function of soil texture (Kölbl and Kögel-Knabner 2004). In contrast, the OLF fraction is dependent on texture, as fine-textured soils aggregate more readily than coarse-textured soils (Kölbl and Kögel-Knabner 2004; Baer et al. 2010). Due to the influence of soil texture on mineral surface area, HF storage is also affected by texture, as smaller mineral particles generally have a higher capacity for SOC storage than larger particles (Hassink 1997; Six et al. 2002). Litter quantity and chemistry (e.g., C:N) may also influence HF storage through their effects on microbial growth and carbon use efficiency, with greater mineral-associated SOC storage under high litter input quantity and low litter C:N (Cotrufo et al. 2013; Castellano et al. 2015). Thus, litter quantity, litter chemistry, and soil texture are expected to be key determinants of SOC fraction change, but an integrated understanding of how each fraction responds to all three factors simultaneously is needed to better predict overall SOC responses to land use-change.
Our objective was to determine changes in the quantity and chemistry (C:N) of soil density fractions following 5 years of bioenergy crop production to better understand the mechanisms controlling SOC storage. We hypothesized that the FLF would increase as a function of litter quantity, OLF would increase in soils with the finer texture (i.e., greater clay content), and HF would increase with higher litter quantity and lower litter C:N. Considering the increasingly greater protective capacity and subsequently slower turnover of OLF and HF fractions, we also hypothesized that changes to fraction C:N would be greatest in the FLF and lowest in the HF. We tested our hypotheses by comparing baseline soil fraction measurements at the initial establishment of four bioenergy cropping systems with those taken after 5 years of continuous management at two sites with dissimilar soil texture.
Methods
Sites and cropping systems
Our study sites were the Arlington Agricultural Research Station (ARL) in Wisconsin, USA (43.296° N, 89.380° W) and the Kellogg Biological Station (KBS) in Michigan, USA (42.395° N, 85.373° W). Mean annual temperature and precipitation, respectively, are 6.9 °C and 869 mm at ARL and 10.1 °C and 1005 mm at KBS (NOAA 2017). Soils at ARL are dominated by Plano silt loam, which are Fine-silty, mixed, superactive, mesic Typic Argiudolls (Soil Survey Staff 2017). KBS soils are Kalamazoo loams, classified as Fine-loamy, mixed, active, mesic Typic Hapludalfs (Soil Survey Staff 2017). ARL soils contain an average of 9% sand and 25% clay and KBS soils contain 65% sand and 5% clay, providing a strong textural contrast (Table 1).
The Biofuel Cropping Systems Experiment (BCSE) was established at each site in 2008. The BCSE is a randomized complete block design consisting of five replicates with individual plots measuring approximately 40 × 30 m (Sanford et al. 2016). Prior to BCSE establishment in 2008, both sites had been in agriculture for at least 100 years. At ARL, three of the blocks had been in alfalfa for the 3 years prior to this study and in maize-soy for the 3 years prior to that. The other two blocks at ARL had been in maize for the 4 years prior to the study and alfalfa for the two preceding years. All five KBS blocks had been in alfalfa for the 6 years prior to the study except for two blocks which had been planted to maize in 2006 only. Animal manure spreading was a typical nutrient management practice at ARL and KBS prior to the BCSE.
Our study focused on four bioenergy cropping systems: Continuous no-till maize (Zea mays L.), switchgrass (Panicum virgatum L.), prairie, and hybrid poplar (Populus nigra × P. maximowiczii A. Henry ‘NM6’). The prairie consisted of an 18-species mix of C4 grasses, C3 grasses, non-leguminous forbs, and leguminous forbs. A full agronomic description of these systems and their establishment is given in Sanford et al. (2016). Maize and switchgrass systems annually received an average of 167 and 56 kg N ha−1, respectively. Poplar received a single N application in 2010 at a rate of 210 and 155 kg N ha−1 at ARL and KBS, respectively. Prairie did not receive N fertilizer. Maize received P and K as needed, but perennials did not receive P or K. Maize grain plus about 50% of maize stover were harvested annually following the growing season. Switchgrass and prairie were harvested annually except during the establishment phase, which occurred in 2008 at ARL and 2008 and 2009 at KBS. At ARL, poplar growth was strongly impaired by Marssonina spp. leaf spot fungus beginning in 2010 and extending through 2013. Poplar was coppiced following the 2013 growing season at both sites, at which time standing woody biomass at KBS was more than double the amount measured at ARL (Sanford et al. 2016).
Aboveground net primary productivity (ANPP) was estimated at peak standing biomass in maize, switchgrass, and prairie by clipping all standing biomass at the ground level within three quadrats per plot as described in Sanford et al. (2016). These systems were harvested annually at the plot level, and aboveground soil litter inputs (i.e., unharvested ANPP) were estimated as the difference between ANPP and harvest. For poplar, herbaceous understory ANPP (i.e., primarily weedy biomass) was measured identically to total ANPP in the other systems, but since the understory biomass was not harvested, all understory ANPP was considered as litter input to soils. At KBS, poplar understory ANPP was not collected in 2010 and 2011, so the trend from 2009 to 2012 was linearly interpolated within each plot to estimate understory ANPP in 2010 and 2011. Leaf litterfall ANPP in poplar was measured in two 0.475-m2 (KBS) or 0.375-m2 (ARL) litter traps per plot. Belowground net primary productivity and subsequent belowground litter input were estimated in the perennial cropping systems using six 5-cm diameter, 13-cm deep root ingrowth cores per plot per growing season (Sprunger et al. 2017). In maize, belowground biomass was excavated down to approximately 50 cm following each growing season. We corrected the maize belowground biomass to a 13-cm depth based on observations at ARL that showed that approximately 60% of peak maize belowground biomass occurs within the top 13 cm. Belowground biomass sampling (ingrowth and excavation) was largely incomplete in 2008, so the 2009 values were assumed to be representative of 2008. All biomass was oven dried and a subsample was used for carbon (C) and nitrogen (N) analysis. For belowground biomass (all systems), litterfall (poplar), and understory ANPP (poplar), biomass input C:N was equal to measured C:N mass ratios of the respective biomass. However, to account for plant N resorption during senescence, unharvested biomass C:N was equal to full plant biomass C:N at harvest in switchgrass and prairie and was equal to the C:N of harvested stover in maize (since grain was harvested in maize). An overall C:N mass ratio of all litter inputs was calculated as the ratio of the mass of all carbon in litter inputs to the mass of all nitrogen in litter inputs.
Soil density fractionation
Baseline soil samples were collected in June and November 2008 at KBS and ARL, respectively. Five-year soil samples were taken in November and December 2013 at KBS and in November 2013 at ARL after 5 years of biomass cropping treatments. Soils were sampled in the southern, center, and northern thirds of the plot for a stratified approach. A 7.6 cm diameter hydraulic probe was used to take soil cores to 100 cm, and the cores were sectioned into 0 to 10-, 10 to 25-, 25 to 50-, and 50 to 100-cm segments. Due to the greater variability of deep SOC dynamics at these sites (Szymanski et al. 2019) and subsequent challenge of detecting statistically significant short-term changes, only the 0 to 10-cm section was used in this study. The soil was sieved to 4 mm at KBS and 2 mm at ARL and then dried. For consistency, all soils were re-sieved to 2 mm prior to density fractionation.
A density-based separation procedure (Fig. S1) was used to divide the bulk soil into three fractions (Golchin 1994; Swanston 2005; Marín-Spiotta et al. 2008). Fractionation was performed on two lab replicates per field sample. Oven-dry weight (65 °C) was determined on a subsample for each lab replicate. Approximately 20 g of soil was placed into a 250-mL centrifuge tube, and sodium polytungstate (NaPT) with a specific gravity of 1.6 g cm−3 was added (Cerli et al. 2012). The sample was gently rotated to ensure complete mixing and then placed in a centrifuge for 1 h at 3500 ge (standard earth gravity). The floating FLF was aspirated and the remaining sample was stirred with a benchtop mixer at 1500 RPM for 1 min. The samples were then placed in an ice bath and sonicated using a QSonica Q500 sonicator (QSonica LLC., Newtown, CT, USA) with 475 J mL−1 (Schmidt et al. 1999). We verified the calibration of our sonicator following Schmidt et al. (1999). Following sonication, the samples were centrifuged for 1 h at 3500 ge and then allowed to settle for at least 12 h prior to aspirating the floating OLF.
The FLF and OLF were rinsed over a 0.4-µm polycarbonate filter a minimum of five times with 200 mL each of ultra-high purity deionized water. The HF was rinsed five times by adding 200 mL of ultra-high purity water to the centrifuge tube, vigorously shaking the tube, centrifuging for 1 to 2 h at 3500 ge, and aspirating the liquid from the HF pellet. All fractions were dried at 65 °C. The FLF and OLF were pulverized with stainless steel beads in a high-speed rocking shaker, and the HF was ground to a fine powder using a mortar and pestle. The HF was fumigated with 12 M HCl for 12 h to remove potential carbonates (Harris et al. 2001). Fractions were analyzed by dry combustion for organic C and total N on a Flash EA 1112 elemental analyzer (Thermo Electron Corp., Milan, Italy), with atropine and soils used as standards.
Percent mass recovery of soil fractions averaged 99.7% (Table S1), and there was no statistical difference in percent recovery between sites (p = 0.92) or among treatments (p = 0.42). Therefore, it was not necessary to make corrections for fraction mass recovery.
We expressed the fraction C in two forms: the concentration of fraction C per unit of bulk soil (g C fraction g bulk soil−1) and the proportion of fraction C relative to the total SOC among all three density fractions (g C fraction g C −1DF ). For convenience, we refer to these metrics as fraction concentrations (g C fraction g bulk soil−1) and C proportions (g C fraction g C −1DF ). We distinguish the SOC from the combined density fractions (CDF) from total SOC because the density fractions isolated in this study do not contain the soluble SOC fraction (Crow 2007).
Microbial biomass C
Soil samples for microbial biomass C (MBC) analysis were collected during the late growing season (mid-September) 2014. A previous study indicated similar soil microbial biomass dynamics at both sites (Jesus et al. 2016), and thus we chose to restrict our MBC sampling to ARL. Samples from 0 to 10-cm were collected using a 2-cm diameter probe from six locations per plot along a north–south transect. Samples from each plot were combined, homogenized, and stored at 4 °C until further processing. Microbial biomass assays were performed on two lab replicates per sample following the direct chloroform extraction method (Gregorich et al. 1990). Extracted organic C was determined with the non-purgeable organic carbon method on a Shimadzu TOC-V CSH (Shimadzu Corp., Kyoto, Japan). The microbial extracted organic C was converted to MBC using an assumed extraction efficiency of 0.17 (Gregorich et al. 1990). We chose not to measure microbial biomass nitrogen because soil microbial biomass C:N ratios are generally consistent among ecosystems (Cleveland and Liptzin 2007).
Statistical analyses
Soil fraction and MBC data were checked for equal variance among treatments using Levene’s test. Datasets that failed Levene’s test (p < 0.05) were natural log transformed to reduce heteroscedasticity prior to mixed-model variance analyses. Site-specific mixed models with block as a random effect and cropping system as a fixed effect were run in SAS 9.4 (SAS Institute Inc., Cary, NC, USA). Since we were interested in the changes of the soil fractions from baseline (i.e., from 2008 to 2013), we used contrasts to assess statistical differences between the two sampling periods (at establishment and after 5 years) for each treatment. Resulting p-values were adjusted for multiple comparisons using the step-down Bonferroni method. No baseline measurement was made for MBC, so treatment differences were directly assessed using least-squared mean differences with Bonferroni adjustments for multiple comparisons (p < 0.05).
We used a structural equation model (SEM) to test the hypothesis that changes in the SOC density fractions were related to percent clay (a proxy for mineral surface area), litter quantity inputs to soil (biomass), and litter input C:N mass ratios. Above- and belowground litter properties were combined due to limitations on the number of SEM parameters given our sample size. Within the same SEM, we also tested whether changes among the soil fractions were interdependent. The SEM was coded in the ‘lavaan’ package (Rosseel 2012) using R version 3.4.1 (R Core Team 2017). The resulting SEM model fit was good, with χ2p = 0.69.
Results
Density fractions and MBC
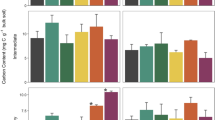
The baseline (2008) FLF concentrations (g C fraction g bulk soil−1) and OLF concentrations did not differ between sites (p > 0.4; Fig. 2a, b). However, the baseline HF concentrations were approximately twice as high at ARL compared to KBS (p < 0.0001). Thus, at ARL a greater proportion (g C fraction g C −1DF ) of baseline C was stored in the HF (p = 0.04) and a smaller proportion of C was stored in the FLF (p = 0.03). On average 93% of baseline C was stored in the HF at ARL while only 86% was stored in the HF at KBS (Fig. 3a, b).

Density fraction bulk soil concentrations at a Arlington (ARL) and b Kellogg Biological Station (KBS). Asterisks indicate significant changes from baseline (2008) samples (p < 0.05 after multiple comparison correction), and error bars are standard errors. Note that the y-axis scales differ between panels and that both y-axes contain breaks

Density fraction proportions at a Arlington (ARL) and b Kellogg Biological Station (KBS). Asterisks identify significant changes from baseline samples (p < 0.05 with correction for multiple comparisons). Baseline samples were taken in 2008, and other treatment samples were taken in 2013
There were no significant changes in the FLF or HF concentrations in any of the cropping systems at ARL, and thus the FLF and HF concentrations were similar among all systems after 5 years (Fig. 2a). However, there was a twofold increase in the OLF concentration in the ARL poplar cropping system (p = 0.005). This change translated to a 117% increase in the OLF proportion and a 5% decrease in the HF proportion (Fig. 3a). The other three ARL cropping systems showed no significant change in OLF concentrations or proportions.
There were no significant changes in FLF concentrations at KBS from baseline (Fig. 2b). However, there were 62 to 72% declines in OLF concentrations in all cropping systems (p < 0.03) except for poplar, and a 23% increase in HF concentrations in poplar (p = 0.04). Declines in the OLF concentrations resulted in 60 to 69% decreases in OLF proportions (p < 0.008), but no significant changes in the FLF or HF proportions in any cropping systems (Fig. 3b).
Baseline fraction C:N decreased in the order FLF > OLF > HF at both sites (Table 2). At ARL, the HF C:N increased significantly in all four systems (p < 0.02) from 2008 to 2013, with the maximum HF C:N change occurring in maize. At KBS, prairie FLF C:N increased from 18.3 to 23.0 (p = 0.0003) and OLF C:N increased from 15.7 to 19.7 (p = 0.0007). When the data were pooled between sites and among cropping systems, the mean FLF C:N increased by 10.4% (p < 0.04), the mean OLF C:N increased by 7.6% (p < 0.02), and the mean HF C:N increased by 5.6% (p < 0.02).
Soil microbial biomass carbon (MBC) at ARL ranged nearly threefold among cropping systems (Fig. 4). Prairie MBC (588 mg C kg soil−1) was greater than maize (214 mg C kg soil−1), and switchgrass and poplar MBC were intermediate to prairie and maize.

Mean (with standard error) microbial biomass C (MBC) at 0 to 10-cm at Arlington, WI during late-summer 2014. Treatments with different letters are statistically different (p < 0.05)
Litter inputs
Unharvested aboveground biomass contributed the most to total biomass inputs to soil in all systems except for poplar, where leaf litterfall was the most abundant input source (Table 3). At both sites, maize had the greatest quantity of unharvested biomass, poplar had the least, and switchgrass and prairie had intermediate amounts. Belowground inputs were smallest in maize and poplar, intermediate in switchgrass, and greatest in prairie. Notably, poplar leaf litterfall was 70% lower at ARL than at KBS. At ARL, total cumulative biomass inputs increased in magnitude as follows: maize > switchgrass > prairie > poplar. At KBS, cumulative biomass inputs increased as follows: poplar > maize > prairie > switchgrass. Overall biomass input C:N mass ratio varied at both sites in the order: maize > switchgrass > prairie > poplar. The overall C:N of maize was more than double that of poplar at both sites.
Structural equation model
Overall, 64% of the change in OLF concentrations was explained by the model (Fig. 5). Clay content was positively related (p < 0.001) and litter C:N was negatively related to changes in the OLF concentration (p < 0.001). However, neither the change in FLF concentrations nor litter quantity were related to changes in the OLF concentrations. Litter quantity was positively related to HF concentration changes (p < 0.05) and litter C:N was marginally negatively related to HF concentration changes (p = 0.063). Changes in the HF concentration were not related to FLF changes, OLF concentration changes, or clay content. There was no effect of litter quantity or litter C:N on changes in FLF concentration.

Fitted structural equation model showing potential relationships between exogenous variables and fraction concentration changes (Δ g C fraction g bulk soil−1). Bolded coefficients indicate statistically significant relationships (p < 0.05) except for the relationship denoted by the asterisk which indicates marginal significance (p = 0.063). Percentages indicate the R2 values for endogenous variables, and the values along each path are the completely standardized coefficients
Discussion
Or results indicate that litter chemistry and quantity (i.e., above- plus belowground) controlled changes in the HF concentration, and litter chemistry regulated changes in the OLF concentration. These findings support the idea that low C:N may be most efficiently converted to stable SOC, as outlined by the Microbial Efficiency-Matrix Stabilization framework and subsequent models (Cotrufo et al. 2013; Castellano et al. 2015). In accord with contemporary SOC stabilization theory, our results suggest that greater overall litter input quantity (i.e., above- plus belowground) has a positive effect on SOC stabilized in the mineral-associated SOM pool. At the same time, we found that C:N changed within all fractions, suggesting that even the mineral-associated pool is dynamic over relatively short timescales (e.g., Torn et al. 2013; Jilling et al. 2018). Overall, our results provide strong empirical support for modern SOC theories and indicate that the soil density fractionation scheme isolates mechanistically relevant pools for developing and calibrating new SOC models (e.g., Abramoff et al. 2018).
Litter quantity and chemistry affected the protected fractions
Changes in OLF concentrations were negatively related to litter C:N, indicating that litter with high N concentration enhanced the OLF. One possible explanation is that the lower litter C:N ratio promoted greater microbial carbon use efficiency (CUE) of the litter or shifted the community towards microbes with higher intrinsic CUE, which increased microbial biomass (Manzoni et al. 2012) and subsequently increased total C inputs into the OLF. Our microbial biomass findings provide some support for this explanation, as perennial systems trended toward greater MBC than annual systems. In agreement, Jesus et al. (2016) found greater total bacterial biomass, total fungal biomass, and arbuscular mycorrhizal fungi biomass in perennial systems compared to maize in soil samples collected from the same two sites as our study. Using laboratory incubations, Szymanski et al. (2019) found greater cumulative microbial CO2 respiration in perennial compared to annual cropping systems at the same sites, indicating higher microbial activity in perennial systems. Considering that microbial residues contribute significantly to SOC pools (Miltner et al. 2012; Kallenbach et al. 2016), we would expect that cropping systems with higher microbial biomass and activity would correspond to greater microbial residue inputs into the protected soil fractions.
The effects of litter quantity and litter C:N on HF concentrations may reflect dynamics of other fractions, especially the FLF. For example, if large quantities of low C:N litter enter the FLF, they are likely to be efficiently converted into microbial biomass and decomposition byproducts (Cotrufo et al. 2013; Vogel et al. 2015), which then may enter the HF directly (Cyle et al. 2016) or through the OLF. Alternatively, if high C:N litter enters the FLF, then low microbial carbon use efficiency may reduce the amount of C available for transfer to other fractions. These dynamics might also be contingent upon the relative C saturation state of mineral surfaces of the HF (i.e., the remaining physical capacity of minerals to adsorb SOC), with lower litter-to-mineral associated SOC conversion rates occurring near saturation (Castellano et al. 2015). While the HF is unlikely to be saturated at either of our long-term agricultural sites (Six et al. 2002), a significant increase in HF concentration was found only in the poplar system at KBS, suggesting that the KBS HF pool may be further from saturation than the ARL HF pool. However, it is also important to note that the five-year cumulative poplar biomass inputs at KBS were twice that of poplar at ARL, which makes direct comparisons difficult.
Despite the reported sensitivity of FLF to land management practices (Wander and Yang 2000; Sequeira and Alley 2011), we did not detect any change in FLF concentrations across a twofold range of litter inputs. In a 20-year litter manipulation study, Lajtha et al. (2014) reported no response of FLF to doubled litter inputs, whereas decreased litter inputs caused declines in the FLF. The authors attributed the consistent FLF under doubled litter inputs to greater microbial activity and respiration (Lajtha et al. 2014). We expect that in systems with high plant litter inputs, most of the litter initially entering the FLF would have been rapidly mineralized and only a small portion transferred to another SOC fraction. It is important to note that both sites in our study had been planted mainly to alfalfa, a perennial legume, for several years prior to the establishment of biofuel cropping systems. Thus, our baseline FLF concentrations were likely somewhat greater than what we would have expected if the previous land use was dominated by typical annual, tilled row crop agriculture (e.g., Jia et al. 2006). Therefore, the high baseline FLF concentration prior to conversion may have limited the potential for further increases under bioenergy cropping systems.
The lack of significant relationships in concentration changes among the three SOC fractions indicates that the fractions can change independently. Thus, while the SOC fractions cannot be considered completely homogenous (Wagai et al. 2009; Schrumpf and Kaiser 2015), our results support the use of these fractions as measurable pools for SOC modelling (Sohi et al. 2001). However, our approach did not account for the spatial and temporal dynamics of litter inputs. Notably, our SEM model contained only one compartment of litter input quantity and chemistry, but the partitioning of above- versus belowground litter may differentially affect SOC fractions (Austin et al. 2017; Ghafoor et al. 2017) and thus may contribute to some of the unexplained variability in our model fit.
C:N changes were evident within all fractions
Across all cropping systems, all fractions showed trends of increasing C:N ratios after 5 years, with greatest fraction-specific gains in C:N in the FLF, intermediate in the OLF, and smallest in the HF. Considering that the fraction C turnover time tends to decrease in the order HF > OLF > FLF (Schrumpf and Kaiser 2015), our finding likely reflects the longer turnover and subsequent lower incorporation of new C inputs into lower order fractions. Given that the HF is often considered a relatively stable SOC pool, with typical mean residence times on the order of hundreds of years (Crow et al. 2007; Schrumpf and Kaiser 2015), it is somewhat surprising that significant changes in the HF C:N ratio were apparent after only 5 years. This could have resulted either from microbial utilization or “mining” of the HF N or from the replacement of older, low C:N stocks with newer, high C:N material. In support of our observation, Szymanski et al. (2019) reported that the mean age of microbial CO2-C respired from established maize and switchgrass systems at ARL was 529 and 267 years, respectively, indicating that older SOC is an appreciable source of total microbial respiration. Overall, the rapid changes in HF C:N in our study support the idea that the mineral-associated pool is somewhat sensitive to land management practices (Grandy and Robertson 2007), that it may be influenced by rhizosphere processes (Jilling et al. 2018), and that some portion may turn over on short timescales (Torn et al. 2013).
The low baseline C:N among the fractions was likely a result of the low C:N alfalfa biomass and manure inputs that had occurred for several years before the biofuel crops were established. For example, aboveground alfalfa biomass grown in 2016 at ARL had a C:N ratio of 15:1 (unpublished data). While the four biofuel cropping systems varied widely in the C:N of biomass inputs, in all systems the biomass input C:N was much greater than the C:N of historical alfalfa biomass inputs. Fornara et al. (2011) found a positive correlation between plant C:N and FLF C:N in temperate grasslands, suggesting a link between litter input C:N and soil fraction C:N. Thus, the trends toward higher fraction-specific C:N in our study partially reflect the increased C:N of litter inputs.
Soil texture influenced aggregate-protected SOC
Consistent with other studies that implicate clay as a key determinant of soil macroaggregation (Kölbl and Kögel-Knabner 2004), clay content was positively related to changes in the OLF C stocks in our study. At least in the short-term, SOC protection in aggregates appears to be much more favorable in fine-textured soils than coarse-textured soils. However, our sites with contrasting soil texture had similar OLF concentrations prior to land use change, indicating that the coarse-textured soils may continue to accrue OLF and eventually reach levels similar to the fine-textured soil. Thus, soil texture may have a greater effect on the rate of aggregate SOC stabilization than on the saturation capacity of the aggregate SOC pool (e.g., Tiemann and Grandy 2015). The lack of a relationship between clay content and HF concentration change does not imply that texture was not a key factor in determining total HF storage. On the contrary, the fine-textured soils in our study stored approximately twice as much C in the HF compared to the coarser textured soils. It should also be noted that soil texture was mostly confounded between our two study sites, so other unaccounted inter-site factors, such as climate-driven differences in soil temperature and moisture, may be implicitly included within the clay variable in our analysis. Nonetheless, our findings are supported by SOC aggregate theories and observations (e.g., Kölbl and Kögel-Knabner 2004), and thus soil texture is likely the dominant driving factor between sites.
Implications for bioenergy production
Our results provided strong evidence that litter input dynamics differentially affected the SOC fractions, and thus specific land management decisions will influence SOC storage potential. Maize, switchgrass, and prairie systems all had similar effects on SOC fractions, but poplar increased aggregate protected OLF on the fine-textured soil and increased mineral associated HF on the coarse-textured soil. Other studies at these sites have also demonstrated divergent SOC properties in poplar at these sites, most notably a larger slow cycling SOC pool compared to the other cropping systems (Sprunger and Robertson 2018; Szymanski et al. 2019). Our results indicated that the contrasting responses of poplar SOC fractions resulted from the large quantity of low C:N poplar litter inputs. Thus, increasing the quantity of litter inputs by harvesting less biomass in the non-poplar systems may increase the C in protected SOC fractions, although this may be contingent on the relative contributions of above- versus belowground biomass inputs. Planting N-rich leguminous cover crops in maize, adding legumes to the switchgrass system (e.g., Jakubowski et al. 2017), or increasing the proportion of legumes in the prairie system may enhance short-term SOC storage (e.g., Fornara and Tilman 2008) so long as the legumes are able to maintain productivity among the other species. Concurrently, increasing the proportion of legumes in prairie or switchgrass systems may come at the cost of lower total litter input quantity, which may subsequently reduce short-term SOC storage (e.g., Lange et al. 2015). Thus, the balance between litter input quantity and litter chemistry (i.e., C:N) is an important consideration for SOC management.
While we did not specifically address the effect of tillage, we found that no-till maize, switchgrass, and prairie all had similar SOC fraction responses within each site. Several studies have reported increases of FLF and OLF in no-till verses tilled annual systems (Wander and Yang 2000; Sequeira et al. 2011) and increases in the particulate organic fraction in untilled perennial systems relative to tilled annual systems (Dou et al. 2013; Kantola et al. 2017), but comparisons between no-till annual and perennial (no-till) systems are scarce. Our results indicated that the no-till practice in maize was as effective as the switchgrass and prairie systems in terms of storing C in the OLF and HF at both sites. Thus, no-till maize may have a similar SOC sequestration potential to common perennial bioenergy cropping systems, at least in the near-surface soil layer, where relative differences in root inputs between crop types are expected to be smallest. However, it is possible that perennial bioenergy cropping systems, which have deeper rooting profiles than annual systems (Black et al. 2017), would have greater long-term SOC sequestration potential deeper within the soil profile compared to annuals.
The SOC legacy of the previous cropping system is also an important consideration for future SOC storage. Although our bioenergy cropping systems were established following long-term agricultural management, our findings may have differed if typical annual row crops, rather than alfalfa, were planted in the years immediately prior to our study. For example, if the FLF and OLF concentrations were somewhat smaller at the beginning of the experiment, as may have been expected in a regularly tilled, continuously annual system, then we may have observed overall increases in those fractions (e.g., Jia et al. 2006; Dou et al. 2013). This implies that the SOC fraction responses to cropping systems observed in our study may not be directly applicable to other locations, and reiterates that the capacity of soils to accumulate SOC is partially dependent on previous land use history (Qin et al. 2016). Experiments with varied SOC fraction baselines will be required to better understand how the soil legacy affects the trajectory of SOC fractions.
Conclusions
Five years after the establishment of bioenergy cropping systems, we found the most prominent SOC changes within the aggregate-protected OLF fraction. The poplar system stood out among the other cropping systems for its capacity to maintain or build OLF. The effect of cropping system on the OLF was driven by the chemistry of litter inputs, with lower C:N litter promoting greater OLF concentrations. Increased HF concentrations were also detected under poplar at the site with coarse-textured soils and was attributable to high litter quantity and low litter C:N. No-till maize, switchgrass, and prairie all had similar influences on SOC fractions, with OLF losses on coarse-textured soils and no change in OLF concentrations on fine-textured soils. All three fractions trended toward higher C:N, indicating short-term sensitivity to land-use change. Our empirical, field-based results largely agreed with contemporary SOC stabilization theories, thus indicating that density-based SOC pools can be incorporated into modern mechanistic SOC models.
References
Abramoff R, Xu X, Hartman M, O’Brien S, Feng W, Davidson E, Finzi A, Moorhead D, Schimel J, Torn M, Mayes MA (2018) The Millennial model: in search of measurable pools and transformations for modeling soil carbon in the new century. Biogeochemistry 137:51–71
Anderson-Teixeira KJ, Davis SC, Masters MD, Delucia EH (2009) Changes in soil organic carbon under biofuel crops. Glob Change Biol Bioenergy 1:75–96
Austin EE, Wickings K, McDaniel MD, Robertson GP, Grandy AS (2017) Cover crop root contributions to soil carbon in a no-till corn bioenergy cropping system. Glob Change Biol Bioenergy 9:1252–1263
Baer SG, Meyer CK, Bach EM, Klopf RP, Six J (2010) Contrasting ecosystem recovery on two soil textures: implications for carbon mitigation and grassland conservation. Ecosphere 1: art5
Black CK, Masters MD, LeBauer DS, Anderson-Teixeira KJ, DeLucia EH (2017) Root volume distribution of maturing perennial grasses revealed by correcting for minirhizotron surface effects. Plant Soil 419:391–404
Blankinship JC, Berhe AA, Crow SE, Druhan JL, Heckman KA, Keiluweit M, Lawrence CR, Marín-Spiotta E, Plante AF, Rasmussen C, Schädel C, Schimel JP, Sierra CA, Thompson A, Wagai R, Wieder WR (2018) Improving understanding of soil organic matter dynamics by triangulating theories, measurements, and models. Biogeochemistry 140:1–13
Castellano MJ, Mueller KE, Olk DC, Sawyer JE, Six J (2015) Integrating plant litter quality, soil organic matter stabilization, and the carbon saturation concept. Glob Change Biol 21:3200–3209
Cerli C, Celi L, Kalbitz K, Guggenberger G, Kaiser K (2012) Separation of light and heavy organic matter fractions in soil - Testing for proper density cut-off and dispersion level. Geoderma 170:403–416
Cleveland CC, Liptzin D (2007) C: N: P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85:235–252
Cotrufo MF, Wallenstein MD, Boot CM, Denef K, Paul E (2013) The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Global Change Biol 19:988–995
Crow SE, Swanston CW, Lajtha K, Brooks JR, Keirstead H (2007) Density fractionation of forest soils: methodological questions and interpretation of incubation results and turnover time in an ecosystem context. Biogeochemistry 85:69–90
Cyle K, Hill N, Young K, Jenkins T, Hancock D, Schroeder P, Thompson A (2016) Substrate quality influences organic matter accumulation in the soil silt and clay fraction. Soil Biol Biochem 103:138–148
Dou FG, Hons FM, Ocumpaugh WR, Read JC, Hussey MA, Muir JP (2013) Soil organic carbon pools under switchgrass grown as a bioenergy crop compared to other conventional crops. Pedosphere 23:409–416
Fornara DA, Tilman D (2008) Plant functional composition influences rates of soil carbon and nitrogen accumulation. J Ecol 96:314–322
Fornara DA, Bardgett R, Steinbeiss S, Zak DR, Gleixner G, Tilman D (2011) Plant effects on soil N mineralization are mediated by the composition of multiple soil organic fractions. Ecol Res 26:201–220
Ghafoor A, Poeplau C, Kätterer T (2017) Fate of straw- and root-derived carbon in a Swedish agricultural soil. Biol Fertil Soils 257
Golchin A, Oades JM, Skjemstad JO, Clarke P (1994) Study of free and occluded particulate organic matter in soils by solid state 13C Cp/MAS NMR spectroscopy and scanning electron microscopy. Aust J Soil Res 32:285–309
Grandy AS, Robertson GP (2007) Land-use intensity effects on soil organic carbon accumulation rates and mechanisms. Ecosystems 10:58–73
Gregorich EG, Wen G, Voroney RP, Kachanoski RG (1990) Calibration of a rapid direct chloroform extraction method for measuring soil microbial biomass C. Soil Biol Biochem 22:1009–1011
Harris D, Horwath WR, van Kessel C (2001) Acid fumigation of soils to remove carbonates prior to total organic carbon or carbon-13 isotopic analysis. Soil Sci Soc Am J 65:1853–1856
Hassink J (1997) The capacity of soils to preserve organic C and N by their association with clay and silt particles. Plant Soil 191:77–87
Jakubowski AR, Casler MD, Jackson RD (2017) Legume addition to perennial warm-season grass swards increases harvested biomass. Crop Sci 57:3343–3351
Jesus EC, Liang C, Quensen JF, Susilawati E, Jackson RD, Balser TC, Tiedje JM (2016) Influence of corn, switchgrass, and prairie cropping systems on soil microbial communities in the upper Midwest of the United States. Glob Change Biol Bioenergy 8:481–494
Jia Y, Li F-M, Wang X-L, Xu J-Z (2006) Dynamics of soil organic carbon and soil fertility affected by alfalfa productivity in a semiarid agro-ecosystem. Biogeochemistry 80:233–243
Jilling A, Keiluweit M, Contosta AR, Frey S, Schimel J, Schnecker J, Smith RG, Tiemann L, Grandy AS (2018) Minerals in the rhizosphere: overlooked mediators of soil nitrogen availability to plants and microbes. Biogeochemistry 139:103–122
Kallenbach CM, Frey SD, Grandy AS (2016) Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat Commun 7:13630
Kantola IB, Masters MD, DeLucia EH (2017) Soil particulate organic matter increases under perennial bioenergy crop agriculture. Soil Biol Biochem 113:184–191
Kögel-Knabner I, Guggenberger G, Kleber M, Kandeler E, Kalbitz K, Scheu S, Eusterhues K, Leinweber P (2008) Organo-mineral associations in temperate soils: Integrating biology, mineralogy, and organic matter chemistry. J Plant Nutr Soil Sci 171:61–82
Kölbl A, Kögel-Knabner I (2004) Content and composition of free and occluded particulate organic matter in a differently textured arable Cambisol as revealed by solid-state (13)C NMR spectroscopy. J Plant Nutr Soil Sci 167:45–53
Lajtha K, Bowden RD, Nadelhoffer K (2014) Litter and root manipulations provide insights into soil organic matter dynamics and stability. Soil Sci Soc Am J 78:S261–S269
Lange M, Eisenhauer N, Sierra CA, Bessler H, Engels C, Griffiths RI, Mellado-Vázquez PG, Malik AA, Roy J, Scheu S, Steinbeiss S, Thomson BC, Trumbore SE, Gleixner G (2015) Plant diversity increases soil microbial activity and soil carbon storage. Nat Commun 6:6707
Luo Z, Baldock J, Wang E (2017) Modelling the dynamic physical protection of soil organic carbon: insights into carbon predictions and explanation of the priming effect. Glob Change Biol 23:5273–5283
Manzoni S, Taylor P, Richter A, Porporato A, Agren GI (2012) Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol 196:79–91
Marín-Spiotta E, Swanston CW, Torn MS, Silver WL, Burton SD (2008) Chemical and mineral control of soil carbon turnover in abandoned tropical pastures. Geoderma 143:49–62
Miltner A, Bombach P, Schmidt-Brücken B, Kästner M (2012) SOM genesis: microbial biomass as a significant source. Biogeochemistry 111:41–55
National Oceanic and Atmospheric Administration (NOAA) (2017) National Centers for Environmental Information: Climate Data Online. www.ncdc.noaa.gov. Accessed 17 September 2017
Oades JM (1988) The retention of organic matter in soils. Biogeochemistry 5:35–70
Qin Z, Dunn JB, Kwon H, Mueller S, Wander MM (2016) Soil carbon sequestration and land use change associated with biofuel production: empirical evidence. Glob Change Biol Bioenergy 8:66–80
R Core Team, 2017. R: A language and environment for statistical computing. https://cran.r-project.org. Accessed 31 October 2017
Robertson GP, Dale VH, Doering OC, Hamburg SP, Melillo JM, Wander MM, Parton WJ, Adler PR, Barney JN, Cruse RM, Duke CS, Fearnside PM, Follett RF, Gibbs HK, Goldemberg J, Mladenoff DJ, Ojima D, Palmer MW, Sharpley A, Wallace L, Weathers KC, Wiens JA, Wilhelm WW (2008) Sustainable biofuels redux. Science 322:49–50
Robertson GP, Hamilton SK, Del Grosso SJ, Parton WJ (2011) The biogeochemistry of bioenergy landscapes: carbon, nitrogen, and water considerations. Ecol Appl 21:1055–1067
Robertson GP, Hamilton SK, Barham BL, Dale BE, Izaurralde RC, Jackson RD, Landis DA, Swinton SM, Thelen KD, Tiedje JM (2017) Cellulosic biofuel contributions to a sustainable energy future: Choices and outcomes. Science 356: eaal2324
Rosseel Y (2012) lavaan: an R package for structural equation modeling. J Stat Softw 48:36
Sanderman J, Hengl T, Fiske GJ (2017) Soil carbon debt of 12,000 years of human land use. P Natl Acad Sci USA 114:9575–9580
Sanford GR, Oates LG, Jasrotia P, Thelen KD, Robertson GP, Jackson RD (2016) Comparative productivity of alternative cellulosic bioenergy cropping systems in the North Central USA. Agr Ecosyst Environ 216:344–355
Schmidt MWI, Rumpel C, Kogel-Knabner I (1999) Evaluation of an ultrasonic dispersion procedure to isolate primary organomineral complexes from soils. Eur J Soil Sci 50:87–94
Schrumpf M, Kaiser K (2015) Large differences in estimates of soil organic carbon turnover in density fractions by using single and repeated radiocarbon inventories. Geoderma 239–240:168–178
Sequeira CH, Alley MM (2011) Soil organic matter fractions as indices of soil quality changes. Soil Sci Soc Am J 75:1766–1773
Sequeira CH, Alley MM, Jones BP (2011) Evaluation of potentially labile soil organic carbon and nitrogen fractionation procedures. Soil Biol Biochem 43:438–444
Six J, Conant RT, Paul EA, Paustian K (2002) Stabilization mechanisms of soil organic matter: implications for C-saturation of soils. Plant Soil 241:155–176
Sohi SP, Mahieu N, Arah JRM, Powlson DS, Madari B, Gaunt JL (2001) A procedure for isolating soil organic matter fractions suitable for modeling. Soil Sci Soc Am J 65:1121–1128
Soil Survey Staff (2017) Natural Resources Conservation Service, U.S. Department of Agriculture. Official Soil Series Descriptions. www.nrcs.usda.gov. Accessed 29 September 2017
Sprunger CD, Robertson GP (2018) Early accumulation of active fraction soil carbon in newly established cellulosic biofuel systems. Geoderma 318:42–51
Sprunger CD, Oates LG, Jackson RD, Robertson GP (2017) Plant community composition influences fine root production and biomass allocation in perennial bioenergy cropping systems of the upper Midwest, USA. Biomass Bioenergy 105:248–258
Swanston CW, Torn MS, Hanson PJ, Southon JR, Garten CT, Hanlon EM, Ganio L (2005) Initial characterization of processes of soil carbon stabilization using forest stand-level radiocarbon enrichment. Geoderma 128:52–62
Szymanski LM, Sanford GR, Heckman KA, Jackson RD, Marín-Spiotta E (2019) Conversion to bioenergy crops alters the amount and age of microbially-respired soil carbon. Soil Biol Biochem 128:35–44
Tiemann LK, Grandy AS (2015) Mechanisms of soil carbon accrual and storage in bioenergy cropping systems. Glob Change Biol Bioenergy 7:161–174
Torn MS, Kleber M, Zavaleta ES, Zhu B, Field CB, Trumbore SE (2013) A dual isotope approach to isolate soil carbon pools of different turnover times. Biogeosciences 10:8067–8081
Vogel C, Heister K, Buegger F, Tanuwidjaja I, Haug S, Schloter M, Kögel-Knabner I (2015) Clay mineral composition modifies decomposition and sequestration of organic carbon and nitrogen in fine soil fractions. Biol Fertil Soils 51:427–442
Wagai R, Mayer LM, Kitayama K (2009) Nature of the “occluded” low-density fraction in soil organic matter studies: a critical review. Soil Sci Plant Nutr 55:13–25
Wander MM, Yang X (2000) Influence of tillage on the dynamics of loose- and occluded-particulate and humified organic matter fractions. Soil Biol Biochem 32:1151–1160
Wiesmeier M, Urbanski L, Hobley E, Lang B, von Lützow M, Marin-Spiotta E, van Wesemael B, Rabot E, Ließs M, Garcia-Franco N, Wollschläger U, Vogel H-J, Kögel-Knabner I (2019) Soil organic carbon storage as a key function of soils—a review of drivers and indicators at various scales. Geoderma 333:149–162
Acknowledgements
Funding was provided by the DOE Great Lakes Bioenergy Research Center (DOE BER Office of Science DE-FC02-07ER64494 and DOE OBP Office of Energy Efficiency and Renewable Energy DE-AC05-76RL01830), the USDA National Institute of Food and Agriculture (Hatch project 0225417-WIS01586), the National Science Foundation (grant DEB-1038759), and the DOE Center for Advanced Bioenergy and Bioproducts Innovation (U.S. Department of Energy, Office of Science, Office of Biological and Environmental Research under Award Number DE-SC0018420). Any opinions, findings, and conclusions or recommendations expressed in this publication are those of the authors and do not necessarily reflect the views of the U.S. Department of Energy. L.G. Oates and G. Sanford provided technical support. Thanks to L. Szymanski and E. Atkinson for demonstrating the density fractionation method and to C. Cavadini, B. Dvorak, C. Rebman, and C. King for lab assistance. Additional thanks to the many individuals involved with the collection and processing of plant and soil samples and to S. Bohm and S. VanderWulp for help with the leaf litter data. We are grateful for feedback provided by M. Rickenbach, M. Ruark, E. Brzostek, and two anonymous reviewers on earlier versions of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Myrna Simpson.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
von Haden, A.C., Kucharik, C.J., Jackson, R.D. et al. Litter quantity, litter chemistry, and soil texture control changes in soil organic carbon fractions under bioenergy cropping systems of the North Central U.S.. Biogeochemistry 143, 313–326 (2019). https://doi.org/10.1007/s10533-019-00564-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-019-00564-7