
Abstract
Plant productivity and tissue chemistry in temperate ecosystems are largely driven by water and nitrogen (N) availability. Although changes in rainfall patterns may influence nutrient limitation, few studies have considered how these two global change factors could interact to influence terrestrial ecosystem productivity and stoichiometry. Here, we examined the influence of experimentally-increased intra-annual rainfall variability and low-level nitrogen addition on aboveground productivity, C and N pools, and C:N ratios in a restored tallgrass prairie across two growing seasons. In the drier first year of the experiment, increased rainfall variability boosted productivity and C pools. In the wetter second year, aboveground productivity and C pools increased with N addition, suggesting a switch in primary resource limitation from water to N. Increased rainfall variability also reduced aboveground N pools in the second year. Community-level C:N increased under increased rainfall variability in the wetter second year and N addition slightly reduced community C:N in both years. These changes in element pools and stoichiometry were mostly a result of increased forb dominance in response to both treatments. Overall, our findings from a restored prairie indicate that increased rainfall variability and N addition can enhance aboveground productivity and C pools, but that N pools may not have a consistent response to either global change factor. Our study also suggests that these effects are dependent on growing season precipitation patterns and are mediated by shifts in plant community composition.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The accrual of carbon (C) in terrestrial ecosystems can mitigate the effect of anthropogenic emissions on climate (Schimel 1995), but may be strongly modulated by continued global change (Reichstein et al. 2013). Multiple lines of evidence suggest that prolonged drought and heavy rainfall events will become more frequent with ongoing climate change, increasing the variability of rainfall and soil moisture in many parts of the world (Alexander et al. 2006; IPCC 2012; Stocker et al. 2014). Similarly, anthropogenic nitrogen (N) deposition has enhanced N availability globally (Galloway et al. 2004). These changes in soil resource availability can strongly influence plant growth, which largely mediates C sequestration and nutrient cycling in terrestrial grasslands (Fay et al. 2002; Knapp et al. 2002).
The effects of rainfall variability on aboveground C and N pools may depend on the system and ambient environmental conditions. In a mesic tallgrass prairie, a 50 % increase in period between rainfall events was shown to reduce plant biomass to the same extent as a 30 % decrease in total rainfall (Fay et al. 2003). There, C4 grasses were strongly inhibited, whereas sub-dominant C3 grasses and forbs were relatively resilient to altered rainfall variability (Fay et al. 2003). Where grassland systems respond similarly to increased rainfall variability as they do to reductions in precipitation, aboveground C and N pools might be suppressed under increased variability as a result of constrained C assimilation and restricted plant N uptake (Knapp et al. 2008). However, not all grasslands respond in this way. Some grasslands have been shown to be particularly resistant to altered precipitation regimes (Kreyling et al. 2008; Jentsch et al. 2011) and plant growth in more xeric systems, where soils are frequently below the wilting point, can even be stimulated by the heavy rainfall events associated with increased variability (Heisler-White et al. 2008; Kulmatiski and Beard 2013). Therefore, the consequences of increased rainfall variability for aboveground element pools are expected to vary between historically dry and wet ecosystems (Knapp et al. 2008) and potentially between wet and dry time periods.
Shifts in plant community composition may also contribute to altered element pools under increased rainfall variability. Compositional shifts toward N-rich forb species with increasing rainfall variability have been observed in at least two grassland systems (Fay et al. 2003; Smith et al. 2016). This effect has been attributed to an enhanced ability of faster-growing forb species to acquire water during heavy rainfall pulses (Smith et al. 2016).
Aboveground C and N pools can also change as a consequence of altered plant plasticity and individual-level stoichiometry. Increased rainfall variability (Fay et al. 2002) and associated dry periods (Jentsch et al. 2011) have been shown to reduce tissue N concentrations and increase C:N ratio in several grasslands by increasing water stress and reducing plant N uptake. While plant N can also increase due to a pulse of plant-available N associated with a re-wetting event, this pulse is not always large enough to offset reductions in plant-available N caused by the preceding dry period (Fierer and Schimel 2002; Borken and Matzner 2009). These changes in plant tissue chemistry suggest that increased rainfall variability can reduce aboveground N pools. However, if increased rainfall variability were to reduce water stress, an increase in tissue N might be expected. Given that few studies have manipulated rainfall variability (Kreyling and Beier 2013), the impact of increased rainfall variability on plant community stoichiometry remains unclear.
Unlike rainfall variability, the response of grasslands to N deposition has been well studied. Nitrogen fertilization typically increases plant-available N, therefore increasing tissue N concentrations and reducing C:N ratios (Ordonez et al. 2009). However, this trend can vary by plant functional type (PFT; Turner and Knapp 1996; Novotny et al. 2007). Increases in plant N concentrations are often associated with enhanced photosynthetic capacity, allowing plants grown under N-enriched conditions to be more productive (Evans 1989; Reich et al. 2001; Borer et al. 2014). Indeed, N deposition has been shown to be a primary driver of grassland productivity, globally (Stevens et al. 2015).
N addition also promotes fast-growing, larger species (Chapin et al. 1986; Vitousek et al. 1997) at the expense of rare species and those adapted to lower N availability (Harpole and Tilman 2006; Dupre et al. 2010; Skogen et al. 2011). Typically, this results in dominant grasses being favored in N-enriched grasslands over sub-dominant forbs, leading to reduced evenness (Foster and Gross 1998; Humbert et al. 2016). However, clonal forb species in particular have been shown to be highly competitive under N fertilization in some grasslands(Fay et al. 2002; Dickson and Gross 2013). Although direct impacts of N addition on plant productivity can dominate ecosystem responses (Manning et al. 2006), these changes in community composition and tissue chemistry also suggest enhanced aboveground C and N pools following N enrichment (Lu et al. 2011a; Yue et al. 2016). Nonetheless, this response may be time-dependent, with stronger effects observed during the peak of the growing season compared to the end of the season, due to plant N resorption at the end of the season.
If increased rainfall variability leads to longer periods of drought stress, increased rainfall variability and N deposition may have counteracting effects on aboveground C and N pools. For instance, N addition has been shown to increase productivity and enhance recovery of grasslands from drought (Kinugasa et al. 2012, but see Bloor and Bardgett 2012), thus allowing for N to partially mitigate impacts of rainfall variability over time. However, many temperate systems are co-limited by water and N (Hooper and Johnson 1999), leading to complex biogeochemical responses to combined shifts in water and N availability (Harpole et al. 2007). Although Leuzinger et al. (2011) identified an antagonistic, non-additive effect of environmental changes on ecosystem processes across many studies, supplemental water and nitrogen have been shown to have synergistic, non-additive effects in some systems (Eskelinen and Harrison 2013). In particular, the pressure on plant communities to shift toward fast-growing species can be synergistically compounded by simultaneous N and rainfall changes (Eskelinen and Harrison 2015; Smith et al. 2016). Thus, strong interactions between increased rainfall variability and N addition are possible, and one might expect these interactions to be antagonistic, synergistic, or additive, depending on the circumstance. Nonetheless, the consequences of these factors for aboveground element pools have not been the focus of many studies.
Here, we evaluate the consequences of increased rainfall variability and N addition for total aboveground biomass as well as C and N pools contained within that biomass in a restored tallgrass prairie. The three measures allowed us to investigate an integrated, broadly comparable metric (aboveground biomass) in addition to two metrics directly influencing biogeochemical cycles (C and N content). We tested three hypotheses: (1) increased rainfall variability decreases aboveground grass biomass more than forb biomass, thus decreasing C and N pools, and increasing C:N ratios, (2) N addition increases aboveground biomass and increases tissue N concentrations, thus increasing C and N pools, and reducing C:N ratios, and (3) in conjunction, these global change factors counteract each other, resulting reduced changes in grass or forb biomass, C pools, N pools, and C:N ratios.
Materials and methods
Experimental site
This research is part of the Prairie Invasion and Climate Experiment (PRICLE), located at the Purdue Wildlife Area near West Lafayette, Indiana, USA (40.4°N, 86.9°W). The mean growing season temperature (May–October; 1901–2011) in the area is 20.3 °C and mean rainfall is 495 mm (Smith et al. 2016). Twelve 4 × 5.5 m plots (each consisting of a 2 × 3.5 m core area surrounded by a buffer 1 m wide) were established in 2012 in a 4-year-old tallgrass prairie restoration strip dominated by Schizachyrium scoparium and Solidago canadensis on a sandy loam soil. The plant community has a mean richness of 10 species per plot, and includes herbaceous species from a wide range of functional types (Smith et al. 2016).
Plots were exposed to a factorial combination of N (ambient or added) and rainfall variability (ambient or increased) treatments over the course of the 2012 and 2013 growing seasons (May–October), within three replicate blocks. Each year, during the first week of June, we added 5 g N m−2 year−1 as slow-release urea fertilizer (ESN, Agrium Advanced Technologies, Loveland, CO, USA) — a rate comparable to those used in other N-enrichment field studies (Reich et al. 2001; Borer et al. 2014). For comparison, ambient wet deposition of N as nitrate and ammonium between 2002 and 2011 was 0.59 g N m−2 year−1 (National Atmospheric Deposition Program Monitoring Location IN41).
Increased rainfall variability was imposed via a combination of partial rainout shelters and hand watering. Shelters were built in an ‘A-frame’ design with a height of 2 m at the center and covering a 4.27 × 5.49 m area (see Smith et al. 2016 for full details). Each rainout shelter used clear, corrugated polycarbonate slats to exclude 50 % of ambient rainfall. Every 30 days, we calculated the total volume of rain excluded from each plot using an on-site rain gauge and sprinkled that volume of water onto plots in the increased rainfall variability treatment. In order to maintain an equal number of rainfall events in each treatment, ambient rainfall variability plots received the equivalent of 1 mm rainfall during each 30-day watering event. Thus, the ambient and increased rainfall variability plots received equal amounts of rainfall in equal numbers of events, but the temporal distribution of rainfall varied between the two treatments. Watering was done using water collected from a nearby well. Ambient variability plots were covered by structures that were identical to those over the increased variability plots, but that did not include polycarbonate slats. Instead, we draped wildlife netting over the ambient variability plot structures, which removed an equivalent proportion of PAR as the polycarbonate slats on average (~5 %) and thus controlled for the shading effects of rainout shelters. We detected no significant differences in temperature between plots covered by rainout shelters or control structures (Smith et al. 2016).
Environmental monitoring
Volumetric water content (VWC; m3 m−3) of the top 10 cm of soil in each plot was measured four times daily throughout each growing season using soil moisture probes (10HS; Decagon Devices, Pullman, WA, USA).
During the second year of the experiment (2013), we used mixed bead ion-exchange resin bags (Ionac NM-60; Sybron Chemicals Inc., Birmingham, NJ, USA) to estimate plant-available NH4 + and NO3 −. Two resin bags (3 g of resin each) were deployed at a depth of 5 cm in each plot concurrently with the June fertilizer application and were retrieved after 77 days in the field. Once collected, each bag was rinsed with deionized water to remove residual soil, roots, and other contaminants. Then, each resin bag was shaken in 90 ml of 2 M KCl over 24 h. Extracts were then analyzed for NH4 + and NO3 − concentrations using a SEAL AQ2 (SEAL Analytical, Mequon, Wisconsin, USA).
Biomass
To examine how increased rainfall variability and N addition influenced plant biomass and tissue chemistry, we harvested biomass from each plot during the middle (July) and at the end (October) of the growing season in 2012 and 2013. In 2012, we harvested all aboveground biomass at a height of 5 cm within two 0.5 × 1 m quadrats in the core area of each plot at each collection date. In 2013, we harvested biomass from the same four quadrats, but sampled half of each quadrat (four 0.5 × 0.5 m quadrats total) during each date in 2013. This was done so that areas sampled in 2012 were evenly distributed between July and October harvests of 2013 and any potential impact of 2012 harvests was distributed evenly across 2013 samples. Thus, while our harvesting technique altered some conditions (e.g. shading and demand for resources), these artifacts were consistent and controlled across plots. Once harvested, biomass was immediately sorted. In 2012, biomass was sorted into two groups: grasses and forbs. In 2013, we sorted biomass by species, and aggregated values based on five plant functional types: C3 grasses, C4 grasses, N-fixers, clonal forbs, and other non-clonal forbs in order to provide more detailed data. Sorted biomass was dried at 65 °C for 48+ h, then weighed, milled, and pulverized. Material from each sorted group was then analyzed for C and N concentrations through combustion analysis (Costech 4010; Costech Analytical Technologies, Valencia, CA).
Calculations and statistical analyses
Soil moisture was analyzed using a repeated measures mixed model analysis of variance (ANOVA). We considered the daily mean VWC in each plot as a function of rainfall variability, N treatment, and the interaction between treatment types. Day was included as a repeated measure.
Plant-available N was analyzed separately for NH4 +, NO3 −, and total plant-available N (NH4 + + NO3 −) concentrations as a function of rainfall variability, N treatment, and the interaction of rainfall variability and N treatment in a hierarchical mixed model ANOVA. Since resin bag data were not pooled by plot, we performed analyses of resin bag data with resin bag nested within plot. Concentrations were log-transformed to meet assumptions of normality.
We calculated C and N pools by multiplying C and N concentration values by biomass for each sorted group (grasses and forbs for 2012 or species for 2013) in each plot during each sampling period. Group-specific biomass and C and N pools were summed within each plot and sampling time. We analyzed these data in three different ways. First, we summed values across all groups for each plot, thus providing community-level biomass, C pools, and N pools. We then analyzed pools and biomass as a function of rainfall variability treatment, N treatment, month (July or October), and year in a repeated measures mixed model ANOVA. Second, we summed these data by type (grass or forb) for each sampling time and analyzed them as a function of type, rainfall variability treatment, N treatment, month, and year in a repeated measures mixed model ANOVA. Lastly, we summed these data by plant functional type (PFT: C3 grasses, C4 grasses, clonal forbs, N-fixing forbs, and all other forbs; see Table S1 for full descriptions) for each harvest of 2013 and examined pools and biomass as a function of PFT, rainfall variability treatment, N treatment, and month in a repeated measures mixed model ANOVA. This analysis was restricted to 2013 data only, as 2012 data were not sorted by species.
To examine the effects of increased rainfall variability and N addition on C:N ratios, we calculated the community-level C:N ratio and the C:N ratio of each PFT. Community-level C:N was defined as the ratio of all sorted groups’ weighted mean C and N concentrations. We then analyzed community C concentration, N concentration, and C:N as a function of rainfall variability, N treatment, month, and year in a repeated measures mixed model ANOVA. Similarly, we calculated the C:N of each PFT as the ratio of its constituent species’ weighted mean C and N concentrations. PFT C:N was analyzed as a function of PFT, rainfall variability, nitrogen treatment, and month in a repeated measures mixed model ANOVA. The PFT-level analysis was restricted to data from 2013 because biomass from 2012 was not sorted by species.
Block and its interactions with rainfall variability and N treatment were included as random factors for all mixed model ANOVAs. All analyses were performed using PROC MIXED in SAS version 9.3.
Results
Soil moisture and N
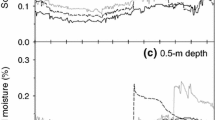
The increased rainfall variability treatment resulted in a 33 % increase in maximum 5-day cumulative rainfall during the 2012 growing season and a 9 % increase in maximum 5-day cumulative rainfall during the 2013 growing season (Schuster 2015). In 2012, the 30-day watering events imposed in the increased rainfall variability treatment provided punctuated relief from a severe drought that took place during the first half of the growing season, as indicated by time spent below the wilting point (Fig. 1; see also Smith et al. 2016). However, these pulses of water came at the expense of lower soil moisture between watering events. In 2013, naturally occurring rainfall was more evenly distributed throughout the season and soil moisture was generally high. During this year, increased rainfall variability almost always acted to increase soil moisture and only reduced soil moisture during the first month of the growing season (Fig. 1). Although the primary effect of increased rainfall variability was to alter the distribution of rainfall, mean volumetric water content (VWC) of the top 10 cm of soil during the growing season was also increased in both 2012 and 2013 (Smith et al. 2016). The magnitude of this difference varied between years (3 % in 2012 and 29 % in 2013), even though the same amount of naturally occurring rain coincidentally fell during the growing season in each year (378 mm).

a Difference in volumetric water content (VWC, m3 m−3) between increased variability (shelter) and ambient variability (control) plots based on data presented in (Smith et al. 2016). Values based on the mean VWC of shelter and control plots at each date. Asterisks indicate peaks associated with watering events conducted as part of the increased rainfall variability treatment. Gray line indicates observed mean VWC for control plots against the right axis. b Mean (±SE) NO3 − concentrations for plots exposed to ambient (left) and increased (right) rainfall variability and ambient N (n, dark) or added N (N, light). Values are least squares means (±SE). Letters signify statistically similar groups (Tukey HSD). Please refer to Smith et al. (2016) for further reference on rainfall and VWC data
The treatments had little effect on the NH4 + and total N captured by the resin bags in 2013 (Table S2). Resin bag extracts also revealed few responses of soil NO3 − concentrations to our treatments, with the exception of a marginally significant interaction of increased rainfall variability and N addition on plant-available NO3 −, where added N increased soil NO3 −, but only under increased rainfall variability (Fig. 1 and Table S2; nitrogen × variability: F1,23 = 3.89, p = 0.06).
Biomass
Community- (Table S3), plant type- (grass or forb; Table S4), and PFT- (Table S5) specific biomass responded strongly to increased rainfall variability and N addition. Community-level biomass was lowest at the mid-season harvest of 2012 when soils were driest, but relatively similar at each of the other three harvests (Fig. S1; month × year: F1,26 = 10.17, p < 0.01). Community-level biomass (Fig. 2) was also enhanced by increased rainfall variability in 2012 (+21 %; variability × year: F1,26 = 4.47, p = 0.04) and tended to increase with N addition in 2013 (N × year: F1,26 = 3.59, p = 0.07). These increases in aboveground biomass were associated with non-additive effects of combined increased rainfall variability and N addition stimulating biomass of forbs by 15 % over fully ambient conditions (N × variability × type: F1,58 = 10.11, p < 0.01). Based on data from 2013, increased forb biomass was largely the result of increased biomass of clonal forbs, which were strongly promoted under the combination of increased rainfall variability and N addition. In contrast, N fixers were synergistically inhibited under the combined treatments (Fig. 3; N × variability × pft: F4,64 = 2.96, p = 0.03).

a Total aboveground biomass of plots exposed to different rainfall variability (ambient or increased). b Total aboveground biomass of plots exposed to different N treatments (center, ambient or added). c Aboveground biomass of grasses and forbs exposed to combined rainfall variability and N treatments harvested in 2012 and 2013. Values are least squares means (±SE). Letters signify statistically similar groups (Tukey HSD). †0.05 < p ≤ 0.10; *p ≤ 0.05

Productivity of C3 grasses (C3G), C4 grasses (C4G), clonal forbs (CF), and N-fixers (NF) under the different rainfall variability (ambient: v, or increased: V) and N treatments (ambient: n, or added: N) in 2013. Values are least squares means (±SE). Letters signify statistically similar groups within each PFT (Tukey HSD). *p ≤ 0.05
Carbon and nitrogen pools
Time was the most consistently influential factor in determining C and N pool size (Table S3). We observed large differences in aboveground C pools and N pools between 2012 and 2013 and with season (+19 %, F1,26 = 9.6, p < 0.01, and +39 %, F1,26 = 20.25, p < 0.01 for C and N respectively). As with biomass, differences in C pools between years were affected by drought at the July 2012 harvest, with a 22 % reduction in pool size compared to the July 2013 harvest (month × year: F1,26 = 10.46, p < 0.01). The end of season harvest showed reductions in aboveground N pools (month: F1,26 = 8.7, p < 0.01), consistent with plant senescence.
Increased rainfall variability increased aboveground C pools in 2012 by 21 %, but did not influence aboveground C pools in 2013 (Fig. 4; variability × year: F1,26 = 4.62, p = 0.04). This resulted in a marginally significant interaction between increased rainfall variability and plant functional type (F4,64 = 2.22, p = 0.08).

Carbon (top) and nitrogen (bottom) pools under the different rainfall variability (left; open: ambient, or hashed: increased) and N treatments (right; horizontal hashing: ambient, or vertical hashing: added) in the 2012 and 2013 growing seasons. Segments in 2012 correspond to contribution to pool by grasses (orange) and forbs (green). Pools are further segmented in 2013 by PFT. Values are least squares means (±SE for complete pool). For complete statistical output and figures of individual segments, see Tables S4–5 and Figs S2–3. †0.05 < p ≤ 0.10; *p ≤ 0.05. (Color figure online)
Nitrogen addition tended to increase aboveground C pools in 2013 (nitrogen × year: F1,26 = 3.26, p = 0.08; Fig. 4). This trend was associated with a significant increase in the contribution of clonal forbs to community C pools under N addition (nitrogen × PFT: F4,64 = 3.71, p < 0.01), which was synergistically enhanced by increased rainfall variability (nitrogen × variability × PFT: F4,64 = 2.91, p = 0.03).
Nitrogen pools were less sensitive to experimental treatments than C pools (Fig. 4). Increased rainfall variability reduced total aboveground N by 19 % in 2013 (variability × year: F1,26 = 8.83, p < 0.01). Increased rainfall variability boosted forb-derived N pools in 2012 (type × variability × year: F1,58 = 5.45, p = 0.02), but this did not lead to increases in the total N pool. Similarly, while N addition tended to stimulate N pools (Fig. 4), the replacement of N-fixers by clonal forbs (nitrogen × pft: F4,64 = 6.11, p < 0.01) resulted in non-significant changes in pool sizes throughout the measurement period (nitrogen × year: F1,2 = 2.91, p = 0.23).
Community and PFT C:N
Increased rainfall variability reduced community tissue N concentration in 2013, but not in 2012 (variability × year: F1,26 = 7.25, p = 0.01).Consequently, increased rainfall variability increased community C:N by 17 % (variability × year: F1,26 = 4.59, p = 0.04) in 2013. N addition never affected community tissue N concentrations or C:N over the experimental period (Table S3). There were no effects of increased rainfall variability or N addition on community C concentration (Table S3).
PFTs within a growth type (grass or forb) responded similarly to N addition and increased rainfall variability. Both C3 and C4 grasses tended to have lower C:N under greater N availability (nitrogen × pft: F4,64 = 2.36, p = 0.06), while N addition did not affect tissue N concentrations or the C:N of forb PFTs (Fig. 5, Table S5). We detected one significant effect of increased rainfall variability on tissue N concentrations (variability × pft × month, Table S5), but this was the result of differences between PFTs over time and not due to differences within PFTs due to rainfall variability (Tukey HSD). Consequently, increased rainfall variability did not alter the C:N of individual PFTs (Fig. 5, Table S5).

C:N ratio of aboveground biomass for C3 grasses (C3G), C4 grasses (C4G), clonal forbs (CF), N-fixing forbs (NF) and all other forbs (OF) based on rainfall variability (left; open: ambient, or hashed: increased) and N treatment (right; dark: ambient, or light: added) for the 2013 growing seasons. Values are least squares means (±SE). Asterisk indicates difference (p ≤ 0.05) between C:N within a PFT
Discussion
Mechanisms of change in biomass, C, and N pools
Biomass and C pools responded strongly to increased rainfall variability—and to a lesser extent, N addition—but the effects of each global change factor were confined to separate years. In the drier, first year of the experiment, rainfall variability had the strongest influence, whereas in the wetter, second year, N addition was the primary driver. This supports the idea that while water can strongly regulate grassland productivity and C pools (Yang et al. 2008), both water and nitrogen can be co-limiting in certain instances (Hooper and Johnson 1999). Furthermore, the positive responses of soil moisture observed here suggest that increased rainfall variability can improve moisture conditions within mesic systems during dry years (Smith et al. 2016). This response is more consistent with those commonly hypothesized for xeric systems (Knapp et al. 2008), and may be due to the non-linear response of soil moisture to rainfall pulses observed during our experimental period (Fig. 1). The response may also have been due to the deep, high field capacity soils at our site, which retained large amounts of water during the rainfall pulses with minimal runoff (Smith et al. 2016). Additionally, biomass and C pools responded similarly over time and to the imposed treatments. This is not surprising given that C typically accounts for close to half of plant dry mass (44 % in this study), and that the C concentrations did not respond to the treatments (discussed below).
Plant-available NO3 − was greatest under increased rainfall variability and N addition, but lowest under increased rainfall variability alone. Although this trend was only marginally significant, it supports that N addition may be able to compensate for reduced mineralization and nitrification (Fierer and Schimel 2002; Borken and Matzner 2009), or increased leaching associated with increased rainfall variability (Schimel et al. 1997; Knapp et al. 2008) to synergistically alter ecosystem function (Rennenberg et al. 2009).
The timing of our treatments’ effects on biomass and C pools implicates community composition as a key regulator of aboveground C pools. Increased rainfall variability and N addition bolstered the dominance of clonal forbs at the expense of C4 grasses and N-fixers, leading to reduced biodiversity and community evenness (Smith et al. 2016). These effects, as well as the changes in plant biomass and C pools we describe here, are consistent with differences in the limiting resources in each year. Increased rainfall variability and associated shifts in the temporal distribution of rainfall and soil moisture stimulated biomass of less drought-resistant clonal forb species over more drought-resistant C4 grasses by aggregating rainfall into larger pulses when water was scarce in the early summer of 2012. Conversely, N addition stimulated C pools when water was more consistently available and N became more limiting (during 2013). These results suggest that these two aspects of global change could increase aboveground biomass and promote aboveground C pools in grasslands, but that increased rainfall variability will influence community composition and productivity the most in relatively dry years and N addition will have the greatest influence in relatively wet years. Thus, the effect of either global change factor is likely partially dependent on inter-annual rainfall variability and associated water availability (Knapp et al. 2008, 2015). Similar compositional effects have been found in response to precipitation variability and fertilization in a mesic, C3 grassland in Europe, but, unlike this study, researchers found a net reductions in biomass in response to the treatments (Grant et al. 2014). The conservative nature of the C4 grasses may have influenced the greater resilience observed here, but more studies or meta-analyses across a diverse range of grassland ecosystems is necessary to fully evaluate this response.
In contrast to C pools, N pools decreased with increased rainfall variability in the wetter, second year of the experiment. Since increased rainfall variability did not detectably affect plant tissue chemistry, reduced N pools were primarily as the result of decreased abundance of N-rich N-fixing forbs in those treatments, indicating that the responses of sub-dominant species can drive ecosystem-level responses. We had expected N addition would have the opposite effect, and that N pools would be greater following N addition. However, we detected a less-reported result of stable N pools under N addition (Lu et al. 2011a). Although N addition increased N concentrations within grasses, the contribution of grasses to N pools did not increase significantly overall (Fig. S3). Instead, our finding of relatively stable aboveground N pools is likely the result of a tradeoff in abundance between clonal forbs and N-fixing forbs, neither of which exhibited altered tissue chemistry in response to N addition. In our system as well as others (Mulder et al. 2002; Oelmann et al. 2011), these functional types have a large contribution to total N pools relative to grasses. Therefore, the responses of clonal forbs and N-fixing forbs mediated community-level N pool dynamics.
Community stoichiometry and stoichiometric flexibility
Changes in the relative abundances of plant functional types largely drove community-level responses of C:N to increased rainfall variability and N addition. The changes we observed in community C:N ratios in response to increased rainfall variability in 2013 can only be ascribed to altered relative abundances of plant functional types since increased rainfall variability did not affect tissue chemistry of individual PFTs. Since increased rainfall variability suppressed lower-C:N N-fixers and promoted higher-C:N clonal forbs (Fig. 5), the increase in community C:N caused by increased rainfall variability was likely the result of a loss of N-fixers (Fig. 3).
The increased biomass of lower-C:N forbs and the reduction of higher-C:N grass species that we observed in response to N addition caused N addition to reduce community C:N. Grasses also maintained higher concentrations of N (lowering their C:N) in response to N addition, which complemented the shift toward lower-C:N forbs. As with previous studies (Turner and Knapp 1996; Manning et al. 2006; Schuster 2015), we found that changes in plant community composition were more important than changes in plant tissue chemistry in determining biogeochemical responses to N addition and increased rainfall variability. Taken as a whole, these findings suggest community composition partially mediates the response of community stoichiometry to global change.
Implications
The increase in aboveground C pools we observed with N addition and increased rainfall variability could indicate a greater capacity for C sequestration under these global changes (Lu et al. 2011b). The stimulatory effects of rainfall variability and N addition on C pools were associated with the increased contribution from clonal forbs, particularly S. canadensis, which has been shown by others to increase soil C pools over a 5-year period (Laungani and Knops 2009). S. canadensis was strongly favored under both increased rainfall variability and N addition in this system, leading to the competitive exclusion of subdominant species and reduced biodiversity (Smith et al. 2016). Reductions in biodiversity like those shown here and in other global change experiments (e.g. Dupre et al. 2010), if persistent over time, can have long-lasting consequences for ecosystem structure and function (Baer et al. 2002) and lead to diminished ecosystem stability (Loreau and de Mazancourt 2013; Hautier et al. 2014). Consequently, the changes in plant community composition reported here may also limit the ability of plant communities to respond to global change (Tilman et al. 2006; Isbell et al. 2009; Hautier et al. 2014). Therefore, while increases in aboveground C pools could translate into increased ecosystem C sequestration, it might do so at the expense of lowered biodiversity.
Care must be taken when interpreting the long-term implications of short-term experiments. Since aboveground C pools in grasslands are not maintained as living biomass, their contribution to ecosystem C sequestration is mediated by decomposition. Leaf litter of forbs decomposes much faster than that of grasses (Prescott 2010; Schuster 2015), and could lead to the increased contribution of C by clonal forbs being offset or overwhelmed by increased decomposition rates and microbial respiration in the long term (Jones and Donnelly 2004; Manning et al. 2006). Furthermore, increased rainfall variability and N addition can increase decomposition rates of forb litter, leading to even greater carbon losses (Schuster 2015). However, short-term studies often considerably overestimate long-term decomposition rates (Harmon et al. 2009), suggesting that C sequestration may be greater than indicated by short-term decomposition trials. Biogeochemical processes may take several years to respond to N or rainfall treatments (Fay et al. 2002; Mueller et al. 2013). Although we were able to detect some strong responses to our treatments, we would likely see more consistent effects over longer time scales, and potentially with greater replication. Thus, while aboveground C pools grew in response to increased rainfall variability and N addition in this study, accompanying changes in decomposition rates leave the longer-term impacts of changes in those pools on ecosystem C sequestration uncertain.
Overall, this study highlights the importance of factors that are extrinsic and intrinsic to the system in determining responses of ecosystem properties to environmental changes. The nature of resource limitation in this system changed from year to year, apparently depending on seasonal precipitation patterns; this extrinsic factor influenced the system’s sensitivity to precipitation variability and N addition. At the same time, element pool responses were strongly responsive to an intrinsic feature of the system: the community’s functional composition and its sensitivity to the treatments. Our results illustrate how projections of ecosystem responses to shifts in short-term climate variability and N deposition must take into account the broader context of the climate and plant community.
References
Alexander LV, Zhang X, Peterson TC et al (2006) Global observed changes in daily climate extremes of temperature and precipitation. J Geophys Res Atmos 111:D05109. doi:10.1029/2005JD006290
Baer SG, Kitchen DJ, Blair JM, Rice CW (2002) Changes in ecosystem structure and function along a chronosequence of restored grasslands. Ecol Appl 12:1688–1701. doi:10.1890/1051-0761(2002)012[1688:CIESAF]2.0.CO;2
Bloor JMG, Bardgett RD (2012) Stability of above-ground and below-ground processes to extreme drought in model grassland ecosystems: interactions with plant species diversity and soil nitrogen availability. Perspect Plant Ecol Evol Syst 14:193–204. doi:10.1016/j.ppees.2011.12.001
Borer ET, Seabloom EW, Gruner DS et al (2014) Herbivores and nutrients control grassland plant diversity via light limitation. Nature 508:517–520. doi:10.1038/nature13144
Borken W, Matzner E (2009) Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Glob Change Biol 15:808–824. doi:10.1111/j.1365-2486.2008.01681.x
Chapin FSI, Vitousek PM, Cleve KV (1986) The Nature of Nutrient Limitation in Plant Communities. Am Nat 127:48–58
Dickson TL, Gross KL (2013) Plant community responses to long-term fertilization: changes in functional group abundance drive changes in species richness. Oecologia 173:1513–1520. doi:10.1007/s00442-013-2722-8
Dupre C, Stevens CJ, Ranke T et al (2010) Changes in species richness and composition in European acidic grasslands over the past 70 years: the contribution of cumulative atmospheric nitrogen deposition. Glob Change Biol 16:344–357. doi:10.1111/j.1365-2486.2009.01982.x
Eskelinen A, Harrison S (2013) Exotic plant invasions under enhanced rainfall are constrained by soil nutrients and competition. Ecology 95:682–692. doi:10.1890/13-0288.1
Eskelinen A, Harrison SP (2015) Resource colimitation governs plant community responses to altered precipitation. Proc Natl Acad Sci 112:13009–13014. doi:10.1073/pnas.1508170112
Evans JR (1989) Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 78:9–19. doi:10.1007/BF00377192
Fay PA, Carlisle JD, Danner BT et al (2002) Altered rainfall patterns, gas exchange, and growth in grasses and forbs. Int J Plant Sci 163:549–557. doi:10.1086/339718
Fay PA, Carlisle JD, Knapp AK et al (2003) Productivity responses to altered rainfall patterns in a C 4-dominated grassland. Oecologia 137:245–251. doi:10.1007/s00442-003-1331-3
Fierer N, Schimel JP (2002) Effects of drying–rewetting frequency on soil carbon and nitrogen transformations. Soil Biol Biochem 34:777–787. doi:10.1016/S0038-0717(02)00007-X
Foster BL, Gross KL (1998) Species Richness in a successional Grassland: effects of Nitrogen Enrichment and Plant Litter. Ecology 79:2593–2602. doi:10.1890/0012-9658(1998)079[2593:SRIASG]2.0.CO;2
Galloway J, Dentener F, Capone D et al (2004) Nitrogen cycles: past, present, and future. Biogeochemistry 70:153–226. doi:10.1007/s10533-004-0370-0
Grant K, Kreyling J, Dienstbach LFH et al (2014) Water stress due to increased intra-annual precipitation variability reduced forage yield but raised forage quality of a temperate grassland. Agric Ecosyst Environ 186:11–22. doi:10.1016/j.agee.2014.01.013
Harmon ME, Silver WL, Fasth B et al (2009) Long-term patterns of mass loss during the decomposition of leaf and fine root litter: an intersite comparison. Glob Change Biol 15:1320–1338. doi:10.1111/j.1365-2486.2008.01837.x
Harpole W, Tilman D (2006) Non-neutral patterns of species abundance in grassland communities. Ecol Lett 9:15–23
Harpole WS, Potts DL, Suding KN (2007) Ecosystem responses to water and nitrogen amendment in a California grassland. Glob Change Biol 13:2341–2348. doi:10.1111/j.1365-2486.2007.01447.x
Hautier Y, Seabloom EW, Borer ET et al (2014) Eutrophication weakens stabilizing effects of diversity in natural grasslands. Nature 508:521–525. doi:10.1038/nature13014
Heisler-White JL, Knapp AK, Kelly EF (2008) Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland. Oecologia 158:129–140. doi:10.1007/s00442-008-1116-9
Hooper DU, Johnson L (1999) Nitrogen limitation in dryland ecosystems: responses to geographical and temporal variation in precipitation. Biogeochemistry 46:247–293. doi:10.1007/BF01007582
Humbert J-Y, Dwyer JM, Andrey A, Arlettaz R (2016) Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: a systematic review. Glob Change Biol 22:110–120. doi:10.1111/gcb.12986
IPCC (2012) Managing the risks of extreme events and disasters to advance climate change adaptation: special report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
Isbell FI, Polley HW, Wilsey BJ (2009) Biodiversity, productivity and the temporal stability of productivity: patterns and processes. Ecol Lett 12:443–451. doi:10.1111/j.1461-0248.2009.01299.x
Jentsch A, Kreyling J, Elmer M et al (2011) Climate extremes initiate ecosystem-regulating functions while maintaining productivity. J Ecol 99:689–702. doi:10.1111/j.1365-2745.2011.01817.x
Jones MB, Donnelly A (2004) Carbon sequestration in temperate grassland ecosystems and the influence of management, climate and elevated CO(2). New Phytol 164:423–439. doi:10.1111/j.1469-8137.2004.01201.x
Kinugasa T, Tsunekawa A, Shinoda M (2012) Increasing nitrogen deposition enhances post-drought recovery of grassland productivity in the Mongolian steppe. Oecologia 170:857–865. doi:10.1007/s00442-012-2354-4
Knapp AK, Fay PA, Blair JM et al (2002) Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science 298:2202–2205. doi:10.1126/science.1076347
Knapp AK, Beier C, Briske DD et al (2008) Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience 58:811–821. doi:10.1641/B580908
Knapp AK, Hoover DL, Wilcox KR et al (2015) Characterizing differences in precipitation regimes of extreme wet and dry years: implications for climate change experiments. Glob Change Biol 21:2624–2633. doi:10.1111/gcb.12888
Kreyling J, Beier C (2013) Complexity in climate change manipulation experiments. Bioscience 63:763–767. doi:10.1525/bio.2013.63.9.12
Kreyling J, Wenigmann M, Beierkuhnlein C, Jentsch A (2008) Effects of extreme weather events on plant productivity and tissue die-back are modified by community composition. Ecosystems 11:752–763. doi:10.1007/s10021-008-9157-9
Kulmatiski A, Beard KH (2013) Woody plant encroachment facilitated by increased precipitation intensity. Nat Clim Change 3:833–837. doi:10.1038/nclimate1904
Laungani R, Knops JMH (2009) The impact of co-occurring tree and grassland species on carbon sequestration and potential biofuel production. GCB Bioenergy 1:392–403. doi:10.1111/j.1757-1707.2009.01031.x
Leuzinger S, Luo Y, Beier C et al (2011) Do global change experiments overestimate impacts on terrestrial ecosystems? Trends Ecol Evol 26:236–241. doi:10.1016/j.tree.2011.02.011
Loreau M, de Mazancourt C (2013) Biodiversity and ecosystem stability: a synthesis of underlying mechanisms. Ecol Lett 16:106–115. doi:10.1111/ele.12073
Lu M, Yang Y, Luo Y et al (2011a) Responses of ecosystem nitrogen cycle to nitrogen addition: a meta-analysis. New Phytol 189:1040–1050. doi:10.1111/j.1469-8137.2010.03563.x
Lu M, Zhou X, Luo Y et al (2011b) Minor stimulation of soil carbon storage by nitrogen addition: a meta-analysis. Agric Ecosyst Environ 140:234–244. doi:10.1016/j.agee.2010.12.010
Manning P, Newington JE, Robson HR et al (2006) Decoupling the direct and indirect effects of nitrogen deposition on ecosystem function. Ecol Lett 9:1015–1024. doi:10.1111/j.1461-0248.2006.00959.x
Mueller KE, Hobbie SE, Tilman D, Reich PB (2013) Effects of plant diversity, N fertilization, and elevated carbon dioxide on grassland soil N cycling in a long-term experiment. Glob Change Biol 19:1249–1261. doi:10.1111/gcb.12096
Mulder CPH, Jumpponen A, Hogberg P, Huss-Danell K (2002) How plant diversity and legumes affect nitrogen dynamics in experimental grassland communities. Oecologia 133:412–421. doi:10.1007/s00442-002-1043-0
Novotny AM, Schade JD, Hobbie SE et al (2007) Stoichiometric response of nitrogen-fixing and non-fixing dicots to manipulations of CO2, nitrogen, and diversity. Oecologia 151:687–696. doi:10.1007/s00442-006-0599-5
Oelmann Y, Buchmann N, Gleixner G et al (2011) Plant diversity effects on aboveground and belowground N pools in temperate grassland ecosystems: development in the first 5 years after establishment. Glob Biogeochem Cycles 25:GB2014. doi:10.1029/2010GB003869
Ordonez JC, van Bodegom PM, Witte J-PM et al (2009) A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Glob Ecol Biogeogr 18:137–149. doi:10.1111/j.1466-8238.2008.00441.x
Prescott CE (2010) Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 101:133–149. doi:10.1007/s10533-010-9439-0
Reich PB, Knops J, Tilman D et al (2001) Plant diversity enhances ecosystem responses to elevated CO2 and nitrogen deposition. Nature 410:809–812. doi:10.1038/35071062
Reichstein M, Bahn M, Ciais P et al (2013) Climate extremes and the carbon cycle. Nature 500:287–295. doi:10.1038/nature12350
Rennenberg H, Dannenmann M, Gessler A et al (2009) Nitrogen balance in forest soils: nutritional limitation of plants under climate change stresses. Plant Biol 11:4–23. doi:10.1111/j.1438-8677.2009.00241.x
Schimel D (1995) Terrestrial ecosystems and the carbon-cycle. Glob Change Biol 1:77–91. doi:10.1111/j.1365-2486.1995.tb00008.x
Schimel DS, Braswell BH, Parton WJ (1997) Equilibration of the terrestrial water, nitrogen, and carbon cycles. Proc Natl Acad Sci 94:8280–8283
Schuster MJ (2015) Increased rainfall variability and N addition accelerate litter decomposition in a restored prairie. Oecologia. doi:10.1007/s00442-015-3396-1
Skogen KA, Holsinger KE, Cardon ZG (2011) Nitrogen deposition, competition and the decline of a regionally threatened legume, Desmodium cuspidatum. Oecologia 165:261–269. doi:10.1007/s00442-010-1818-7
Smith NG, Schuster MJ, Dukes JS (2016) Rainfall variability and nitrogen addition synergistically reduce plant diversity in a restored tallgrass prairie. J Appl Ecol. doi:10.1111/1365-2664.12593
Stevens CJ, Lind EM, Hautier Y et al (2015) Anthropogenic nitrogen deposition predicts local grassland primary production worldwide. Ecology 96:1459–1465. doi:10.1890/14-1902.1
Stocker T, Qin D, Plattner G-K et al (2014) Climate change 2013: the physical science basis. Cambridge University Press, Cambridge
Tilman D, Reich PB, Knops JMH (2006) Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 441:629–632. doi:10.1038/nature04742
Turner CL, Knapp AK (1996) Responses of a C4 grass and three C3 forbs to variation in nitrogen and light in tallgrass prairie. Ecology 77:1738–1749. doi:10.2307/2265779
Vitousek PM, Aber JD, Howarth RW et al (1997) Technical report: human alteration of the global nitrogen cycle: sources and consequences. Ecol Appl 7:737–750. doi:10.2307/2269431
Yang Y, Fang J, Ma W, Wang W (2008) Relationship between variability in aboveground net primary production and precipitation in global grasslands. Geophys Res Lett 35:L23710. doi:10.1029/2008GL035408
Yue K, Peng Y, Peng C et al (2016) Stimulation of terrestrial ecosystem carbon storage by nitrogen addition: a meta-analysis. Sci Rep 6:19895. doi:10.1038/srep19895
Acknowledgments
We thank Siying Long, Raj Lal, Tanvi Lad, Alejandro Salazar, and Emmalyn Terracciano for field assistance. The PRICLE project was supported by the Purdue Climate Change Research Center (PCCRC). M.J.S. was supported by USDA Agro-ecosystem Services National Need Fellowship. N.G.S. was supported by a NASA Earth and Space Science Fellowship and a PCCRC Graduate Fellowship. J.S.D. gratefully acknowledges support from NSF (DEB-0955771). This is publication 1637 of the PCCRC.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Melany Fisk.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Schuster, M.J., Smith, N.G. & Dukes, J.S. Responses of aboveground C and N pools to rainfall variability and nitrogen deposition are mediated by seasonal precipitation and plant community dynamics. Biogeochemistry 129, 389–400 (2016). https://doi.org/10.1007/s10533-016-0240-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-016-0240-6