
Abstract
Roots influence root litter decomposition through multiple belowground processes. Hydraulic lift or redistribution (HR) by plants is one such process that creates diel drying–rewetting cycles in soil. However, it is unclear if this phenomenon influences decomposition. Since decomposition in deserts is constrained by low soil moisture and is stimulated when dry soils are rewetted, we hypothesized that diel drying–rewetting, via HR, stimulates decomposition of root litter. We quantified the decomposition of root litter from two desert shrubs, Artemisia tridentata ssp. tridentata and Sarcobatus vermiculatus, during spring and summer in field soil core treatments designed to have abundant roots and high magnitude HR cycles (DenseRoot) or few roots and low magnitude HR (SparseRoot). To help explain our decomposition results, we not only evaluated HR, but multiple factors (i.e., soil moisture, soil temperature, dissolved soil organic C concentrations, and litter chemistry) that are often influenced by roots and regulate decomposition. Root length density in the DenseRoot treatment was at least four times higher than in the SparseRoot treatment for both Artemisia and Sarcobatus by the beginning of spring. During spring and summer, there was only one instance when decomposition rates differed between the treatments. This occurred in soils beneath Artemisia in the summer when decomposition rates were 25% higher in the DenseRoot than in the SparseRoot treatments. Of the factors evaluated, only a threefold increase in the magnitude of drying–rewetting cycles created by HR in the DenseRoot compared to the SparseRoot treatment coincided with this change in decomposition. Additionally, the lower soil Ψw present in the Artemisia DenseRoot treatment should have resulted in a decline in decomposition rates, but the presence of higher magnitude HR cycles seemed to nullify this effect. There was no evidence of this result in Sarcobatus soils, possibly due to Sarcobatus only creating HR cycles for a short period of time in the summer before soil Ψw dropped below −7 MPa. As hypothesized, our results suggest that the presence of high magnitude HR cycles stimulated decomposition. The most plausible mechanism for this stimulation; however, was not solely due to HR drying–rewetting cycles but HR creating a diel rhythm of root-driven water fluxes and rhizodeposition. These together heightened microbial activity and, subsequently, enhanced the decomposition of surrounding litter. Our findings are the first field data supporting suggestions that HR influences belowground ecosystem processes and demonstrates that this relationship is seasonally variable.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Decomposition of litter in soils is both stimulated and impeded by the presences of roots. Roots stimulate decomposition through rhizodeposition. In this process, roots deposit organic compounds into the rhizosphere, thereby, enhancing microbial activity and the decomposition of surrounding litter (Kuzyakov and Domanski 2000; Kuzyakov 2002). Roots impede decomposition through extracting soil moisture and nutrients that slow microbial metabolic processes and reduce decomposition (Ehrenfeld et al. 1997; Bottner et al. 1999; Schimel et al. 2007). Besides altering these processes, roots conduct hydraulic lift or redistribution (HR) adjacent to decomposing root litter. It remains unclear; however, if this phenomenon influences decomposition.
Hydraulic redistribution creates diel drying–rewetting cycles in the rhizosphere. During the day, plant roots extract and transpire soil moisture causing soils to dry. Conversely, during the night when transpiration is reduced, HR by roots replaces this depleted soil moisture. If root systems span both wet and dry soils, HR redistributes soil moisture from wetter to drier soils as roots act as passive conduits for moisture (Richards and Caldwell 1987; Caldwell et al. 1998; Ryel 2004). This phenomenon is often thought of as a plant-centric phenomenon that confers a benefit to plants in terms of whole-plant water budgets and photosynthetic activity (e.g., Richards and Caldwell 1987; Caldwell and Richards 1989; Dawson 1993; Burgess et al. 1998; Ryel et al. 2002; Scholz et al. 2002). There has long been speculation about the effects of HR on ecosystem processes such as decomposition (Caldwell and Richards 1989; Horton and Hart 1998), yet no studies have directly tested these effects, especially in the field (Ryel 2004).
Decomposition of root litter in deserts is constrained by low soil moisture and is stimulated when dry soils are rewetted (Strojan et al. 1987; Klopatek et al. 1998; Kemp et al. 2003; Yahdjian et al. 2006). Therefore, we hypothesized that diel drying–rewetting, via HR, stimulates decomposition of root litter. We predicted that this effect was mainly present in summer, rather than spring, when soils were drier and potentially experienced higher magnitude HR diel drying–rewetting cycles.
To determine the potential influence of HR on root litter decomposition, we manipulated root density and tracked the decay of fine root litter in soils beneath two widespread cold desert shrub species, Artemisia tridentata Nutt. ssp. tridentata (Asteraceae) and Sarcobatus vermiculatus (Hook.) Torrey (Chenopodiaceae) (hereafter Artemisia and Sarcobatus). For this 2 year field experiment in a California cold desert, we used Artemisia and Sarcobatus as examples of how shrub species with different HR dynamics may influence decomposition (Caldwell et al. 1998; Donovan et al. 2003). To help explain our decomposition results, we not only evaluated the magnitude of HR diel cycles, but multiple factors (i.e., soil moisture, soil temperature, dissolved soil organic C concentrations, and litter chemistry) that are often influenced by roots and regulate decomposition. Interpretations of HR effects on decomposition are confounded by these other root effects, because HR cannot occur without roots present. However, we believe that these effects may be separated by incorporating the seasonal component of HR in tandem with root manipulations, evaluating continuous real-time measurements of soil Ψw, and comparing our decomposition results against a suite of belowground factors.
Methods
Research site, design, and harvests
This research was conducted at the Mono Basin Ecosystem Research Site (MBERS; 38°5′ N, 118°58′ W, 1,970 ± 20 m elevation a.s.l.) just northeast of Mono Lake, California. In October 2003, 40 Artemisia and 40 Sarcobatus shrubs were randomly selected from populations in the Diverse Dunes complex (see Donovan et al. 1997; Donovan and Richards 2000 for vegetation descriptions). Soils within this dune are Xeric Torripsamment with %C = 0.80, %N = 0.05, pH(H2O) = 8.6, and a soil texture of loamy sand (sand = 75%, silt = 22.5%, and clay = 2.5%) in Artemisia soils; and %C = 0.38, %N = 0.04, pH(H2O) = 9, and a soil texture of sand (sand = 92%, silt = 6%, and clay = 1%) in Sarcobatus soils. In soils beneath these 80 shrubs, two PVC tubes (15 cm diameter × 45 cm deep) were driven into the ground at the edge of the shrub’s canopy to create enclosed, undisturbed soil cores. PVC tubes were randomly placed around each shrub based on the four cardinal directions. Prior to installation, each PVC tube had 50% of its surface area removed and replaced with stainless steel mesh (either 1 mm or ≤60 μm mesh). These tubes created two root and HR manipulations for the enclosed soil cores. Firstly, a dense root treatment = DenseRoot (1 mm mesh) that allowed roots to penetrate the core from all sides, permitted direct extraction of soil moisture by roots, and encouraged HR to occur and potentially influence decomposition. Secondly, a sparse root treatment = SparseRoot (≤60 μm mesh) that discouraged roots from entering the core, prevented direct soil moisture extraction by roots (soil moisture changed by evaporation and equilibration through the fine mesh screen or the bottom of the core) and minimized possible HR effects on decomposition. Originally the SparseRoot treatment was supposed to be a no root treatment; however, a few roots penetrated the cores through the seams where the mesh contacted the outside of the tube and from the base of the core (45 cm deep). Direct comparisons between the treatments were made but comparisons to undisturbed soil were not appropriate because tube installation (e.g., root cutting) was expected to affect root density and decomposition during the experimental period.
From March 2004 until March 2005, root growth and rhizosphere processes were allowed to develop undisturbed, but at different intensities in the two treatments. In March 2005, cores were randomly assigned to be harvested either at the beginning of the second year’s spring, at the end of that spring, or at the end of summer (2 species × 2 treatments × 13–14 replicates × 3 harvests = 160 cores). The first harvest occurred immediately after snow melt and represented the beginning point for the spring period (30 March–30 June 2005) when HR was minimal. The summer period (1 July–23 September 2005) evaluated decomposition when HR was more pronounced.
Root ingrowth
To quantify root growth into the cores, root ingrowth bags (1 cm plastic mesh, 3 cm diameter × 15 cm length) were inserted into all DenseRoot cores and 5–6 randomly selected replicates of SparseRoot cores in March 2004. These cylindrical ingrowth bags were inserted by: first, removing soil from the eastern half of each core with a 3 cm soil auger; second, sieving this soil to 2 mm; third, placing this soil into the bag; and, lastly, inserting the bag into the existing hole. The volume of the ingrowth bag represented 16% of the total soil volume from 10–25 cm, which was the depth spanned by the root ingrowth bags. The hole above the bag was filled with the remaining sieved soil. At each harvest, any roots that had grown into ingrowth bags were washed clean, stored in 5% ethanol at 3°C, and scanned on a WinRHIZO 4.1C system (Regent Instrument Inc., Quebec, Canada) to determine root length density (Bauhus and Messier 1999; Bouma et al. 2000). After scanning, root biomass was determined by drying roots at 60°C to constant weight and weighing root mass.
Decomposition and litter quality
In March 2004, 1-mm-nylon-mesh litter bags (8 cm width × 10 cm length), containing 5 g of rinsed, air-dried fine roots (<3 mm diameter), were installed into the center of each soil core. These flat litter bags were inserted in the center of the cores by: first, cutting a rectangle (8 cm wide × 25 cm deep × 2 cm thick) in the soil with a soil knife; second, carefully removing this soil with a custom-made rectangular soil spatula; third, inserting the bag into the existing hole; and, lastly, refilling the hole above the bag with the removed soil. These bags spanned a similar soil depth (15–25 cm depth) as the root ingrowth bags; however, neither of these bags physically touched each other inside the core. The fine roots used in the litter bags were collected at MBERS, and only Artemisia roots were used beneath Artemisia canopies, and only Sarcobatus roots were used beneath Sarcobatus canopies.
At harvest, live roots that grew into the decomposing litter were removed from litter bags, and litter was dried at 60°C to constant weight, weighed, and ground. Litter was intentionally not rinsed to prevent loss of fine decomposed litter and leaching of soluble organic C. All litter was contaminated by soil. To correct for this contamination, all samples were fumigated with HCl to eliminate inorganic soil carbonates (Harris et al. 2001) prior to organic C analysis. The %C of litter was determined through continuous flow direct combustion and gas chromatography at the University of California Davis Stable Isotope Facility (UCD-SIF, http://stableisotopefacility.ucdavis.edu). Further, a portion of the ground root litter was ashed at 450°C for 4 h to give an ash-free mass that corrected for any inorganic soil mass (ash) that was in the sample (Mackay et al. 1987; Connin et al. 2001). The ash-free mass of each sample, along with the litter C content, was used to calculate decomposition rates. Decomposition rates were expressed as the %C loss month−1 and calculated as the difference in organic C mass in ash-free litter from the start to end of each period divided by the ash-free litter at the start of each season. These decomposition values represent potential decomposition caused by microorganisms and fungi since the 1 mm mesh size of the litter bags largely excluded soil macro- and mega-fauna (Harmon et al. 1999).
The proportions of labile (i.e., proteins, cellulose, and hemicellulose) and lignin fractions in litter were determined on a random subset of samples from each treatment (n = 5 for the start and end of the spring period, and n = 8 for the end of the summer period), via a modified Klasson lignin extraction method (Kirk and Obst 1988; Horwath and Elliott 1996).
Hydraulic redistribution, soil moisture, and soil temperature
Over the 2 years of the experiment, soil Ψw and temperature were measured hourly in soils adjacent to the litter bags (20 cm depth) in both treatments beneath six randomly selected Sarcobatus and Artemisia shrubs. These measurements were made with individually calibrated thermocouple psychrometers (Merrill Specialty Equipment, Logan, Utah) and CR7 data loggers (Campbell Scientific, Logan, Utah) (see Richards and Caldwell 1987 and Donovan et al. 2003 for details). Hydraulic redistribution and soil moisture were expressed as soil water potential (Ψw; potential energy of water in soil) due to this measurement’s universal implication for microbial stress physiology and ecosystem processes regardless of edaphic soil characteristics (Wilson and Griffin 1975; Potts 1994). These data were used to calculate the magnitude of HR diel drying–rewetting cycles and longer-term soil moisture patterns in the treatments. The magnitude of HR cycles was calculated as the diel difference in soil Ψw at 6:00 am of a given day minus the soil Ψw at 6:00 pm of the previous day. This time period incorporates the relatively wettest time of day (6:00 am) following HR during the night, and the driest period of the day (6:00 pm) following transpiration by shrubs during the day.
Dissolved organic C
Dissolved organic C (i.e., water-extractable soil organic C) (DOC; mg C kg soil−1) was evaluated on soils at the start of spring and the end of spring and summer periods. All soils were extracted within 72 h via a water extraction (1:2 w/v), passed through a 0.20 μm nylon filter, and measured on a TOC analyzer (Shimadzu, Columbia, MD). All soils were sieved to 2 mm prior to analyses.
Statistical analyses
The experimental design was a complete randomized design. Each randomly selected shrub of the two species contained both core treatments and was randomly assigned to one of the three harvests. The influence of HR by Sarcobatus and Artemisia was not compared directly, rather these serve as two examples of how HR may influence decomposition. Therefore, all treatment comparisons were conducted within species. To minimize damage to shrub root systems during insertion of the soil cores (PVC tubes), only one set of treatments was installed beneath each replicate shrub. Therefore, we did not use repeated measures to evaluate decomposition through time. Instead, we used all possible combinations of changes in root litter C for a given treatment between the start and end of each season. Two-way ANOVA by treatment and season, followed by specific contrasts, were used to determine treatment effects on decomposition rate (Zar 1996). Assumptions of ANOVA were tested using the Shapiro-Wilk test for normality and Levene’s test for homogeneity of variance. Two-way ANOVA by treatment and season, followed by specific contrasts, was also used to determine treatment effects on root characteristics, magnitude of HR diel cycles, soil moisture, and temperature. Litter chemical characteristics (e.g., labile fraction and lignin) and dissolved organic C were compared by one-way ANOVA by season for each species. Since all pair-wise combinations were of interest, means were separated using Tukey’s student range test (SAS 2001).
Results
Root characteristics in treatment cores
By the beginning of the spring period, root length density (RLD) in DenseRoot treatment was at least four-times greater than in SparseRoot for both Artemisia and Sarcobatus (Table 1). For both shrub species, roots that did penetrate the SparseRoot treatment rarely reached the litter. Within each treatment, there were no differences in RLD between spring and summer periods (Artemisia F < 0.2, P > 0.70, df = 1; Sarcobatus F < 1.1, P > 0.30, df = 1). Regardless of species and season, we observed more thick roots with many fine branches in DenseRoot cores than the very fine roots present in SparseRoot. This observation was supported by roots in the DenseRoot treatment having approximately half the specific root length (SRL) as roots in SparseRoot.
Root litter decomposition
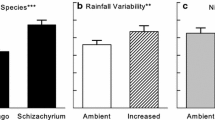
Root litter decomposition rates differed between the DenseRoot and SparseRoot treatment only in the summer period in soils beneath Artemisia (Fig. 1). During the summer period, the %C loss month−1 in the Artemisia DenseRoot treatment was 25% higher than the %C loss month−1 in the SparseRoot treatment. In the spring, there was a marginal difference in the %C loss month−1 between SparseRoot and DenseRoot treatments (F = 3.2, P = 0.07, df = 1) in Artemisia soils, with the SparseRoot treatment having higher rates than the DenseRoot treatment. Litter decomposition beneath Sarcobatus demonstrated similar trends as treatments beneath Artemisia soils; however, there were no significant differences. In addition to differences between DenseRoot and SparseRoot within a season, the %C loss month−1 in the treatments increased in Artemisia soils (F > 134, P < 0.001, df = 1) and decreased in SparseRoot Sarcobatus soils (F = 15, P < 0.001, df = 1) from the spring to the summer period. In Sarcobatus DenseRoot there was a marginal difference between decomposition rates from the spring to the summer period (F = 3.5, P = 0.06, df = 1).

Decomposition rates of root litter in Artemisia and Sarcobatus DenseRoot and SparseRoot treatments over spring and summer periods. Values are means ± 1 SE (n = 11–13). P values indicate differences between treatments within a season. The spring period was from 30 March to 30 June 2005 and the summer period was from 1 July to 23 September 2005
For the first year of the experiment, as roots established in the cores, root litter decomposition rates were similar between the two treatments (F > 3.0, P < 0.09, df = 1). Rates in soils beneath Artemisia ranged from 4.1 to 4.2 %C loss year−1 and Sarcobatus from 21 to 22 %C loss year−1 (data not shown).
Hydraulic redistribution diel cycles, and mean soil Ψw and temperature
The diel drying–rewetting cycles observed in DenseRoot and SparseRoot treatments beneath Artemisia and Sarcobatus followed the classic pattern of HR, as soil Ψw declined during the day and increased at night (Fig. 2). The magnitude of these cycles; however, only demonstrated a seasonal component in soils beneath Artemisia. Hydraulic redistribution by Artemisia in the DenseRoot treatment was more pronounced in the summer than in spring period (Fig. 3; Artemisia F = 7.8, P < 0.01, df = 1). The magnitude of HR diel cycles created by Artemisia was three-times greater in summer than in spring. Most importantly, in summer, the magnitude of HR diel cycles created by Artemisia was three-times higher in the DenseRoot than SparseRoot treatment. Sarcobatus demonstrated a similar trend in HR patterns but the difference between the DenseRoot and SparseRoot treatment was only marginally significant (F = 3.5, P = 0.08, df = 1). Hydraulic redistribution by Artemisia created cycles throughout the summer, while HR by Sarcobatus only created cycles through July. Following July, mean soil Ψw beneath Sarcobatus in the DenseRoot treatment dropped below −7 MPa, causing soil Ψw to be immeasurable with thermocouple psychrometers. Also, at the end of August, soil Ψw beneath Sarcobatus in the SparseRoot treatment also dropped below −7 MPa.

Fluctuations in soil Ψw (20 cm depth) in soils beneath Artemisia and Sarcobatus DenseRoot and SparseRoot treatments over spring and summer periods. Data are from one replicate that represent examples of the typical changes in soil Ψw for each treatment and species. The diel fluctuations present in the DenseRoot treatments in the summer period for both species are consistent with patterns of HR. Data shown were measured hourly over 24 days within each time period (spring = 4–27 June 2005 and summer = 5–28 July 2005). Vertical, dashed gray lines in each graph indicate midnight

Magnitude of HR diel cycles and average soil Ψw in Artemisia and Sarcobatus DenseRoot and SparseRoot treatments over spring and summer periods. Values are means ± 1 SE (n = 4–6). P values indicate differences between treatments within a season
In addition to creating differences in HR diel cycles, the treatments impacted mean soil Ψw. During the summer period, mean soil Ψw declined in both treatments but the mean soil Ψw in DenseRoot compared with SparseRoot declined to a much more dramatic extent (Fig. 3). This depression resulted in significant mean soil Ψw differences between treatments of 1.2 MPa for Artemisia and 2.3 MPa for Sarcobatus. During the spring period, there was a tendency for mean soil Ψw to be lower in DenseRoot relative to SparseRoot treatment in both Artemisia and Sarcobatus soils. However, this trend was not significant, despite mean soil Ψw differing at least fourfold between SparseRoot and DenseRoot treatments. Besides these differences between the treatments, mean soil Ψw declined from the spring to the summer in both Artemisia (F > 16, P < 0.01, df = 1) and Sarcobatus soils (F > 27, P < 0.001, df = 1).
Soil temperatures were similar between DenseRoot and SparseRoot treatments during the spring and summer periods. The only exception to this occurred in Artemisia soils during the summer where mean temperature in SparseRoot (20.9°C) was higher than in DenseRoot soils (19.5°C; F = 11, P < 0.01, df = 1, data not shown). Between seasons, soil temperatures were lower in the spring than in the summer period (Artemisia F > 409, P < 0.001, df = 1, Sarcobatus F > 328, P = < 0.001, df = 1). Soil temperatures beneath Artemisia were 12.5–13.4°C in the spring and 19.5–20.9°C in the summer, while temperatures beneath Sarcobatus were 9.3–9.6°C in the spring and 20.6–21.8°C in the summer (data not shown).
Litter chemistry
The chemistry of Artemisia and Sarcobatus litter, measured as % labile fraction and % lignin, were similar between DenseRoot and SparseRoot treatments at the start of the spring, end of the spring, and end of the summer periods (Artemisia F < 2.2, P > 0.15, df = 2; Sarcobatus F < 2.5, P > 0.1, df = 2). Therefore, these litter chemistry characteristics were combined and expressed by each time period for each species (Fig. 4). The only exception to this pattern occurred at the end of the summer period when root litter in Artemisia soils had a 2.5% higher lignin content in SparseRoot than DenseRoot (data not shown). Artemisia litter chemistry did vary slightly between the end of the spring and summer periods, while there were no differences in Sarcobatus chemistry between any of the time periods.

Chemistry of Artemisia and Sarcobatus root litter at the start of the spring period, end of the spring period, and end of the summer period. Labile (i.e., proteins, cellulose, and hemicellulose) and lignin fractions in litter were quantified using Klasson lignin extractions. Values are means ± 1 SE (n = 10–16). Different letters indicate significant differences (P ≤ 0.05) between time periods. The duration of each time period was as follows: start of spring = 28 March 2004–29 March 2005, end of spring = 30 March–30 June 2005, and end of summer = 1 July–23 September 2005
Dissolved organic C
The concentration of DOC in Artemisia and Sarcobatus soils were similar between DenseRoot and SparseRoot treatments at the start of the spring, end of the spring, and end of the summer periods (Artemisia F < 1.4, P > 0.29, df = 1; Sarcobatus F < 2.4, P > 0.13, df = 1, data not shown). Therefore, DOC data were combined and expressed by each time period for each species (Fig. 5). Only in Sarcobatus soils was there a change in DOC over the spring and summer periods, with DOC increasing consistently from the start of spring through the end of summer.

Dissolved organic C (i.e., water-extractable soil organic C) at the start of the spring period, end of the spring period, and end of the summer period. Values are means ± 1 SE (n = 24–27). Different letters indicate significant differences (P ≤ 0.05) between time periods. The duration of the time periods are as indicated in Fig. 4
Discussion
During spring and summer, there was only one instance when decomposition rates differed between the DenseRoot than SparseRoot treatments. This occurred in soils beneath Artemisia in the summer when decomposition rates were higher in the DenseRoot than in the SparseRoot treatments. In the DenseRoot treatment, the more than twofold increase in root density stimulated decomposition rates by 25%. Of the factors evaluated, only diel drying–rewetting cycles created through HR corresponded to the increase in decomposition rates that occurred in Artemisia soils during the summer.
As predicted, HR by Artemisia created higher magnitude drying–rewetting cycles in summer than spring, and, in summer, the magnitude of HR was higher in DenseRoot than SparseRoot treatments. The threefold increase in the magnitude of drying–rewetting cycles created by HR coincided with the higher decomposition rates that occurred in the Artemisia DenseRoot during the summer. Further, this increase in decomposition occurred even as roots were allowed to directly extract soil moisture (DenseRoot) and mean soil Ψw reached levels where microbial metabolic activity slows (Wilson and Griffin 1975; Paul and Clark 1996; Sommers et al. 1981; Fierer et al. 2003; Conant et al. 2004). The lower bulk soil Ψw present in the Artemisia DenseRoot treatment should have resulted in a decline in decomposition rates, but the presence of higher magnitude HR cycles seemed to nullify this effect. There was no evidence of this increase in root decomposition in Sarcobatus soils, possibly due to Sarcobatus only creating HR cycles for a short period of time in the summer, before soil Ψw dropped below −7 MPa. As hypothesized, our results suggest that the presence of high magnitude HR cycles, created by Artemisia, stimulated decomposition.
The presence of higher magnitude HR cycles may have stimulated decomposition through a variety of mechanisms that are not mutually exclusive. First, higher magnitude cycles may elevate the abiotic degradation of root litter, thus exposing new nutrient-rich microsites for microbial metabolism. Second, microbial communities shift in response to drying–rewetting cycles (Fierer et al. 2003; Pesaro et al. 2004), and these cycles may select for microbial communities that maintain decomposition rates despite highly variable and drier soil moisture conditions in DenseRoot treatment. However, it is unlikely that high moisture variability selects microbial communities composed of relatively more efficient or active microorganisms due to the higher energy costs associated with living in a stressful environment (Schimel et al. 2007). Lastly, Cardon and Gage (2006) proposed that HR creates a diel rhythm of root-driven water fluxes and rhizodeposition—where water transported, via HR, deposits nutrient-rich organic compounds in soils surrounding the unsuberized portion of a plant’s root system. This diel delivery of both water and new labile C substrates is the most plausible explanation for our higher decomposition rates under HR. Higher magnitude HR in the summer did not increase the overall soil moisture availability to microbes, thus, the diel delivery of water alone probably did not stimulate decomposition. Further, higher root densities and presumably more rhizodeposition in the spring did not stimulate decomposition in the absence of HR. However, the root-driven fluxes of both water and rhizodeposition simultaneously in relatively dry soils may create a synergy that accounts for the measured increase in decomposition under higher magnitude HR cycles. If HR creates both a diel rhythmic supply of water and C sources, microbial activity may be heightened and stimulate the decomposition of surrounding litter. To support this hypothesis, future research needs to, first, evaluate the amounts and timing of both water and C source delivery via HR, and, second, compare the relative contribution of HR and other soil processes to stimulating decomposition, specifically the temporal patterns of rhizodeposition.
Other soil factors, besides drying–rewetting cycles induced by HR, had little impact on decomposition or should have resulted in a higher rate of root litter decomposition in Artemisia SparseRoot than the DenseRoot treatment. For example, DOC did not differ during the summer between the two treatments, and, therefore, probably did not influence decomposition rates. During the summer, Artemisia soil temperatures also were on average 1.4°C lower in the DenseRoot than in the SparseRoot treatment. This slightly lower temperature, along with the lower soil Ψw in the DenseRoot than the SparseRoot, should have impeded decomposition in the DenseRoot treatment. However, this depression in decomposition did not occur.
In our study, we did not analyze all root processes or factors that impact decomposition. In addition to the diel fluctuations in rhizodeposition that may occur during HR, the higher density of roots in the DenseRoot treatment may have induced higher levels of fine root turnover and root exudation of labile C substrates (Marschner 1995; Kuzyakov and Domanski 2000). This subsequent ‘rhizosphere priming effect’ may have stimulated decomposition in the Artemisia DenseRoot during the summer. Unfortunately, we only partially evaluated differences in rhizodeposition in terms of total DOC. We found no differences in DOC between the treatments at the start of spring or the end of spring and summer in Artemisia soils. However, we may have missed rhizosphere deposited organic compounds that are often ephemeral in nature, excluded from water extracts since they are bound to soil particles, and only contribute a relatively small amount to total DOC concentrations.
Seasonal variation in decomposition was very apparent in all treatments beneath Artemisia and Sarcobatus. The most probable explanation for this variation was associated with seasonal differences in temperature and soil moisture availability. The most striking difference in seasonal decomposition was the more than sevenfold increase in rates between Artemisia treatments from spring to the summer. The increase of soil temperature (~10°C) from spring to summer in Artemisia soils may have influenced decomposition rates by elevating microbial activity and root litter decomposition in both treatments. Decomposition in Sarcobatus treatments demonstrated the opposite seasonal shift where rates were lower in summer than spring. The impact of elevated summer temperatures on decomposition is irrelevant without the presence of soil moisture. Sarcobatus decomposition rates were not stimulated by higher summer soil temperatures due to the relatively rapid decline of mean soil Ψw in the treatments. In all Sarcobatus soils, Ψw dropped below −7 MPa in August and were thus immeasurable with psychrometers for more than a month in the summer. This was predominantly due to differences in soil texture. Soils beneath Sarcobatus were coarser in texture than beneath Artemisia, and this texture decreased the ability of Sarcobatus soils to hold and retain moisture.
Manipulating field interactions between root growth, HR, and ecosystem processes is challenging and encompasses multiple temporal factors. For example, this study had to be 2 years to allow time for roots to grow into the treatment cores and establish in already decomposing litter. Over the first year, there were no apparent differences in litter chemistry between the treatments; however, there was no way to prohibit the decomposition of root litter during this time. Thus, much of the more labile portion of the litter was potentially degraded leaving behind more recalcitrant litter, which may have responded less dramatically. Further, differences in litter chemistry at the onset of the spring and summer periods in the second year could have influenced decomposition rates. Litter chemistry heavily impacts rates of decomposition (Meentemeyer 1978; Connin et al. 2001; Fierer et al. 2005). As litter decomposes, microorganisms preferentially used more of the labile fraction (e.g., proteins, hemi-cellulose, and cellulose), thus leaving behind a greater percentage of lignin that induces lower decomposition rates. Fortunately, the impact of litter chemistry on our results was minimal, since we found only minor differences in litter chemistry at the end of the summer.
Conclusion
Our results suggest that the presence of high magnitude HR cycles stimulated decomposition even as roots were actively depleting soil moisture. The most plausible mechanism for this stimulation; however, was not solely due to HR drying–rewetting cycles but HR creating a diel rhythm of root-driven water fluxes and rhizodeposition that together heightened microbial activity and subsequently enhanced the decomposition of surrounding litter. The influence of HR on decomposition was visible in soils beneath Artemisia but not Sarcobatus. This difference may have resulted from Sarcobatus creating HR cycles for only a short time in the summer and the extremely low soil Ψw measured in soils beneath Sarcobatus. Our findings are the first field data supporting suggestions that HR influences belowground ecosystem processes and demonstrates that this relationship is seasonally variable. It is important to note that none of the other soil factors evaluated explained our decomposition results. To solidify our findings, research needs to compare the relative contribution of HR and other soil processes to stimulating decomposition, especially differentiating the impact of rhizodeposition in the presence and absence of HR in relatively dry soils. Due to the wide and prevalent distribution of Artemisia and Sarcobatus in Great Basin, Mojave, and Navajoan Deserts (Benson and Darrow 1981; Baldwin et al. 2002), and the role of halophytes (i.e., Sarcobatus) and non-halophytes (i.e., Artemisia) in worldwide deserts, these results have broad implications for C storage and cycling in arid and semi-arid ecosystems. Additionally, many plant species, besides desert shrubs, conduct HR (Caldwell et al. 1998; Horton and Hart 1998). These results may offer insights into C dynamics in other ecosystems where HR is prevalent.
References
Baldwin BG, Boyd S, Fritter BJ, Patterson RW, Rosatti TJ, Wilken DH (2002) The Jepson desert manual. University of California Press, Berkeley
Bauhus J, Messier C (1999) Evaluation of fine root length and diameter measurements obtained using RHIZO image analysis. Agron J 91:142–147
Benson L, Darrow RA (1981) Trees and shrubs of the southwestern deserts. University of Arizona Press, Tuscon
Bottner P, Pansu M, Sallih Z (1999) Modelling the effect of active roots on soil organic matter turnover. Plant Soil 216:15–26
Bouma TJ, Nielsen KL, Koutstaal B (2000) Sample preparation and scanning protocol for computerized analysis of root length and diameter. Plant Soil 218:185–196
Burgess SSO, Adams MA, Turner NC, Ong CK (1998) The redistribution of soil water by tree root systems. Oecologia 115:306–311
Caldwell MM, Richards JH (1989) Hydraulic lift—water efflux from upper roots improves effectiveness of water-uptake by deep roots. Oecologia 79:1–5
Caldwell MM, Dawson TE, Richards JH (1998) Hydraulic lift: consequences of water efflux from the roots of plants. Oecologia 113:151–161
Cardon ZG, Gage DJ (2006) Resource exchange in the rhizosphere: molecular tools and microbial perspectives. Annu Rev Ecol Evol S 37:459–488
Conant RT, Dalla-Betta P, Klopatek CC, Klopatek JM (2004) Controls of soil respiration in semiarid soils. Soil Biol Biochem 36:945–951
Connin SL, Feng X, Virginia RA (2001) Isotopic discrimination during long-term decomposition in an arid land ecosystem. Soil Biol Biochem 33:41–51
Dawson TE (1993) Hydraulic lift and water-use by plants—implications for water-balance, performance and plant-plant interactions. Oecologia 95:565–574
Donovan LA, Richards JH (2000) Juvenile shrubs show differences in stress tolerance, but no competition or facilitation, along a stress gradient. J Ecol 88:1–16
Donovan LA, Richards JH, Schaber EJ (1997) Nutrient relations of the halophytic shrub, Sarcobatus vermiculatus, along a soil salinity gradient. Plant Soil 190:105–117
Donovan LA, Richards JH, Linton MJ (2003) Magnitude and mechanisms of disequilibrium between predawn plant and soil water potentials. Ecology 84(2):463–470
Ehrenfeld JG, Parsons WFJ, Han X, Parmelee RW, Zhu W (1997) Live and dead roots in forest soil horizons: contrasting effects on nitrogen dynamics. Ecology 78(2):348–362
Fierer N, Schimel JP, Holden PA (2003) Influence of drying-rewetting frequency on soil bacterial community structure. Microb Ecol 45:63–71
Fierer N, Craine JM, McLauchlan K, Schimel JP (2005) Litter quality and the temperature sensitivity of decomposition. Ecology 86(2):320–326
Harmon ME, Nadelhoffer KJ, Blair JM (1999) Measuring decomposition, nutrient turnover, and stores in plant litter. In: Robertson PG, Coleman DC, Bledsoe CS, Sollins P (eds) Standard soil methods for long-term ecological research. Oxford University Press, New York
Harris D, Horwath WR, van Kessel C (2001) Acid fumigation of soils to remove carbonates prior to total organic carbon or carbon-13 isotopic analysis. Soil Sci Soc Am J 65:1853–1856
Horton JL, Hart SC (1998) Hydraulic lift: a potentially important ecosystem process. Trends Ecol Evol 13:232–235
Horwath WR, Elliott LF (1996) Ryegrass straw component decomposition during mesophilic and thermophilic incubations. Biol Fertil Soil 21:227–232
Kemp PR, Reynolds JF, Viginia RA, Whitford WG (2003) Decomposition of leaf and root litter of Chihuahuan desert shrubs: effects of three years of summer drought. J Arid Environ 53:21–39
Kirk TK, Obst JR (1988) Lignin determination. Meth Enzymol 161:87–101
Klopatek JM, Conant RT, Francis JM, Malin RA, Murphy KL, Klopatek CC (1998) Implications of patterns of carbon pools and fluxes across a semiarid environmental gradient. Landsc Urban Plan 39:309–317
Kuzyakov Y (2002) Review: factors affecting rhizosphere priming effects. J Plant Nutr Soil Sci 165:382–396
Kuzyakov Y, Domanski G (2000) Carbon input by plants into the soil. Review. J Plant Nutr Soil Sci 163:421–431
Mackay WP, Fisher FM, Silva S, Whitford WG (1987) The effects of nitrogen, water and sulfur amendments on surface litter decomposition in the Chihuahuan Desert. J Arid Environ 12:223–232
Marschner H (1995) Mineral nutrition of higher plants, 2nd edn. Academic Press, London
Meentemeyer V (1978) Macroclimate and lignin control of litter decomposition rates. Ecology 59(3):465–472
Paul EA, Clark FE (1996) Soil microbiology and biochemistry, 2nd edn. Academic Press, San Diego
Pesaro M, Nicollier G, Zeyer J, Widmer F (2004) Impact of soil drying-rewetting stress on microbial communities and activities and on degradation of two crop protection products. Appl Environ Microb 70:2577–2587
Potts M (1994) Desiccation tolerance of prokaryotes. Microbiol Rev 58:755–805
Richards JH, Caldwell MM (1987) Hydraulic lift—substantial nocturnal water transport between soil layers by Artemisia tridentata roots. Oecologia 73:486–489
Ryel RJ (2004) Hydraulic redistribution. Prog Bot 65:413–435
Ryel RJ, Caldwell MM, Yoder CK, Or D, Leffler AJ (2002) Hydraulic redistribution in a stand of Artemisia tridentata: evaluation of benefits to transpiration assessed with a simulation model. Oecologia 130:173–184
SAS (2001) SAS/STAT user’s guide, Version 8. SAS Institute, Cary
Schimel JP, Balser TC, Wallenstein M (2007) Microbial stress-response physiology and its implications for ecosystem function. Ecology 88(6):1386–1394
Scholz FG, Bucci SJ, Goldstein G, Meinzer FC, Franco AC (2002) Hydraulic redistribution of soil water by neotropical savanna trees. Tree Physiol 22:603–612
Sommers L, Gilmour C, Wildung R, Beck S (1981) The effect of water potential on decomposition processes in soil. In: Parr J, Gardner W, Elliott L (eds) Water potential relations in soil microbiology. Soil Science Society of America, Madison
Strojan CL, Randall DC, Turner FB (1987) Relationship of leaf litter decomposition rates to rainfall in the Mojave Desert. Ecology 68(3):741–744
Wilson JM, Griffin DM (1975) Water potential and the respiration of microorganisms in the soil. Soil Biol Biochem 7:199–204
Yahdjian L, Sala O, Austin A (2006) Differential controls of water input on litter decomposition and nitrogen dynamics in the Patagonian steppe. Ecosystems 9:128–141
Zar JH (1996) Biostatistical analysis, 3rd edn. Prentice-Hall, Upper Saddle River
Acknowledgments
We thank the Kearney Foundation of Soil Science (2003.316) and the California Agricultural Experiment Station for funding this research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Aanderud, Z.T., Richards, J.H. Hydraulic redistribution may stimulate decomposition. Biogeochemistry 95, 323–333 (2009). https://doi.org/10.1007/s10533-009-9339-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-009-9339-3