
Abstract
The deposition of nitrogen (N) is high in subtropical forest in South China and it is expected to increase further in the coming decades. To assess effects of increasing deposition on N cycling, we investigated the current N status of two selected 40–45-year-old masson pine-dominated Chinese subtropical forest stands at Tieshanping (TSP, near Chongqing City) and Caijiatang (CJT in Shaoshan, Hunan province), and explored the applicability of several indicators for N status and leaching, suggested for temperate and boreal forest ecosystems. Current atmospheric N deposition to the systems is from 25 to 49 kg ha−1 year−1. The concentration of total N in the upper 15 cm of the soil is from as low as 0.05% in the B2 horizon to as high as 0.53% in the O/A horizon. The concentration of organic carbon (C) varies from 0.74 (B2) to 9.54% (O/A). Pools of N in the upper 15 cm of the soils range from 1460 to 2290 kg N ha−1, where 25–55% of the N pool is in the O/A horizon (upper 3 cm of the soil). Due to a lack of a well-developed continuous O horizon (forest floor), the C/N ratio of this layer cannot be used as an indicator for the N status, as is commonly done in temperate and boreal forests. The net N mineralization rate (mg N g−1 C year−1) in individual horizons correlates significantly with the C/N ratio, which is from as high as 18.2 in the O/A horizon to as low as 11.2 in the B2 horizon. The N2O emission flux from soil is significantly correlated with the KCl extractable NH +4 –N in the O/A horizon and with the net nitrification in the upper 15 cm of the soil. However, the spatial and temporal variation of the N2O emission rate is high and rates are small and often difficult to detect in the field. The soil flux density of mineral N, defined as the sum of the throughfall N input rate and the rate of in situ net N mineralization in the upper 15 cm of the soil, i.e., the combination of deposition input and the N status of the system, explains the NO −3 leaching potential at 30 cm soil depth best. The seasonality of stream water N concentration at TSP and CJT is climatic and hydrologically controlled, with highest values commonly occurring in the wet growing season and lowest in the dry dormant season. This is different from temperate forest ecosystems, where N saturation is indicated by elevated NO −3 leaching in stream water during summer.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In the past decades, anthropogenically derived reactive nitrogen (N) has exceeded natural N input through biological N fixation as the dominant N source in many forest areas (e.g., Emmett et al. 1998; Matson et al. 1999; Galloway et al. 2002; Aber et al. 2003). In forest ecosystems in subtropical south China, current deposition of N is high and of the same order of magnitude as in northwest Europe, ranging from 4 to 40 kg N ha−1 year−1 (Chen et al. 2004). In the coming decades, further increases of N deposition in subtropical regions in south China are to be expected due to elevated NH3 emissions from agriculture and increasing NOx emissions from fossil fuel combustion, and biomass burning (Zhao and Wang 1994; Zhu and Chen 2002; Hao et al. 2002; Galloway et al. 2002).
Increased N deposition alters the N status of forest ecosystems through disturbances of the internal N cycle (Gundersen et al. 1998a). Eventually, elevated N deposition may cause N saturation and increased NO −3 leaching loss from forest ecosystems (Ågren and Bosatta 1988; Aber et al. 1998). Yet, the response of forest ecosystems to atmospheric N deposition may differ according to climate, species composition, and initial N status of the ecosystem (Dise and Wright 1995; Gundersen et al. 1998a, b; MacDonald et al. 2002; Hall and Matson 2003; Aber et al. 2003). Aber et al. (1998) defined four stages (0–3) to describe the N saturation of forests, each stage being characterized by different symptoms, including changes in N concentrations, pools, and fluxes. The N status of forests, defined as the degree of N saturation (e.g., Currie 1999; MacDonald et al. 2002), can be used to further quantify the condition of forest ecosystems with respect to N saturation.
Several indicators have been proposed, which empirically relate deposition, status, and leaching of N for temperate and boreal forest (Gundersen 1991; Dise et al. 1998b; Andersson et al. 2002; Bengtsson et al. 2003). The N concentration and C/N ratio of the forest floor as well as the in situ N mineralization and nitrification rate were identified as key indicators of the ecosystem’s “nitrogen status” (Gundersen et al. 1998a; Currie 1999; Andersson et al. 2002). Based on data from the NITREX sites, Gundersen et al. (1998a) concluded that although the mineral soil has a large capacity to retain N, changes in forest floor and vegetation are paramount to explain the increased NO −3 leaching from temperate and boreal forest soils. Researches along N deposition gradients in the USA and Europe revealed a negative correlation between the C/N ratio in the organic horizon and N mineralization, nitrification, and leaching rates (Fenn et al. 1996; Tietema et al. 1997; Dise et al. 1998b; Wright et al. 1998). Recently, the soil flux density of mineral N, defined as the sum of the atmospheric N deposition (as determined in throughfall) and net N mineralization rates, has been proposed as an indicator for NO −3 leaching from boreal forest soils (Andersson et al. 2002). In addition, gaseous emission of nitrous oxide (N2O), resulting from both aerobic nitrification and anaerobic denitrification, has been found to increase with increasing N status of terrestrial ecosystems (Bouwman et al. 1993). Thus, the N2O emission flux is proposed as a nondestructive indicator of the N status of forest soil (Parsons et al. 1996; Hall and Matson 2003)
In temporal and boreal-forested catchments, N saturation stages are defined by levels and seasonality of NO −3 concentrations in stream water. Increases in N saturation result in a shift in NO −3 concentration from low in summer and high in winter to high all year round. The early stages of N saturation are characterized by increases in the severity and frequency of NO −3 episodes in the growing season, whereas later stages also have elevated base flow concentrations of NO −3 (Stoddard 1994; Dise et al. 1998a).
Forest ecosystems in subtropical south China are different from those in temperate and boreal regions, e.g., with respect to climate, species composition, and soil properties. However, little is known about status, dynamics, and leaching risk of N in these subtropical forests. Information on current N status and N leaching are urgently needed for assessment of the effects of increasing N deposition on N cycling in these ecosystems. Within the 5-year Sino–Norwegian cooperated Integrated Monitoring Program on Acidification of Chinese Terrestrial Systems (IMPACTS; Larssen et al. 2004) project, a detailed N study was conducted in two coniferous-broad leaved mixed Chinese subtropical forests, dominated by masson pine, at TieShanPing (TSP, near Chongqing City) and CaiJiaTang (CJT in Shaoshan, Hunan province). The objectives of this paper are: (1) investigate the current N status and N leaching of two selected subtropical Chinese forest stands and (2) test the applicability of different indicators for N status and leaching to subtropical Chinese forest ecosystems.
Methods
Site descriptions
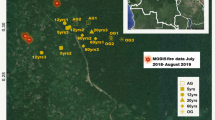
In total five monitoring sites were selected for IMPACTS (Fig. 1). At two of these sites, TieShanPing (TSP) and CaiJiaTang (CJT) stands dominated by masson pine were selected for detailed N research. The main characteristics of TSP and CJT are as show in Table 1 (for more details, see Larssen et al. 2004).

Location of the research sites TieShanPing (TSP) and CaiJiaTang (CJT), China
At both sites, soils, dominated by Haplic Acrisols, are acidic (pH 3.5–3.8 in the O/A horizon and 3.8–4.0 in the B). Base saturation decreases from an average of 26% (TSP) and 38% (CJT) in the O/A horizon to 9% (TSP) and 16% (CJT) in the B. Due to rapid decomposition a continuous forest floor is lacking. Both at TSP and CJT, an O/A horizon, on average 2–4 cm thick, is situated on top of a deep B horizon. At a depth of 40–50 cm there is a gradual transition to the C horizon. At CJT, but not at TSP, the soils contain a considerable amount of stones.
Monitoring program and chemical analysis
Bulk deposition, throughfall, soil water and stream water chemistry data are for 2001–2003. For detailed sampling procedures and chemical methods reference is made to (Chen et al. 2004). Data were collected at four (TSP) and three (CJT) monitoring plots, dominated by Masson pine. Throughfall was collected below the shrub vegetation using four collectors per monitoring plot. Weekly samples from all four collectors were pooled to one volume weighted monthly sample per plot and stored in the refrigerator prior to analysis. At each monitoring plot soil water was sampled, using one zero tension lysimeter (30 × 30 cm2) at 0 cm soil depth (below freshly-fallen litter) to collect the water infiltrating into the mineral soil. In addition, two ceramic tension lysimeters (50 kPa) were used to sample soil water from each of three to four layers of the mineral soil. Stream water discharge was registered continuously using a V-notch weir and stream water samples were taken at weekly intervals. Major cations and anions, including ammonium (NH +4 ) and nitrate (NO −3 ) were analyzed as described in ISO14911 and ISO10304-1, using ion chromatography. Annual water fluxes of throughfall and L0 lysimeter were estimated according to the recorded weekly water volume and surface area of collectors and zero tension lysimeters, respectively. Annual fluxes of solutes were calculated by multiplying annual volume weighted concentrations with the corresponding water fluxes during the same period. The annual average of soil solution concentration at 30 cm soil depth was used to estimate the leaching potential at this depth. Annual solute fluxes in stream water were calculated using the catchment area and volume-weighted mean of weekly concentrations and annual total discharge.
In situ N transformation rates
In situ N transformation rates were determined after incubation in the field. Three (CJT) and four (TSP) incubation plots, each adjacent to one of the lysimeter plots for soil water monitoring, had masson pine (Pinus massoniana) as the dominant tree species. Net N transformation rates were quantified during five subsequent intervals in the period May 2002 to May 2003, yielding annual in situ net N transformation rates. Incubation periods were 2 months during the growing season (May to end of September 2002; March to May 2003) and 3 months during the winter season (October through March 2003). Soil cores (diameter 7 cm, length 15 cm) were taken, after removal of freshly fallen, but not partly decomposed litter, with a sharpened steel auger, lined with a PVC cylinder. Removal of freshly-fallen litter implies that our data exclude mineralized N from the initial phase of litter decomposition. For each incubation period one set of nine soil cores (background) was brought back to the laboratory for immediate extraction of NH +4 and NO −3 . A duplicate set of nine soil cores were incubated in the field. The tops of the PVC cylinders were covered with a perforated plastic sheet, thus allowing gas exchange, but preventing infiltration of rain water. At the end of each incubation period, the incubated samples were collected. Each one of the nine soil cores was partitioned into different horizons, i.e., the O/A, B1, and B2 horizons and the depth per horizon were recorded. The individual horizons from three cores were pooled in plastic bags and transferred to the laboratory. In total there were three pooled samples from each O/A, B1, and B2 horizon per plot for each study period. The total weight (<2 mm fraction) was determined for each horizon and sieved subsamples (<2 mm) were extracted with 2 M KCl to recover NH +4 and NO −3 . After filtration (ashless paper filter, microscience, Germany), the extracts were analyzed colorimetrically with respect to NH +4 –N and NO −3 –N. For further details reference is made to Chen et al. (2004).
In background soil samples total C and N were determined using a CHN-analyzer as described in ISO10694 (1995) and ISO 13878 (1998), respectively. Using the C and N concentration, together with horizon depth, the total weight and moisture content of the soil (< 2mm fraction) obtained for each horizon and the volume of the PVC cylinder, the total C and N pools were calculated. Data are presented on per weight and per area basis.
Net ammonification and nitrification for each horizon are calculated separately using the equation Nt = Ni−Np, where Nt is the net transformation of NH +4 (ammonification) and NO −3 (nitrification), respectively, Ni is the NH +4 –N and NO −3 –N concentration in the incubated soil core, and Np is the concentration of NH +4 –N and NO −3 –N prior to incubation. The sum of net ammonification and net nitrification equals net mineralization. Rates were determined by dividing Nt with in situ incubation time.
In situ N2O flux
The collection method for gas samples is as described by Sitaula et al. (1995). Briefly, atmospheric gas samples were collected at different plots at TSP and CJT during summer 2002 and 2003 using the enclosure method. Two cylinders (height 20 cm, diameter 20 cm) were pressed with their opening into the soil surface (about 2 cm) and the bottom up. The bottom was equipped with three holes. Prior to inserting the cylinder into soil, all above ground parts of plants were removed. Care was taken that the soil–cylinder interface was free of leaks to prevent gas exchange with the atmosphere after deployment. Finally, the holes in the cylinder were closed with rubber septa and immediately a gas sample was collected from the headspace with a syringe, representing the background. After 15 and 30 min, the headspace was resampled. All gas samples were transferred into 12 ml preevacuated vials. N2O concentrations in the vials were measured using gas chromatography as described by Sitaula et al. (1995).
Quality control and statistical analysis
The quality control of analysis in different IMPACTS laboratories involves occasional ring-test. Results indicate that analytical differences between the various laboratories were relatively small. All statistical analyses were performed using SPSS 11.1 for Windows (SPSS, Inc., 2001). Pearson correlations were tested using bivariate correlation functions, and were considered significant if p < 0.05.
Results and discussion
N concentration and N pool size in the soils
The N concentration in the upper 15 cm of the mineral soils at TSP and CJT ranges from as low as 0.05% in the B2 to as high as 0.53% in the O/A (Table 2). The organic C concentration, ranging from 0.74 to 9.54% and from 2.12 to 7.19% at TSP and CJT, respectively, correlates significantly with the total N concentration in all horizons (r = 0.993, p < 0.001; data not shown).
Average N pools in the upper 15 cm of the soils at TSP and CJT vary from 1460 to 2290 kg N ha−1. These values are similar to the range of values reported for tropical montane forest soils (870–2100 kg N ha−1, Vitousek and Matson 1988) and humid subtropical forest stands in India (Maithani et al. 1998). In temperate forest the forest floor (O horizon) contributes as much as 50% to total N pool. However, soils at TSP and CJT lack a continuous and well-developed forest floor with decomposing litter. The N pools in the upper 15 cm at TSP and CJT are of the same order of magnitude as those of mineral horizons in boreal forests. The pool of organic C in the upper 15 cm of the mineral soil at TSP and CJT is nearly half of that in mineral horizons of boreal forest soil (Andersson et al. 2002; Schulze 2000). Thus, soil organic matter at TSP and CJT has a lower C/N than boreal forests. Nitrogen in the O/A horizon (0–3 cm depth) at TSP and CJT contributes 25–55% to the total N pool of the upper 15 cm of the soil (Table 2).
C/N ratio
The soil C/N ratio varies among the plots and decreases with depth at all plots at TSP and CJT (Table 2). The lower C/N ratios in deeper soil layers are mainly due to the decreasing organic C concentration along the soil profile. C/N ratios of all horizons in the upper 15 cm of the soil are in the range of 11.2–18.2. These values are consistent with the reported global trends of C/N in different life zones. Globally, C/N ratios of surface soil horizons range from <10 in tropical deserts to >20 in cool, wet forests. Relatively large amounts of N in surface soil in tropical and subtropical regions are associated with recalcitrant humic material in advanced state of decay with low C/N (Post et al. 1985).
Net N mineralization and nitrification
Average annual net N mineralization rates in the upper 15 cm of the soils (i.e., excluding freshly fallen litter) are CJT 52 kg N ha−1 year−1 and TSP 62 kg N ha−1 year−1 (Table 3) with a contribution of about 30% from the O/A horizon. Our data suggest that about 2.8–3.5% of the N pool contributes to net mineralization per year (Tables 2 and 3) and more than half of this is further nitrified (Table 3). Net nitrification rates are substantial at both sites, with average values for the upper 15 cm of the mineral soil of about 35 kg N ha−1 year−1 (Table 3). Along the profile, the nitrification rate increases with the depth. On average, the O/A horizon contributes about 30% to total net nitrification in the upper 15 cm of the soils.
The in situ mineralization and nitrification rates presented here do not include initial contributions from decomposing freshly fallen litter, as this was removed from the incubation cores prior to covering the tops with perforated plastic caps. Since, the freshly deposited litter is rapidly decomposed, leachate from the 0 cm percolation lysimeter (Table 4) may be assumed to include net mineralization products from litter decomposition. The average net N production in this layer is 10 and 23 kg N ha−1 year−1 for TSP and CJT, respectively (data not shown). However, probably this represents an overestimate of the contribution to N mineralization of freshly fallen litter that was removed from the soil cores. So, net N mineralization rates, including the litter layer, at TSP and CJT may be as high as 72 and 75 kg N ha−1 year−1, respectively. Andersson et al. (2002) reported N mineralization rates of 51 kg N ha−1 year−1 for some boreal forest stands. In boreal forest soils, however, the forest floor contributes as much as 80 and 30% to net N mineralization and nitrification, respectively.
The net mineralization rate, expressed in mg N g−1 C year−1, increases with soil depth at TSP and CJT. Mean values range from as low as 1.5 (A/O) to as high as 3.6 (B1) mg N g−1 C year−1. The increase with soil depth of N mineralization rate per gram soil organic C is different from the pattern found in boreal forest soils (Andersson et al. 2002) with our values being several times greater. This suggests a greater microbial activity in the mineral soils of subtropical forests compared with boreal systems. In addition, it may imply that the substrate in this subtropical forest soil has better quality (related to C/N ratio) and the C use efficiency of microorganisms is greater than that in boreal and temperate forest soils.
In situ soil N2O fluxes
N2O fluxes show high spatial and temporal variations (Fig. 2a, b). During the growing season of 2002 the estimated fluxes (data not shown) at CJT are in the range of 0.92–26.8 μg N2O–N m−2 h−1. At TSP these values are from 3.4 to 59.1 μg N2O–N m−2 h−1. These values are comparable to values reported by Butterbach-Bahl et al. (1997, 2002) for pine stands in the Netherlands (6–53 μg N m−2 h−1), but greater than those presented by Oura et al. (2001) for Japanese red pine stands (0.5–14.1 μg N m−2 h−1). The large spatial variability of N2O emission fluxes reflects the diversity of micro sites and the heterogeneity of the soil.

Spatial and temporal variation of soil N2O emission flux. (a) Temporal variation. Values are averages for seven plots at TSP and CJT. Measurement dates: 28 May, 29 July, 20 September 2002 at TSP and 21 May, 21 July, and 24 Sept 2002 at CJT; (b) Spatial variation. Values are the average of observations in each plot during May, July, and September 2002
Atmospheric input and leaching of N
Table 4 shows the total atmospheric N deposition input, total inorganic N concentration in soil water at 0 and 30 cm depth, and the total inorganic N flux in stream water for 2001, 2002, and 2003. The total atmospheric N input to the soil is estimated by the throughfall flux of N collected below the ground vegetation (FTF). Throughfall fluxes of N, which are considerably greater than wet-only deposition fluxes, vary greatly between plots, ranging from 25 to 51 kg N ha−1year−1. On average, the atmospheric deposition of N at the two sites is similar, ranging from 36 to 44 kg N ha−1year−1. Total inorganic N concentration in soil water at 30 cm depth also varies from plot to plot, ranging from 3.3 to 18.3 mg l−1, with plot K at TSP having the highest values. This plot also has the highest annual net N mineralization rate (Table 3).
Mean weekly concentrations of total inorganic N (TIN; NO −3 –N+NH +4 –N) and total N (sum of total inorganic and dissolved organic N) in stream water during the period of 2001–2003 are 1.04 and 1.13 mg N l−1 for TSP and 0.43 and 0.59 mg N l−1 for CJT (Fig. 3). Both total N and total inorganic N concentrations at TSP are significantly greater than those at CJT (Fig. 3, p < 0.001, and p < 0.001, respectively). Nitrate contributes more than 90% to total inorganic N at both sites, whereas the contribution of NO −3 –N to total N in stream water is 90 and 70% at TSP and CJT, respectively. Inorganic N fluxes in stream water range from as low as 2.4 at CJT to as high as 10.8 kg N ha−1 year−1 at TSP (Table 4), with TSP having the greatest fluxes in 2002 (the wettest year).

Total inorganic (TIN) and total N (Tot_N) in stream water at TSP and CJT. Both total inorganic N and total N is dominated by NO −3 –N. Error bars show the standard deviation
Indicators for N status
Several indicators for N status have been identified for temporal and boreal forest ecosystems. Most commonly used are N concentration (Fernandez et al. 2000) and C/N ratio of the forest floor (e.g., Gundersen et al. 1998b; Dise et al. 1998b; Currie 1999; Andersson et al. 2002; Bengtsson et al. 2003). As indicated previously, soils at TSP and CJT lack a well-developed forest floor, so that other indicators for N status have to be assessed.
At TSP and CJT, the N pool in the upper 15 cm of the soil is not significantly correlated with in situ net N mineralization rate (data not shown). The N pool in the O/A horizon correlates weakly with both net mineralization rate and net nitrification rate in the upper 15 cm of the soil (Fig. 4a, b), the correlation with the net nitrification rate shows a trend towards significant (p < 0.1, Fig 4b). This may suggest that the amount of N in the O/A horizon has a positive effect on net rates of N transformation and nitrification in the upper 15 cm of the soil. The C/N ratio of the O/A horizons at TSP and CJT is not significantly correlated with either the total net N mineralization or nitrification rate in the upper 15 cm of the soil (Fig. 5a). Thus the C/N ratio in O/A horizon cannot be used as indicator for the N status in these subtropical forest ecosystems.

The relation between N pool in O/A horizon at TSP and CJT and net annual N mineralization rate (a) and net nitrification rate (b) in the upper 15 cm of the soil

Relation between C/N ratio and the net N transformation rate (a). Relation between the C/N ratio of individual O/A, B1, and B2 horizons and the net N mineralization rate (b) (r = −0.495, p = 0.022) and the net nitrification rate (c) (r = −0.546, p = 0.011). The figures show the correlation lines as well as the 95% confidence interval
The net N mineralization rate and net nitrification rate per unit organic C (mg N g−1 C year−1) in each horizon is significantly correlated with the C/N ratio in that horizon (r = −0.495, p = 0.022, and r = −0.546, p = 0.011, respectively, Fig. 5b, c). Thus the C/N ratio in mineral soils may be used as a predictor of net N mineralization rate in these subtropical forest soils. Correlations suggest that a C/N between 20 and 24 is required before net N mineralization and nitrification may occur in the mineral soil of Chinese subtropical forests. This threshold value is consistent with that for organic horizons in temperate forest soil (Ollinger et al. 2002). Additional studies of N transformation in soils with a greater range of C/N ratios are necessary to confirm the threshold values.
This correlation for mineral soil horizons is inconsistent with findings for temperate and boreal forest soils. In temperate and boreal forest soils, the C/N ratio of the forest floor was found to correlate negatively with net mineralization rate per unit organic C (Andersson et al. 2002). The difference between temperate and boreal forest soils compared with subtropical forest soils may be explained by the difference in the distribution pattern of C and N along the soil profile. In temperate forest ecosystems, the dominant role of the forest floor can be attributed to the high contribution of C and N in the forest floor to the total pool of soil organic matter. By contrast, the correlation in our datasets may be explained by the similar turnover rate of C and N in all three horizons of the upper 15 cm soil. Previously, the C/N of soil organic matter has been proposed as a mechanistic indicator of the quality or degradability of the substrate in the soil (Curri 1999; Fernandez et al. 2000; Bengtsson et al. 2003; Theodose and Martin 2003). Also in the mineral horizons of our soils, the correlation between net transformation rates and the C/N may be related to the substrate quality along the soil profile. Ultimately, it may be attributed to the source and stability of the soil organic matter along the soil profile of upper 15 cm.
Previously, N2O emissions were found to correlate positively with parameters indicating soil N status, including the pool of mineral N, total N concentration, NO −3 concentration and nitrification rate in the soil (Hall and Matson 2003; Parsons et al. 1996). At our Chinese sites, the N2O flux is not significantly correlated with the N concentration in any of the horizons (data not shown). However, the N2O flux is significantly correlated with KCl extractable NH +4 –N in the O/A horizon (Fig. 6a) and with the net nitrification rate in the upper 15 cm of the soil (Fig. 6b). This is consistent with finding from a tropical forest in Atherton Tablelands, Australia, where gross nitrification was found to correlate positively with in situ N2O-emission rates (Breuer et al. 2000), even though it should be noted that the pattern for gross nitrification is not always the same as that for net nitrification. The significant correlation between N2O flux and net nitrification rate suggests that the N2O flux also may be employed as indicator of N status of subtropical forest ecosystems. However, the great spatial and temporal variations of N2O emission (Fig. 2a, b) make this parameter difficult to use.

Soil N2O emission flux as related to KCl exchangable NH +4 –N in O/A horizon (a) and the net nitrification rate in the upper 15 cm of the soil (b)
Indicators for N leaching
Elevated NO −3 loss to groundwater or surface waters was defined as the primary symptom of N excess in forest ecosystem, due to increased anthropogenic N input (Aber et al. 1998). From two European field-scale manipulation projects (NITREX and EXMAN), a correlation between N deposition and leaching was found and a threshold of atmospheric input rate was defined in the range of 10–30 kg N ha−1 year−1 (MacDonald et al. 2002). In addition, NO −3 leaching from coniferous forest across Europe was found to depend on the forest floor C/N ratio (Dise et al. 1998b). MacDonald et al. (2002) proposed a forest floor C/N ratio of 25 as a threshold for NO −3 leaching. The average annual input of N at TSP and CJT is around 40 kg N ha−1 year−1 (Table 4, Chen et al. 2004) and well above the threshold for atmospheric deposition rates that give rise to NO −3 leaching in boreal and temperate forests. At TSP and CJT, the total throughfall input of mineral N into the forest soils has no apparent correlation with the TIN concentration at 30 cm soil depth (Fig. 7a). Neither do C/N ratios of the O/A horizon correlate significantly with total inorganic N concentration at the 30 cm soil depth (Fig. 7b). This suggests that neither throughfall N deposition nor C/N ratio of the O/A horizon explains N leaching at TSP and CJT.

Correlation between concentration of total inorganic N (NH +4 –N + NO −3 –N, TIN) and various factors at TSP and CJT. Data in (a) and (b) are from 2001 to 2003, others are from May 2002 to May 2003 (the same period as the mineralization study). The soil flux density of mineral N (d, e, f) is defined as the sum of the throughfall input of inorganic N and the net mineralization rate in the upper 15 cm of the soil. Alternatively, the flux density of mineral N* was defined as the sum of the input of inorganic N in the O/A horizon (as determined by the tension lysimeter at 0 cm soil depth) and the net mineralization rate in the upper 15 cm of the mineral soil
The N mineralization rate, an important process regulating N dynamics, has been suggested as an index for potential risk of N leaching in boreal forest (Andersson et al. 2002). At TSP and CJT, both total annual net N mineralization rate (Fig. 7c) and nitrification rate (not shown) in the upper 15 cm of the soil show significant correlation with the total inorganic N concentration at the 30 cm soil depth. This suggests that N transformations are important contributing factors to N leaching in these subtropical forests. As the N transformation rates are the physiological symptoms for N status of the system, it also indicates that N leaching from these forests is closely related to the N status of the systems.
Recently, the soil flux density of mineral N, defined as the sum of total atmospheric N input and net N mineralization, has been proposed as an indicator for NO −3 leaching in boreal forest soils (Andersson et al. 2002). Also at TSP and CJT it appears that the soil flux density of mineral N is significantly correlated to the TIN concentration in soil water at 30 cm depth (Fig. 7d). Thus, both internal N cycling and external N input determine inorganic N in soils available for leaching, also at these subtropical research sites at TSP and CJT.
As discussed previously, the in situ mineralization rate does not include initial decomposition of fresh litter. Thus, we also used an alternative definition for the soil flux density of mineral N*, being the sum of the annual net inorganic N input into the mineral soil as determined in the 0 cm lysimeter (Table 4), and the net N mineralization rate in the upper 15 cm of the mineral soil. This soil flux density of mineral N* in the upper 15 cm correlates significantly with the TIN concentration in soil water at 30 cm depth (Fig. 7e). These suggest that the flux density of mineral N (and flux density of mineral N*), which is the combination of deposition input and N status of system, can be used as indicator for N leaching in this system. However, the correlations are based on the TIN concentration and not fluxes at 30 cm soil depth. The N flux depends both on the concentration and the water flux, which may transport the N out the soil. So, the correlations only indicate the potential risk of leaching, the real leaching loss will also depend on the soil hydrological conditions.
Link between N status and leaching
The concentration and seasonality of leached NO −3 in stream water was taken as an indicator for the N saturation of forested catchments in Europe and North America. The early stages of N saturation are characterized by apparent seasonality and increases in the severity and frequency of NO −3 episodes in the growing season. The later stages of N saturation are marked by elevated concentrations of NO −3 even at base flow (Stoddard 1994; Dise et al. 1998b). For the conditions in Europe and North America, the forests are generally N limited and a tight N cycling in the ecosystem is proposed. N leaching loss in such ecosystems is supposed to be biologically controlled. However, the seasonality of stream water N concentration at TSP and CJT catchments from 2001 to 2003 show clear seasonality different from that for forested catchments in Europe and North America, with greatest concentrations commonly occurring in the wet growing season (from April through October). Lowest concentrations occur in the dry dormant season (from November through early April, Fig. 8). The concentration of N in stream water is correlated with discharge in these catchments, which is highest in the growing season (data not shown). This holds also true for episodic discharge in the dormant season of 2001–2002 (Fig. 8). The seasonal pattern is associated to the monsoon climate of wet and warm summers and dry and cold winters. Wet and warm summers favor N mineralization and nitrification in the soil, whereas dry and cold winter enhances the conservation of N in the soil. Similar observations of seasonality of NO −3 leaching, due to climatic and hydrological variations were made for forests in Japan with a monsoonal climate (Mitchell et al. 1997; Ohrui and Mitchell 1997). This may indicate that the seasonality of stream water NO −3 –N concentration in subtropical forested catchments cannot be interpreted with respect to N saturation using the same criteria as those used for boreal and temperate systems. The levels of annual N export differ significantly between TSP and CJT (Fig. 3). Levels of NO −3 –N concentration in stream water are higher at TSP than at CJT during base flow period (Fig. 8). This may suggest that the forest at TSP has a higher N status than the forest at CJT.

Seasonality of NO −3 –N in stream water in TSP and CJT catchment during January 2001 to December 2003. It also shows the wet and dormant season during the year
Conclusion
The N and C pools in subtropical Chinese forest soils are lower than those in temperate forest soils. This is primarily because the Chinese forest soils lack a continuous forest floor and a well-developed O horizon. In the upper 15 cm of the soil, the O/A horizon (0–3 cm) contributes about 25–55% to the total N pool and more than 30% to net N mineralization and nitrification. The C/N ratio of the O/A horizon cannot be used as indicator for N status and N leaching. However, the net N mineralization rate per unit organic C (mg N g−1 C year−1) in each of the O/A, B1, and B2 horizons is strongly correlated with the C/N ratio (an indication of substrate quality), indicating that the C/N ratio of mineral soil may be a promising indicator for N status of these subtropical forests. C/N values smaller than 22–24 are required before mineralization and nitrification take place.
Emission rates of N2O from Chinese subtropical forest soil correlate significantly with the total net nitrification rate (in mg N m−2 day−1) in the upper 15 cm of the soil. This suggests that N2O emission fluxes from the soil also can be used as an indicator for the N status of our forest stands. However, considerable spatial and temporal variation of N2O emissions from soil makes this parameter difficult to use.
Neither inorganic N deposition in throughfall, collected below the ground vegetation, nor the C/N ratio of the O/A soil horizon correlates with the concentration of inorganic N in soil water at 30 cm depth of soil. Thus these factors cannot be used as indicators for N leaching. The soil flux density of mineral N, which is the combination of deposition input and N status of the system, explains N leaching best. The seasonality of stream water N concentration, with highest values in summer, depends on climatic and hydrological conditions and differs from that found in boreal and temperate catchments.
References
Aber J, McDowell W, Nadelhoffer K, Magill A, Berntson G, Kamakea M, McNulty S, Currie W, Rustad L, Fernandez I (1998) Nitrogen saturation in temperate forest ecosystems—hypotheses revisited Bioscience 48:921–934
Aber JD, Goodale CL, Ollinger SV, Smith ML, Magill AH, Martin ME, Hallett RA, Stoddard JL (2003) Is nitrogen deposition altering the nitrogen status of northeastern forests? Bioscience 53:375–389
Ågren GI, Bosatta E (1988) Nitrogen saturation of terrestrial ecosystems. Environ Pollut 54:185–197
Andersson P, Berggren D, Nilsson I (2002) Indices for nitrogen status and nitrate leaching from Norway spruce (Picea abies (L.) Karst.) stands in Sweden. Forest Ecol Manage 157:39–53
Bengtsson G, Bengtsson P, Mansson KF (2003) Gross nitrogen mineralization-, immobilization-, and nitrification rates as a function of soil C/N ratio and microbial activity. Soil Biol Biochem 35:143–154
Bouwman AF, Fung I, Matthews E, John J (1993) Global analysis of the potential for N2O production in natural soils. Global Biogeochem Cycles 7:557–597
Breuer L, Papen H, Butterbach-Bahl K (2000) N2O emission from tropical forest soils of Australia. J Geophy Res-Atmos 105:26353–26367
Butterbach-Bahl K, Gasche R, Breuer L, Papen H (1997) Fluxes of NO and N2O from temperate forest soils: impact of forest type, N deposition and of liming on the NO and N2O emissions. Nutr Cycling Agroecosyst 1–2:79–90
Butterbach-Bahl K, Rothe, Papen H (2002) Effect of tree distance on N2O and CH4-fluxes from soils in temperate forest ecosystems. Plant Soil 240(1):91–103
Chen XY, Mulder J, Wang YH, Zhao DW, Xiang RJ (2004) Atmospheric deposition, minerlization and leaching of nitrogen in subtropical forested catchments, South China. Environ Geochem Health 26:179–186
Currie WS (1999) The responsive C and N biogeochemistry of the temperate forest floor. Trends Ecol Evol 14:316–320
Dise NB, Wright RF (1995) Nitrogen leaching from European forests in relation to nitrogen deposition. Forest Ecol Manag 71:153–161
Dise NB, Matzner E, Gundersen P (1998a) Synthesis of nitrogen pools and fluxes from European forest ecosystems. Water Air and Soil Pollut 105:143–154
Dise NB, Matzner E, Forsius M (1998b) Evaluation of organic horizon C:N ratio as an indicator of nitrate leaching in conifer forests across Europe. Environ Pollut 102:453–456
Emmett BA, Boxman D, Bredemeier M, Gundersen P, Kjonaas OJ, Moldan F, Schleppi P, Tietema A, Wright RF (1998) Predicting the effects of atmospheric nitrogen deposition in conifer stands: evidence from the NITREX ecosystem-scale experiments. Ecosystems 1:352–360
Fenn ME, Poth MA, Johnson DW (1996) Evidence for nitrogen saturation in the San Bernardino Mountains in southern California. Forest Ecol Manag 82:211–230
Fernandez IJ, Simmons JA, Briggs RD (2000) Indices of forest floor nitrogen status along a climate gradient in Maine, USA. Forest Ecol Manag 134:177–187
Galloway J, Cowling E, Kessler E (2002) Reactive nitrogen. Ambio 31:59
Gundersen P (1991) Nitrogen deposition and the forest nitrogen-cycle—role of denitrification. Forest Ecol Manag 44:15–28
Gundersen P, Emmett BA, Kjonaas OJ, Koopmans CJ, Tietema A (1998a) Impact of nitrogen deposition on nitrogen cycling in forests: a synthesis of NITREX data. Forest Ecol Manag 101:37–55
Gundersen P, Callesen I, de Vries W (1998b) Nitrate leaching in forest ecosystems is related to forest floor C/N ratios. Environ Pollut 102:403–407
Hall SJ, Matson PA (2003) Nutrient status of tropical rain forests influences soil N dynamics after N additions. Ecol Monogr 73:107–129
Hao JM, Tian HZ, Lu YQ (2002) Emission inventories of NOx from commercial energy consumption in China, 1995–1998. Environ Sci Technol 36:552–560
Larssen T, Tang D, He Y (eds) (2004) Integrated monitoring program on acidification of Chinese terrestrial systems—IMPACTS—Annual report results 2003. NIVA Report 4905-2004. NIVA, Oslo, Norway
MacDonald JA, Dise NB, Matzner E, Armbruster M, Gundersen P.and Forsius M (2002) Nitrogen input together with ecosystem nitrogen enrichment predict nitrate leaching from European forests. Global Change Biol 8:1028–1033
Maithani K, Arunachalam A, Tripathi RS, Pandey HN (1998) Nitrogen mineralization as influenced by climate, soil and vegetation in a subtropical humid forest in northeast India. Forest Ecol Manag 109:91–101
Matson PA, McDowell WH, Townsend AR, Vitousek PM (1999) The globalization of N deposition: ecosystem consequences in tropical environments. Biogeochemistry 46:67–83
Mitchell MJ, Iwatsubo G, Ohrui K, Nakagawa Y (1997) Nitrogen saturation in Japanese forests: an evaluation. Forest Ecol Manag 97:39–51
Ohrui K, Mitchell MJ (1997) Nitrogen saturation in Japanese forested watersheds. Ecol Appl 7:391–401
Ollinger SV, Smith ML, Martin ME, Hallett RA, Goodale CL, Aber JD (2002) Regional variation in foliar chemistry and N cycling among forests of diverse history and composition. Ecology 83:339–355
Oura N, Shindo J, Fumoto T, Toda H, Kawashima H (2001) Effects of nitrogen deposition on nitrous oxide emissions from the forest floor. Water Air Soil Pollut 130:673–678
Parsons DAB, Scholes MC, Scholes RJ, Levine JS (1996) Biogenic NO emissions from savanna soils as a function of fire regime, soil type, soil nitrogen, and water status. J Geophy Res-Atmos 101: 23683–23688
Post WM, Pastor J, Zinke PJ, Stangenberger AG (1985) Global patterns of soil-nitrogen storage. Nature 317:613–616
Schulze E-D (ed) (2000) Carbon and nitrogen cycling in European forest ecosystems. Ecological studies, vol. 142. Springer
Sitaula BK, Bakken LR, Abrahamsen G (1995) Nutrient balance in Scots pine (Pinus sylvestris L.) forest. 3. Fluxes of N2O from lysimeter as influenced by nitrogen input. Water Air Soil Pollut 85:1155–1159
Stoddard JL (1994) Long-term changes in watershed retention of nitrogen—its causes and aquatic consequences. Environ Chem Lakes Reservoirs 237:223–284
Theodose TA, Martin J (2003) Microclimate and substrate quality controls on nitrogen mineralization in a New England high salt marsh. Plant Ecol 167:213–221
Tietema A, Beier C, deVisser PHB, Emmett BA, Gundersen P, Kjonaas OJ, Koopmans CJ (1997) Nitrate leaching in coniferous forest ecosystems: the European field-scale manipulation experiments NITREX (nitrogen saturation experiments) and EXMAN (experimental manipulation of forest ecosystems). Global Biogeochem Cycles 11:617–626
Vitousek PM, Matson PA (1988) Nitrogen transformations in a range of tropical forest soils. Soil Biol Biochem 20:361–367
Wright RF, Beier C, Cosby BJ (1998) Effects of nitrogen deposition and climate change on nitrogen runoff at Norwegian boreal forest catchments: the MERLIN model applied to Risdalsheia (RAIN and CLIMEX projects). Hydrol Earth Syst Sci 2:399–414
Zhao DW, Wang AP (1994) Estimation of anthropogenic ammonia emissions in Asia. Atmos Environ 28:689–694
Zhu ZL, Chen DL (2002) Nitrogen fertilizer use in China—contributions to food production, impacts on the environment and best management strategies. Nutr Cycling Agroecosyst 63:117–127
Acknowledgments
We thank all partners of the Sino-Norwegian IMPACTS project, in particular project coordinators Tang Dagang (Chinese Research Academy of Environmental Sciences), and Espen Lydersen (Norwegian Institute for Water Research). Financial support for the IMPACTS was received from NORAD, the Norwegian Agency for Development Cooperation, from the State Environmental Protection Agency (SEPA) in Beijing, as well as from the provincial Environmental Protection Bureaus (Chongqing and Changsha for nitrogen study). The manuscript benefited greatly from two anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Chen, X.Y., Mulder, J. Indicators for nitrogen status and leaching in subtropical forest ecosystems, South China. Biogeochemistry 82, 165–180 (2007). https://doi.org/10.1007/s10533-006-9061-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-006-9061-3