
Abstract
Anaerobic bacteria have been used to produce 2,4-dihydroxylamino-nitrotoluene (2,4DHANT), a reductive metabolite of 2,4,6-trinitrotoluene (TNT). Here, an aerobic TNT biodegrader Pseudomonas sp. strain TM15 produced 2,4DHANT as evidenced by the molecular ion with m/z of 199 identified from LC-TOFMS analyses. TNT biodegradation with a high cell concentration (109 cells/ml) led to a significant accumulation of 2,4DHANT in the culture medium, as well as hydroxylamino-dinitrotoluenes (HADNTs), although these products were not accumulated when a low cell concentration was used; also, the accumulation of diamino-nitrotoluene and of an unidentified metabolite were observed in the culture medium with the high cell concentration (1010 cells/ml). 2,4DHANT overproduction was a function of the aeration speed since cultures with low aeration speeds (30 rpm) had a 19-fold higher DHANT productivity than those aerated with high speeds (180 rpm); this indicates that molecular oxygen was related to the formation of 2,4DHANT. The quantification of dissolved oxygen (DO) in the media demonstrated that the productivity of 2,4DHANT was increased at low DO values. Moreover, supplying oxygen to the culture media produced a remarkable decrease of 2,4DHANT accumulation; these results clearly indicate that high 2,4DHANT production was a consequence of the oxygen deficit in the culture medium. This finding is useful for understanding the TNT biodegradation (bioremediation technology) in an anoxic environment.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Highly explosive 2,4,6-trinitrotoluene (TNT) has been extensively used for military purposes, and its synthesis reached a maximum during World War II (Harter 1985). High concentrations of TNT and its analogs are still found in soil and groundwater at former manufacturing sites (Fernando et al. 1990; Hawari et al. 2000; Lewis et al. 2004; Maeda et al. 2006a). The persistence of these pollutants in the environment indicates that TNT and its analogs are recalcitrant to microbial biodegradation (Rieger and Knackmuss 1995) although some bacteria are chemotactic toward nitroaromatic compounds (Leungsakul et al. 2005). Also, TNT, its analogs, and its metabolites have strong cytotoxicity and mutagenicity for various organisms (Ahlborg et al. 1988; Berthe-Corti et al. 1998; Letzel et al. 2003; Padda et al. 2003; Saka 2004; Tan et al. 1992; Won et al. 1976); thereby, developing bioremediation technologies for TNT and its analogs is significant.
The TNT biotransformation pathway by aerobic or anaerobic bacteria has been described (Esteve-Nunez et al. 2001; Hawari et al. 2000; Snellinx et al. 2002); commonly, TNT is biotransformed by converting nitro groups into nitroso, hydroxylamino and amino groups. Thus, because of its high electron deficiency, initial microbial transformations of TNT are characterized by reductive rather than by oxidative reactions (Barrows et al. 1996). In several bacterial species, the major reduction metabolites from TNT are hydroxylamino-dinitrotoluenes (HADNTs) and amino-dinitrotoluenes (ADNTs) that are formed by reducing one of three nitro groups (Ahmad and Hughes 2002; Borch et al. 2005; Fleischmann et al. 2004; Huang et al. 2000; Kroger et al. 2004; Vorbeck et al. 1998; Yin et al. 2005). Also dihydroxylamino-nitrotoluene (DHANT) is accumulated in some anaerobic bacteria during TNT metabolism (Ahmad and Hughes 2002; Borch et al. 2005; Fleischmann et al. 2004; Lewis et al. 1996; Yin et al. 2005); aerobically, DHANT is only produced by Pseudomonas pseudoalcalogenes JS52 (Fiorella and Spain 1997), which is a spontaneous mutant derived from strain JS45 (Nishino and Spain 1993). P. pseudoalcalogenes produces DHANT by the conversion of two nitro groups into two hydroxylamino groups. Why different reduction metabolites are produced from the diverse TNT-metabolizing bacteria performing the same TNT reduction reaction is unknown; therefore, an understanding of the mechanism of the TNT reduction reaction is significant for developing TNT bioremediation technologies since the initial step for TNT biodegradation is a reduction reaction.
Pseudomonas sp. strain TM15 was isolated from the TNT-contaminated soil in the Yamada Green Zone, Kitakyushu, Japan (Maeda et al. 2006a). This strain aerobically produces HADNTs and ADNTs from TNT, as previously described (Maeda et al. 2006a, c), and it was found that HADNTs accumulate in the culture medium using a high cell density (108 cells/ml) in comparison with low cell densities (107 cells/ml) (Maeda et al. 2006b). This implies that TNT degradation at high cell density may trigger different TNT-biodegradation patterns. This phenomenon is important for the TNT bioremediation because it requires specific bacteria with high activity for TNT degradation and enables more rapid decontamination of TNT-polluted sites. The goal of this paper is to examine the accumulation pattern of TNT reduction metabolites in various conditions such as different cell numbers, shaking speeds and oxygen concentrations.
Materials and methods
Chemicals
2,4,6-Trinitrotoluene (TNT) was obtained from Chugoku Kayaku Co. Ltd., Hiroshima, Japan and 2-hydroxylamino-4,6-dinitrotoluene (2HADNT), 4-hydroxylamino-2,6-dinitrotolulene (4HADNT), 2,4-diamino-nitrotoluene (2,4DANT) were kindly provided by Dr. R. Spanggord (Chemical Sciences and Technology Department, SRI International, Melno Park, CA). 2-Amino-4,6-dinitrotoluene (2ADNT), and 4-amino-2,6-dinitrotolulene (4ADNT) were purchased from AccuStandard, Inc., New Haven, USA. All chemicals used were of the highest purity commercially available.
Bacterial strains, growth and total protein determination
Pseudomonas sp. strain TM15 was isolated from the soil in the Yamada Green Zone, Kitakyushu, Japan (Maeda et al. 2006a). Cells were initially streaked from −80°C glycerol stocks on Luria-Bertani (LB) agar plates (Sambrook et al. 1989) containing 100 mg/l TNT and incubated at 30°C. After growth on the LB agar plates, Pseudomonas sp. strain TM15 was cultured from a fresh single colony in LB medium (Sambrook et al. 1989) supplemented with 100 mg/l TNT at 30°C in the dark with shaking at 120 rpm (MM-10, TAITEC Co. Ltd., Saitama, Japan). Then, the cells were harvested by centrifugation at 5000 × g for 5 min at 4°C, washed twice in 50 mM autoclaved phosphate buffer (pH 7.0), and resuspended in the same phosphate buffer. The cell suspensions were used for TNT biodegradation experiments. Cell growth was measured using turbidity at 600 nm with a UV/vis Spectrophotometer (V-530, JASCO Co. Ltd., Tokyo, Japan), and total protein for Pseudomonas sp. TM15 was determined by the following formula: 0.20 mg/OD/ml.
TNT biodegradation
The cells (106–1010 cells/ml) were inoculated in M8 minimal medium (200 ml), which is M9 minimal medium without NH4Cl (Sambrook et al. 1989), including acetate (10 mM) and 100 mg/l TNT as carbon and nitrogen sources. The mixtures were aerobically incubated at 30°C in the dark with shaking (30, 60, 120 or 180 rpm; MM-10, TAITEC Co. Ltd.).
Measurement of TNT metabolites
Cells were removed by centrifugation at 5000 × g for 20 s, and then the culture fluid was filtered with membrane filters (0.2 μm, Toyo Roshi Kaisha Ltd., Tokyo, Japan). TNT and its metabolites (2HADNT, 4HADNT, 2ADNT, 4ADNT, DANT, and unidentified metabolite (2,4DHANT)) were determined by high-performance liquid chromatography (HPLC; Shimadzu Co. Ltd., Kyoto, Japan) without sample concentration. HPLC measurements were performed on an Inertsil ODS-2 column (4.0 mm × 150 mm; GL Sciences Inc., Tokyo, Japan) with acetonitrile-water (40:60) as the mobile phase, with a flow rate of 0.4 ml/min. All compounds were detected at 254 nm with a Shimadzu SPD-10AVP UV–vis detector. Productivity for 2,4DHANT was indicated as peak area/h/mg-protein from the 2,4DHANT proportional production time because standard chemical of 2,4DHANT was not available (i.e. it was impossible to determine the concentration for 2,4DHANT).
Extraction, thin-layer chromatography, and detection
The culture fluid (200 ml) was extracted twice with 50 ml of dichloromethane and/or diethyl ether, then the extracts were dried over anhydrous sodium sulfate, and excess solvent was removed by rotary evaporation (R-114-EW-3, Sibata Scientific Technology Ltd., Tokyo, Japan). Thin-layer chromatography (TLC) was performed to analyze the TNT metabolites extracted in organic solvents. The samples (20 μl) were spotted on a TLC sheet (10 cm × 20 cm; Silica gel 60 TLC aluminum sheet, Merck KGaA, Darmatadt, Frankfurt, Germany), and subsequently developed once by the ascending method with a solvent system consisting of ethanol–benzene–hexane (1:2:2 by volume) for a distance of 16 cm (total time 2 h). Spots indicating TNT metabolites were visualized at 254 nm under UV illumination (LPR-33/JM, TAITEC Co. Ltd.).
Purification of metabolites and LC/TOF–MS instrumentation
To purify 2,4DHANT from various TNT metabolites, the target compounds were extracted into acetonitrile by collecting the spots. Purity of the purified metabolite was immediately assayed by HPLC. To obtain the mass spectra of this compound, the samples were measured by liquid chromatography/time-of-flight mass spectrometer (LC/TOF–MS; Agilent 6210 LC/MS time-of-flight mass spectrometer, Agilent Technologies, Santa Clara, CA, USA). LC/TOF–MS measurements were performed on a ZORBAX SB-Aq column (4.6 mm × 150 mm; Agilent Technologies Japan, Ltd., Tokyo, Japan) with acetonitrile–water (the gradient program was 0% acetonitrile for 5 min at 0.4 ml/min, and then 2.5% acetonitrile/min to reach up to 50% (for 20 min)) as the mobile phase, with a flow rate of 0.4 ml/min. Ionization to obtain mass spectrum was the ESI system with 150 V and positive mode.
Calculation of composition formula
The LC/TOF–MS system allows accurate mass analyses in the molecular ion peak and fragment ion peak; hence, composition formula can be obtained from the accurate mass in the peak. The instrument software constantly corrects the measured masses of all the spectra using the known masses as reference. LC/MS accurate mass spectra were recorded across the range m/z 90–1100. The full-scan data recorded was processed with Applied Biosystems/MDS-SCIEX Analyst QS software (Frankfurt, Germany) with accurate mass application-specific additions from Agilent MS TOF software.
Measurement of dissolved oxygen
Dissolved oxygen (DO) in samples (50 ml) was measured by using DO meter (DO-14P, TOA electronics Ltd., Tokyo, Japan).
Sparging with oxygen gas
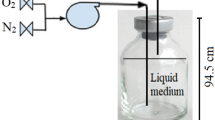
To examine the effect of oxygen on the production of DHANT, oxygen gas (Itochu Industrial Gas Co. Ltd., Tokyo, Japan) was supplied into the TNT culture medium at a flow rate of 50 ml/min while TNT was degraded by Pseudomonas sp. strain TM15. TNT and its metabolites were analyzed by HPLC as described above.
Results
Accumulation of TNT metabolites from Pseudomonas sp. strain TM15
Pseudomonas sp. strain TM15 produced 2ADNT and 4ADNT from TNT as previously described (Maeda et al. 2006a). Also, TNT biodegradation using higher cell densities (more than 109 cells/ml) showed that Pseudomonas sp. strain TM15 accumulates one unidentified metabolite which has a 4.5 min retention time with HPLC (Fig. 1a), as well as accumulates HADNTs up to 9-fold at high cell density (over 108 cells/ml) compared to the low cell density (107 cells/ml) (Maeda et al. 2006b). As shown in Fig. 1b, TNT completely disappeared after a 2-h cultivation with 109 cfu/ml. On the other hand, the concentration of HADNTs reached a maximum during the 2-h cultivation then gradually decreased. Also, the unidentified metabolite reached a maximum after 6 h with Pseudomonas sp. TM15, and the amount of this metabolite was constant for 6- to 10-h cultivation (Fig. 1b).

Accumulation of TNT reduction metabolites by Pseudomonas sp. strain TM15. (a) HPLC chromatogram of 2,4,6-trinitrotoluene (TNT), hydroxylamino-dinitrotoluenes (HADNTs) and an unidentified metabolite (retention time: 4.5 min). (b) Time course of TNT (●), HADNTs (▴) and an unidentified metabolite (RT 4.5) (■) during TNT biodegradation. The cells (109 cfu/ml) were inoculated into M8 minimal medium including TNT (100 mg/l) and then aerobically incubated at 30°C with shaking (120 rpm)
Identification of the unknown metabolite
TNT metabolites were extracted into dichloromethane and/or diethyl ether in order to isolate the unknown product with the 4.5 min retention time; this enabled us to identify this metabolite by using infrared resonance spectroscopy and LC–MS. This unknown metabolite was not extracted with dichloromethane whereas it was extracted with diethyl ether as shown in Fig. 2, while other metabolites were extracted with both organic solvents. Therefore, a two-step extraction was performed for isolating the unknown product; first an extraction with dichloromethane was performed in order to extract other metabolites except the unidentified metabolite, and then a second extraction step was done with diethyl ether in order to extract the remaining metabolites; thereby, the unknown metabolite was found in diethyl ether and not in dichloromethane. As expected, this two-step extraction clearly showed that the unidentified product could be extracted without contamination (Fig. 2). The unknown metabolite was purified by collecting the spots on TLC plate and by extracting into acetonitrile (Supplementary Fig. 1), and then the purity was measured by HPLC. HPLC analyses indicated that purity of the unknown product was 99.5 ± 1.0%. Then, this purified sample was measured by IR analysis (Supplementary Fig. 2) and LC/MS analysis (Supplementary Fig. 3). Furthermore, LC/TOF–MS analysis was conducted to reveal what the unidentified metabolite is. The mass spectrum (conducted by using a high voltage (150 V)) showed a m/z 200 ([M+H]+) for the molecular ion and the typical M-17 ion loss (m/z 183), which is specific for nitro groups ortho in nitroaromatics harboring a methyl group (Fig. 3). Additionally, accurate mass analyses showed the m/z was 200.0661 for the molecular ion, revealing that the composition of the unknown was C7H10N3O4, which is identical to that of 2,4DHANT. Also, the fragment ion peak (m/z 183.0632 obtained by the 17 ion loss) was 183.0632, indicating C7H9N3O3 as the composition; hence, judging from the result of LC/TOF–MS spectra, we propose this unknown metabolite is 2,4DHANT. The results were absolutely confirmed by Dr. Robin Gerlach (Montana State University, Bozeman, MT), who had a low amount of 2,4DHANT and who analyzed our purified sample to reveal whether our unknown product was 2,4DHANT (Supplementary Fig. 4). Hence, we identified the undetermined metabolite showing a 4.5 min of retention time was without a doubt 2,4DHANT for the subsequent experiments.

Thin-layer chromatogram of standard chemicals (lanes 1–3) and samples (lanes 4–6). TLC was utilized with the ascending method with a solvent system consisting of ethanol–benzene–hexane (1:2:2 by volume) for a distance of 16 cm (total time 2 h). Standard chemicals were TNT (lane 1), 2HADNT (lane 2) and 4HADNT (lane 3). TNT metabolites produced by Pseudomonas sp. strain TM15 were extracted by dichloromethane (lane 4) or diethyl ether (lane 5) or both solvents (lane 6) from the TNT reaction solution after 6-h cultivation at 30°C with shaking (120 rpm)

HPLC chromatogram (a) and mass spectrum (b) for 2,4DHANT. Samples were measured by using LC/TOF–MS analyses
Accumulation pattern of DHANT according to cell number increase
To reveal if the accumulation pattern of 2,4DHANT depended on cell number, the amount of TNT and the metabolites in the culture medium at various cell densities (105, 106, 107, 108, 5 × 108, 109, and 1010 cfu/ml) were measured. Increasing cells in the culture certainly elevated the TNT biotransformation rate (Table 1). TNT biodegradation with high cell densities (more than 109 cells/ml) significantly accumulated 2,4DHANT after 6- to 10-h cultivation and after 0.5- to 1-h cultivation in the cultures containing 109 and 1010 cfu/ml of cells, respectively (Fig. 4). In addition, 2,4DHANT produced from 1010 cfu/ml cells gradually decreased with time and then 2,4DANT accumulated in the culture medium, although it was not observed in the condition with 109 cfu/ml cells.

Metabolic dynamics of TNT and its metabolites (TNT (●), HADNTs (▴), 2,4DHANT (■) and 2,4DANT (♦)) during TNT biodegradation using cells at 109 cfu/ml (a) and 1010 cfu/ml (B) with shaking at 120 rpm
Effect of aeration speed in the production of DHANT
To further investigate the accumulation pattern of 2,4DHANT, TNT and its metabolites were measured using cultures with different aeration speeds (30, 60, 120, and 180 rpm). TNT biodegradation rates were almost the same with 30, 60, 120 and 180 rpm (Table 2); however, production of HADNTs and 2,4DHANT showed a different accumulation pattern for each aeration speed. Notably, TNT biodegradation at low aeration speeds (30 and 60 rpm) increased the accumulation of 2,4DHANT as shown in Fig. 5 with the maximum amount of the 2,4DHANT accumulated at 30 rpm (11.4 times higher than that with an aeration speed of 180 rpm) (Table 2). Similarly 2,4DHANT productivity remarkably decreased as aeration speed increased, indicating that oxygen may relate to the accumulation of 2,4DHANT. On the other hand, TNT degradation with low aeration (30 rpm) and high cell density (109 cells/ml) formed 2,4DANT after the decrease of 2,4DHANT at 2 h (Fig. 6). Furthermore, a new unidentified metabolite which has 5.5 min of retention time was detected after the 2,4DANT decrease after 6 h. The accumulation of this unknown metabolite (RT 5.5) was not observed for the culture incubated at 120 or 180 rpm. Taking together, our results showed that high cell density and low aeration speed during TNT biodegradation leads to a high accumulation of 2,4DHANT.

Productivity of 2,4DHANT by Pseudomonas sp. strain TM15 (5 × 108 cfu/ml) with various aeration speeds (30 rpm, ●; 60 rpm, ○; 120 rpm, ▴ 180 rpm, Δ)

Metabolic dynamics of TNT and its metabolites (TNT (●), HADNTs (○), 2,4DHANT (▴), 2,4DANT (Δ), and unidentified metabolite (RT 5.5, ■) in TNT biodegradation using cells at 109 cfu/ml with shaking (30 rpm)
Effect of oxygen in production of 2,4DHANT
To reveal the mechanism for forming 2,4DHANT, the effect of oxygen in TNT biotransformation was tested. First, we measured dissolved oxygen (DO) in the culture medium with the different conditions (Table 3). After 10-h of cultivation, the culture media of the bacteria with high aeration speeds (120 and 180 rpm) clearly showed higher DO values than those with slow aeration (30 and 60 rpm); hence, this experiment showed that DO in the culture medium is responsible for high productivity of 2,4DHANT. Next, to directly reveal the effect of oxygen in producing 2,4DHANT, we ascertained production of 2,4DHANT while sparging the culture medium with oxygen gas during TNT biodegradation, and found the supply of oxygen dramatically decreased the accumulation of 2,4DHANT (Fig. 7). These two experiments showed clearly that the molecular oxygen concentration was related to the high production of 2,4DHANT.

Effect of oxygen supply in metabolic dynamics of TNT and its metabolite (TNT (●, ○), HADNTs (▴, Δ) and 2,4DHANT (■, □)) in TNT biodegradation by Pseudomonas sp. strain TM15 (cells: 109 cfu/ml and shaking speed: 60 rpm). The TNT solutions were sparged with (open) or without (solid) pure oxygen gas as described in Section “Materials and methods”
Discussion
The persistence of TNT and its metabolites is an environmental concern because they may be cytotoxic and genotoxic to many living organisms (Ahlborg et al. 1988; Berthe-Corti et al. 1998; Letzel et al. 2003; Sun et al. 2005); therefore, it is imperative to construct an effective remediation technology for TNT as soon as possible. Recently, it has been reported that TNT treatment in strictly anaerobic bioreactors results in tight binding of TNT reduction products to soil organic matter and that TNT mineralization is due to an aerobic treatment rather than the anaerobic TNT degradation (Newcombe and Crawford 2007); hence, TNT degradation by aerobic bacteria hold promise for mineralizing TNT. In addition, discoveries of bacteria possessing a high potential for TNT degradation and the use of high cell concentrations that enable rapid degradation of TNT will be necessary for constructing a complete TNT-degradation. Based on this concept, aerobic TNT degradation using a high cell density was conducted herein, and the results show that HADNTs and 2,4DHANT accumulate significantly in the culture medium and the accumulation is due to low levels of oxygen. Hence, our results are helpful for developing TNT-degradation technologies that can mineralize TNT completely since we have shown here the possibility that an anoxic environment is generated by consuming molecular oxygen by bacteria during aerobic degradation.
To date, many TNT-metabolizing bacteria have been reported (Esteve-Nunez et al. 2001; Hawari et al. 2000; Heiss and Knackmuss 2002; Snellinx et al. 2002); usually aerobic and anaerobic bacteria metabolize TNT via reductive pathways, producing hydroxylamino-, amino-compounds or a combination of both substituted compounds. The impossibility of an oxidative attack, which occur with other nitroaromatics such as mono- and dinitrotoluenes, stems from the extremely small electron density on the aromatic ring of TNT because three symmetrically arranged nitro groups of TNT cover the aromatic ring (creating steric hindrance), and more important, reduce the electron density of the ring by resonance (Barrows et al. 1996). Therefore, most organisms produce HADNTs, ADNTs and DANT as common metabolites. Under strictly anaerobic conditions, further reduction of TNT to triaminotoluene is possible (Esteve-Nunez et al. 2001; Hawari et al. 2000). On the other hand, some TNT-metabolizing anaerobes (Ahmad and Hughes 2002; Borch et al. 2005; Fleischmann et al. 2004; Huang et al. 2000; Lewis et al. 1996) and one aerobic bacterium (Pseudomonas pseudoalcaligenes JS52) (Fiorella and Spain 1997) are able to convert two of three nitro groups into hydroxylamino-substituted compounds, resulting in the production of 2,4DHANT. Nitroreductase, which catalyzes the conversion of nitro groups into nitroso, hydroxylamino and amino groups, is responsible for producing a variety of reductive TNT metabolites (Hannink et al. 2001); however, why different reduction metabolites are produced during TNT biotransformation by various bacteria is unclear.
Our results show that TNT biodegradation with a high cell density of Pseudomonas sp. strain TM15 can provide rapid degradation of TNT (Table 1). Also, TNT metabolism with a high cell density and with low aeration speeds produced high amounts of HADNTs and 2,4DHANT in the culture medium (Tables 2 and 3), as well as ADNTs that is a mutagen (Banerjee et al. 2003; Tan et al. 1992; Won et al. 1976). HADNTs are strong mutagens (Maeda et al. 2006a; Padda et al. 2003); it has been reported that these compounds cause oxidative DNA damage (formation of 8-oxo-7,8-dihydro-2′-deoxyguanosine) in the presence of NADH and Cu(II) (Homma-Takeda et al. 2002), although these chemicals have no cytotoxicity (Maeda et al. 2006a). HADNTs may be toxic to living organisms because they may induce many mutations for DNA without themselves being directly cytotoxic. On the other hand, 2,4DHANT has higher mutagenicity than HADNTs (Padda et al. 2003), although it is unknown whether this compound is cytotoxic; hence, we will have to consider this while developing effective TNT bioremediation technologies. Our results are significant for handling this problem and may contribute to the development of TNT-biodegradation technology.
TNT biodegradation with a high cell density and with low aeration speeds showed high accumulation of HADNTs and 2,4DHANT and these accumulations are responsible for decreasing molecular oxygen in the culture medium (Fig. 7); these conditions will create an anoxic environment in the culture medium through bacterial oxygen consumption. Also, supplying oxygen gas to the culture medium remarkably repressed the accumulation of 2,4DHANT whereas TNT degradation rates were the same under both conditions; this means that the presence of oxygen affects the reductive metabolism from TNT. The accumulations of HADNTs, ADNTs and 2,4DANT were almost the same between the oxic and anoxic conditions; hence, the oxygen supply may promote the subsequent metabolism of 2,4DHANT or may lead to other metabolic pathways (e.g., oxidation pathway; production of 2,4,6-trinitrobenzoic acid as well as benzoic acid production from toluene) (Chen et al. 2005; Liou et al. 2004). We will have to investigate whether TNT metabolism under aerobic or anaerobic conditions proceed with the same or different pathways, although our latest results showed the same pathway between these conditions; hence, we need to further characterize other metabolites produced during the TNT metabolism. On the other hand, since HADNTs are spontaneously converted into tetranitroazoxytoluene in the presence of molecular oxygen (Haidour and Ramos 1996; Maeda et al. 2007a); 2,4DHANT may be converted into azoxytoluene-related compounds through the same pathway. However, to date, such metabolites from 2,4DHANT have been not found. It has been reported that DHANT was stable in anoxic aqueous solutions, and in the presence of oxygen, was converted to yellow polar metabolites having absorbance maxima at 258, 325, and 405 nm (Fiorella and Spain 1997). Such metabolites having three absorbance maxima were not detected from the culture medium sparged with oxygen although the accumulation of 2,4DHANT was decreased by supplying molecular oxygen. Hence, our results here are not the same as the previous study and it is proposed that the different TNT degradation will happen in oxic or anoxic conditions.
We hypothesize that the production of significant amounts of 2,4DHANT by Pseudomonas sp. strain TM15 in an anoxic environment may be due to non-specific electron transfer to TNT; namely, it suggests that the electrons created from the respiratory chain that usually react with molecular oxygen in oxic conditions (which has −80 mV of a single reduction potential compared to −565 mV for TNT (Maeda et al. 2007b)); in the anoxic environment created through the bacterial oxygen consumption, the electrons could attack TNT instead of oxygen (it may be irrelevant for the reduction reaction by nitroreductase enzymes), enhancing the 2,4DHANT production. Actually, in situ bioremediation may create anoxic environments by using a high bacterial concentration to degrade chemical pollutants (i.e., low oxygen concentration generated by bacterial oxygen consumption); hence, our finding that showed the different TNT degradation patterns in anoxic environment provides substantial information to develop bioremediation technologies for various chemical pollutants.
Conclusions
TNT biodegradation with high cell densities or with low aeration speeds led to a significant accumulation of 2,4DHANT using the aerobic TNT degrader Pseudomonas sp. strain TM15. The accumulation is clearly responsible for molecular oxygen; the presence or absence of oxygen in culture medium showed differences in the accumulation pattern of TNT reductive metabolites such as HADNTs, 2,4DHANT, and 2,4DANT. Our finding contributes to an understanding of TNT metabolism in anoxic environments that is created through oxygen consumption by TNT-metabolizing bacteria.
References
Ahlborg G Jr, Einisto P, Sorsa M (1988) Mutagenic activity and metabolites in the urine of workers exposed to trinitrotoluene (TNT). Br J Ind Med 45:353–358
Ahmad F, Hughes JB (2002) Reactivity of partially reduced arylhydroxylamine and nitrosoarene metabolites of 2,4,6-trinitrotoluene (TNT) toward biomass and humic acids. Environ Sci Technol 36:4370–4381
Banerjee H, Hawkins Z, Dutta S, Smoot D (2003) Effects of 2-amino-4,6-dinitrotoluene on p53 tumor suppressor gene expression. Mol Cell Biochem 252:387–389
Barrows SE, Cramer CJ, Truhlar DG, Elovitz MS, Weber EJ (1996) Factors controlling regioselectivity in the reduction of polynitroaromatics in aqueous solution. Environ Sci Technol 30:3028–3038
Berthe-Corti L, Jacobi H, Kleihauer S, Witte I (1998) Cytotoxicity and mutagenicity of a 2,4,6-trinitrotoluene (TNT) and hexogen contaminated soil in S. typhimurium and mammalian cells. Chemosphere 37:209–218
Borch T, Inskeep WP, Harwood JA, Gerlach R (2005) Impact of ferrihydrite and anthraquinone-2,6-disulfonate on the reductive transformation of 2,4,6-trinitrotoluene by a gram-positive fermenting bacterium. Environ Sci Technol 39:7126–7133
Chen WS, Juan CN, Wei KM (2005) Mineralization of dinitrotoluenes and trinitrotoluene of spent acid in toluene nitration process by Fenton oxidation. Chemosphere 60:1072–1079
Esteve-Nunez A, Caballero A, Ramos JL (2001) Biological degradation of 2,4,6-trinitrotoluene. Microbiol Mol Biol Rev 65:335–352, table of contents
Fernando T, Bumpus JA, Aust SD (1990) Biodegradation of TNT (2,4,6-trinitrotoluene) by Phanerochaete chrysosporium. Appl Environ Microbiol 56:1666–1671
Fiorella PD, Spain JC (1997) Transformation of 2,4,6-trinitrotoluene by Pseudomonas pseudoalcaligenes JS52. Appl Environ Microbiol 63:2007–2015
Fleischmann TJ, Walker KC, Spain JC, Hughes JB, Morrie Craig A (2004) Anaerobic transformation of 2,4,6-TNT by bovine ruminal microbes. Biochem Biophys Res Commun 314:957–963
Haidour A, Ramos JL (1996) Identification of products resulting from the biological reduction of 2,4,6-trinitrotoluene, 2,4-dinitrotoluene, and 2,6-dinitrotoluene by Pseudomonas sp. Environ Sci Technol 30:2365–2370
Hannink N, Rosser SJ, French CE, Basran A, Murray JA, Nicklin S, Bruce NC (2001) Phytodetoxification of TNT by transgenic plants expressing a bacterial nitroreductase. Nat Biotechnol 19:1168–1172
Harter DR (1985) The use and importance of nitroaromatic compounds in the chemical industry. In: Rickert DE (ed). Hemisphere publishing, Washington, DC
Hawari J, Beaudet S, Halasz A, Thiboutot S, Ampleman G (2000) Microbial degradation of explosives: biotransformation versus mineralization. Appl Microbiol Biotechnol 54:605–618
Heiss G, Knackmuss HJ (2002) Bioelimination of trinitroaromatic compounds: immobilization versus mineralization. Curr Opin Microbiol 5:282–287
Homma-Takeda S, Hiraku Y, Ohkuma Y, Oikawa S, Murata M, Ogawa K, Iwamuro T, Li S, Sun GF, Kumagai Y, Shimojo N, Kawanishi S (2002) 2,4,6-trinitrotoluene-induced reproductive toxicity via oxidative DNA damage by its metabolite. Free Radic Res 36:555–566
Huang S, Lindahl PA, Wang C, Bennett GN, Rudolph FB, Hughes JB (2000) 2,4,6-trinitrotoluene reduction by carbon monoxide dehydrogenase from Clostridium thermoaceticum. Appl Environ Microbiol 66:1474–1478
Kroger M, Schumacher ME, Risse H, Fels G (2004) Biological reduction of TNT as part of a combined biological-chemical procedure for mineralization. Biodegradation 15:241–248
Letzel S, Goen T, Bader M, Angerer J, Kraus T (2003) Exposure to nitroaromatic explosives and health effects during disposal of military waste. Occup Environ Med 60:483–488
Leungsakul T, Keenan BG, Smets BF, Wood TK (2005) TNT and nitroaromatic compounds are chemoattractants for Burkholderia cepacia R34 and Burkholderia sp. strain DNT. Appl Microbiol Biotechnol 69:321–325
Lewis TA, Goszczynski S, Crawford RL, Korus RA, Admassu W (1996) Products of anaerobic 2,4,6-trinitrotoluene (TNT) transformation by Clostridium bifermentans. Appl Environ Microbiol 62:4669–4674
Lewis TA, Newcombe DA, Crawford RL (2004) Bioremediation of soils contaminated with explosives. J Environ Manage 70:291–307
Liou MJ, Lu MC, Chen JN (2004) Oxidation of TNT by photo-Fenton process. Chemosphere 57:1107–1114
Maeda T, Kadokami K, Ogawa HI (2006a) Characterization of 2,4,6-trinitrotoluene (TNT)-metabolizing bacteria isolated from TNT-polluted soils in the Yamada Green Zone, Kitakyushu, Japan. J Environ Biotechnol 6:33–39
Maeda T, Kubota A, Nagafuchi N, Kadokami K, Ogawa HI (2006b) Crucial problem in rapid spectrophotometric determination of 2,4,6-trinitrotoluene and its breakthrough method. J Microbiol Methods 66:568–571
Maeda T, Nagafuchi N, Kubota A, Kadokami K, Ogawa HI (2006c) One-step isolation and identification of hydroxylamino-dinitrotoluenes, unstable products from 2,4,6-trinitrotoluene metabolites, with thin-layer chromatography and laser time-of-flight mass spectrometry. J Chromatogr Sci 44:96–100
Maeda T, Nagafuchi N, Kubota A, Kadokami K, Ogawa HI (2007a) Identification of spontaneous conversion products of unstable 2,4,6-trinitrotoluene metabolites, hydroxylamino-dinitrotoluenes, by combination of thin-layer chromatography and laser time-of-flight mass spectrometry. J Chromatogr Sci 45:345–349
Maeda T, Nakamura R, Kadokami K, Ogawa HI (2007b) Relationship between mutagenicity and reactivity or biodegradability for nitroaromatic compounds. Environ Toxicol Chem 26:237–241
Newcombe DA, Crawford RL (2007) Transformation and fate of 2,4,6-trinitrotoluene (TNT) in anaerobic bioslurry reactors under various aeration schemes: implications for the decontamination of soils. Biodegradation 18:741–754
Nishino SF, Spain JC (1993) Degradation of nitrobenzene by a Pseudomonas pseudoalcaligenes. Appl Environ Microbiol 59:2520–2525
Padda RS, Wang C, Hughes JB, Kutty R, Bennett GN (2003) Mutagenicity of nitroaromatic degradation compounds. Environ Toxicol Chem 22:2293–2297
Rieger P-G, Knackmuss H-J (1995) Basic knowledge and perspectives on biodegradation of 2,4,6-trinitrotoluene and related nitroaromatic compounds in contaminated soil. In: Spain JC (ed). Plenum Press, New York, NY
Saka M (2004) Developmental toxicity of p,p′-dichlorodiphenyltrichloroethane, 2,4,6-trinitrotoluene, their metabolites, and benzo[a]pyrene in Xenopus laevis embryos. Environ Toxicol Chem 23:1065–1073
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular Cloning, A Laboratory Manual, 2nd edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Snellinx Z, Nepovim A, Taghavi S, Vangronsveld J, Vanek T, van der Lelie D (2002) Biological remediation of explosives and related nitroaromatic compounds. Environ Sci Pollut Res Int 9:48–61
Sun Y, Iemitsu M, Shimojo N, Miyauchi T, Amamiya M, Sumi D, Hayashi T, Sun G, Shimojo N, Kumagai Y (2005) 2,4,6-Trinitrotoluene inhibits endothelial nitric oxide synthase activity and elevates blood pressure in rats. Arch Toxicol 79:705–710
Tan EL, Ho CH, Griest WH, Tyndall RL (1992) Mutagenicity of trinitrotoluene and its metabolites formed during composting. J Toxicol Environ Health 36:165–175
Vorbeck C, Lenke H, Fischer P, Spain JC, Knackmuss HJ (1998) Initial reductive reactions in aerobic microbial metabolism of 2,4,6-trinitrotoluene. Appl Environ Microbiol 64:246–252
Won WD, DiSalvo LH, Ng J (1976) Toxicity and mutagenicity of 2,4,-6-trinitrotoluene and its microbial metabolites. Appl Environ Microbiol 31:576–580
Yin H, Wood TK, Smets BF (2005) Reductive transformation of TNT by Escherichia coli: pathway description. Appl Microbiol Biotechnol 67:397–404
Acknowledgments
We are grateful to Chugoku Kayaku Co. Ltd., Hiroshima, Japan, for their gift of 2,4,6-trinitrotoluene, and to Dr. R. Spanggord, Chemical Sciences and Technology Department, SRI International, Melno Park, CA, for his gift of 2-hydroxylamino-4,6-dinitrotoluene, 4-hydroxylamino-2,6-dinitrotolulene, and 2,4-diamino-nitrotoluene. We express much gratitude to Dr. Robin Gerlach, Montana State University, Bozeman, MT, for his help in identifying 2,4-dihydroxylamino-6-nitrotoluene, to Dr. Hideaki Uchida and Dr. Yoshizumi Takigawa, Agilent Technologies Japan, Ltd., for their support with LC/TOF–MS, and to Dr. Norikazu Nishino, Department of Biological Functions and Engineering, Graduate School of Life Science and Systems Engineering, Kyushu Institute of Technology, for his support with the LC/MS analyses. We are also grateful for the assistance of Dr. Thomas K. Wood of Texas A & M University, College Station, TX, for his kind helps with preparing this manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Kubota, A., Maeda, T., Nagafuchi, N. et al. TNT biodegradation and production of dihydroxylamino-nitrotoluene by aerobic TNT degrader Pseudomonas sp. strain TM15 in an anoxic environment. Biodegradation 19, 795–805 (2008). https://doi.org/10.1007/s10532-008-9182-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10532-008-9182-6