
Abstract
Studied was the effect of temperature in the range 12–46 °C on the rate of bacterial decolorization of the mono-azo dye Acid Orange 7 by Alcaligenes faecalis 6132 and Rhodococcus erythropolis 24. With both strains the raise of temperature led to a corresponding raise of decolorization rate better manifested by R. erythropolis. The analysis of the Arrhenius plot revealed a break near the middle of the temperature range. The regression analysis showed practically complete identity of the observed break point temperatures (T BP): 20.7 °C for Alc. faecalis and 20.8 °C for R. erythropolis. The values of the activation energy of the decolorization reaction (E a) were found to depend on both the organism and the temperature range. In the range below T BP the estimated values of E a were 138 ± 7 kJ mol−1 for Alc. faecalis and 160 ± 8 kJ mol−1 for R. erythropolis. In the range above T BP they were 54.2 ± 1.8 kJ mol−1 for Alc. faecalis and 37.6 ± 4.1 kJ mol−1 for R. erythropolis. Discussed are the possible reasons for the observed abrupt change of the activation energy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The azo dyes wasting from different branches of the industry are among the most widespread pollutants of water basins (Clark and Anliker 1980; O’Neill et al. 1999). These synthetic compounds are regarded as typical xenobiotics due to the presence of one or several azo bonds in their structure which renders them high resistance to microbial attack (Stolz 2001). Microbial degradation of the azo dyes starts with a reductive split of the azo bond resulting in formation of colorless products.
The micro-organisms are ectothermic, i.e., the environmental temperature directly establishes organismal temperature. Microbial cell responds to a temperature change by adaptation to its new value via biochemical mechanisms. Being enzymatic these mechanisms are themselves subjected to the influence of the temperature change. On this reason temperature is a factor of paramount importance for all processes associated with microbial vitality including the remediation of water and soil.
In our previous works we reported on the azo bond reduction kinetics of the mono-azo dye Acid Orange 7 (AO7) caused by two bacterial strains of opposite Gram-staining type: the Gram-negative Alcaligenes faecalis 6132 and the Gram-positive Rhodococcus erythropolis 24 (Mutafov et al. 2006). Reported was also the effect of some surfactants of different ionic characteristic on the decolorization process (Avramova et al. 2007). This study is dedicated to the effect of temperature on the decolorization reaction observed by the same bacteria.
Materials and methods
Micro-organisms
Bacterial strains A. faecalis 6132 and R. erythropolis 24 were from the microbial collection of the Department of Microbial Biochemistry.
Reagents and media
Acid Orange 7 for microscopy (Fluka Chemie AG, Buchs, Swiss) (Fig. 1) [synonyms: AO7; Tropaeolin 000 No. 2; 4-(2-hydroxy-1-naphthylazo)benzenesulfonic acid sodium salt] was used as a model mono-azo dye. Nutrient broth Standard I (Merck AG, Darmstadt, Germany) was used for cultivation of the micro-organisms. All other reagents were of analytical grade.

Structures of the mono-azo dye Acid Orange 7 [4-(2-hydroxy-1-naphthylazo)benzenesulfonic acid sodium salt] and the products formed after reductive split of the azo bond
Cultivation conditions
The micro-organisms were cultivated for 24 h at 28 °C on a rotary shaker in 300-ml cotton-plugged Erlenmeyer flasks, each containing 50 ml nutrient broth Standard I (Merck AG). The cells were harvested by centrifugation, washed and resuspended in the phosphate buffered mineral salt solution, pH 7.2, based on prescription No. 465 (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany) with omitted sources of N and S. The concentration of thus prepared suspension was adjusted to 27 mg l−1 cells (calculated as dry mass) and used immediately for carrying out the decolorization reaction.
Decolorization reaction
The decolorization reaction was carried out statically in duplicated 100-ml stoppered iodine flasks, each containing 30 ml cell suspension and 10 mg l−1 AO7. To minimize the influence of cell aging all reactions for each one of the strains were carried out simultaneously using a series of thermostatic water baths.
Analytical procedure
A sample of 1 ml was taken immediately after preparation of the reaction mixture and clarified by centrifugation. Residual AO7 was evaluated by the optical density of the clear supernatant at 483 nm (spectrocolorimeter SPECOL, Carl Zeiss, Jena, Germany). The time of the first analysis (8–10 min after preparation of the reaction mixture) was indicated as zero time.
Quantifications
The decolorization of AO7 by both Alc. faecalis 6132 and R. erythropolis 24 follows the model of first order exponential decay (Mutafov et al. 2006), \( C_{t} = C_{0} e^{{ - \lambda t}} \), where: λ stands for the decolorization rate (h−1); C 0 and C t (mg l−1) are the concentrations of AO7 at the zero time and at time t (h), respectively. For t > 0 the decolorization rate, λ, was evaluated as
For each set of n λ-values obtained at one temperature the mean decolorization rate, \( \overline{\lambda } = \frac{1} {n}{\sum\limits_{i = 1}^n {\lambda _{i} } } \), and its standard error, \( \sigma _{n} = {\sqrt {\frac{1} {n}{\sum\limits_{i = 1}^n {{\left( {\overline{\lambda } - \lambda _{i} } \right)}^{2} } }} } \), were calculated. \( \overline{\lambda } \) was then used for analyzing the temperature effect according to the Arrhenius (1889) equation,
In Eq. (2) T stands for the absolute temperature in Kelvins (K = °C + 273.15), R is the universal gas constant (8.314 J mol−1 K−1), A is a pre-exponential factor expressing the empirical dependence of the rate coefficient on temperature, and E a is the activation energy (kJ mol−1). The normal logarithms of the evaluated mean values of the decolorization rates, \( {\text{ln}}\ifmmode\expandafter\bar\else\expandafter\=\fi{\lambda } \), were plotted against the corresponding reciprocal values of the absolute temperature, \( \frac{1} {T} \), in accordance with the linearized form of Eq. (2),
The activation energy, E a , was estimated by the tangent of the angle α which the straight line describing the function {\( \ln \overline{\lambda } ,\,\frac{1} {T} \)} forms with the abscissa: \( {\mathop E\nolimits_a } = R\tan \alpha \), where \( \tan \alpha = \frac{{{\text{d}}{\left( {\ln \overline{\lambda } } \right)}}} {{{\text{d}}{\left( {1/T} \right)}}}. \)
A program of for two-part linear least-squares regression analysis (Yeager and Ultsch 1989) was used for estimation and statistical evaluation of the regression coefficients and for localization of the break points on the Arrhenius plot.
Results
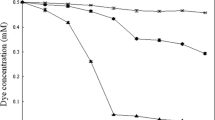
The plot of \( - {\text{ln}}\,({\mathop C\nolimits_t }/{\mathop C\nolimits_0 }) \) against time (Fig. 2) according to Eq. (1) provides the helpful option for visual control on the stability of the decolorization process: If the decolorization follows the first order exponential decay pattern then the data form a straight line which slope equals the value of decolorization rate, λ. It is evident from Fig. 2 that at identical temperatures the absolute decolorization rates observed with R. erythropolis are considerably higher than that observed with Alc. faecalis. In this, the rise of temperature causes a stronger increase in decolorization rate with R. erythropolis. It is evident as well that, compared to Alc. faecalis, R. erythropolis is not as tolerant to high temperatures: at 46 °C the decolorization of AO7 with this strain starts with a rate lower than expected one and stops completely after 3 h incubation most probably because of a thermal inactivation of the cells. At 40 °C both strains follow the first order exponential decay pattern up to the fourth hour from the beginning of the decolorization reaction but still Alc. faecalis reveals a greater stability to the increased temperature.

Decolorization rates of AO7 by Alcaligenes faecalis 6132 and Rhodococcus erythropolis 24 at different temperatures. Explanation in the text
The aberration from the typical first order exponential decay observed with R. erythropolis at 33 and 36 °C is associated in our opinion to both the high-decolorization rate and to a supposed insufficient clarification of the reaction mixture: the extrapolation of the data showed that after 10 h incubation at temperatures 33 and 36 °C in the reaction medium would remain less than 3% of the initial amount AO7. In this, we have all reasons to suppose that at these temperatures and in the lack of any source of carbon and energy the reaction conditions can provoke certain slight partial autolysis of the cells which along with the very low concentration of AO7 is sufficient to interfere with the colorimetrical method used. On this reason for further consideration we took only the experimental points obtained in the time ranges indicated with solid lines on Fig. 2. These lines were drown by means of the estimated mean values of the decolorization rate, \( \overline{\lambda } \), summarized in Table 1.
For any of the strains studied the plot of \( \ln \overline{\lambda } \) against \( \frac{1} {T} \) according to Eq. (3) results in a discontinuous chevron-like graph (Fig. 3) which inflection point is near the middle of the temperature range tested.

Arrhenius plot for bacterial decolorization rate of AO7 by Alcaligenes faecalis 6132 and Rhodococcus erythropolis 24. The results of the two-part linear regression analysis are summarized in Table 2
The plots and the results of the regression analysis summarized in Table 2 show that in the temperature range below the inflection point (12–20 °C) the reaction of bacterial decolorization with each of the strains have high activation energy, E a , decreasing sharply at the inflection point and remaining constant at this level in the whole range of raised temperatures (27–46 °C).
Table 2 summarizes the results of the regression analysis and the estimated values of the activation energy. The numbers are rounded to four significant figures. The table includes also some additional information which could be useful for further statistical analysis. What we find important to underline here is the fact that irrespective of all differences between the strains studied (Avramova et al. 2007; Mutafov et al. 2006) the exact localization of the break point temperatures reveal their practically full identity: 20.7 °C for Alc. faecalis and 20.8 °C for R. erythropolis.
Discussion
It is generally recognized that the enzymatic reactions are background of all living processes. Therefore, it is not surprising that there is a vast majority of studies using the Arrhenius equation for interpretation of numerous indices associated with different physiological functions of various classes of organisms. Nonetheless, we know only a few reports dealing with the activation energy of microbial decolorization of azo dyes (Chang and Kuo 2000; dos Santos 2005; dos Santos et al. 2004; Yu et al. 2001). Most of these works analyze the subject either in the narrow temperature ranges having predominantly practical significance or study the decolorization of the azo dyes by extremely complex consortia of micro-organisms inhabiting the active sludge. This is probably the reason why no one of them gives any evidence for the existence of break points on the Arrhenius plot.
On the other hand, the sudden alteration of the activation energy as a response of temperature change is a well known phenomenon in microbial physiology. Break points on the Arrhenius plot have been reported for the effect of temperature on microbial specific growth rate on different substrates (Ingraham 1958; Kuhn et al. 1980; Mutafov and Minkevich 1986; Minkevich et al. 2006), biomass yield (Chistyakova et al. 1983; Mutafov and Minkevich 1986), Monod’s saturation constant (Mutafov and Minkevich 1986), thermal inactivation of microbial cells (Kuhn et al. 1980; Verrips and Kwast 1977), the nitrification rate of Nitrosomonas cells (Benyahia and Polomarkaki 2005).
The reasons for the sudden change of the activation energy are still unclear. There are two basic hypotheses considered in this connection: (a) change of physical properties of the water taking place at 15, 30, 45, and 60 °C (Kuhn et al. 1980), and (b) change of the limiting rate reaction in a succession of chained enzymatic reactions known also as hypothesis of the single “bottle neck” reaction.
The high diversity of Arrhenius break point temperatures estimated in different studies along with the values obtained in our study do not provide sound confirmation for the first hypothesis.
The change of the bottle neck reaction seems a more realistic hypothesis but it must be taken into account that the mechanism of such a change cannot be explicitly confirmed: the change of bottle neck reaction may depend on the thermal characteristic of the particular enzyme but in the extremely complex intracellular organization it may depend also on the properties of the membrane structures supporting or surrounding this particular enzyme which are themselves temperature dependent (e.g., Ceuterick et al. 1978).
Based on the experimental results of our present investigation and on some of our earlier studies (Mutafov and Minkevich 1986) we incline to support the bottle neck hypothesis.
The great differences between Gram-positive and Gram-negative bacteria and, in particular, between Alc. faecalis and R. erythropolis give no reasons to expect identical values for the activation energy measured in identical temperature ranges. Nevertheless, there is an obvious similarity in their response to temperature changes. This similarity is strongly underlined by practically identical values of the break point temperatures observed. In our opinion this fact indicates certain similarity in the biochemical mechanism of decolorization of AO7 by these strains. This point of view gains a strong support by the fact that the reductive split of the azo bond occurs outside the cell (Keck et al. 1997) and the cells of both strains had identical surrounding in the course of the experiment: a factor which is not species-dependant. In this connection it should be added only that the role of cell surrounding is not studied thoroughly but there are data confirming its decisive role for the abrupt change of the activation energy. For example, the rate of nitrification by Nitrosomonas reveals a break point on the Arrhenius plot with cells immobilized in alginate gel but not with free cells (Benyahia and Polomarkaki 2005).
Concluding remarks
The fact that the observed decolorization reaction follows the Arrhenius equation, indicating that an increase in temperature decreases the activation energy requirements, thus improving the kinetics of the reaction is, in our opinion, interesting by itself and could have certain practical significance but it could hardly be qualified as unexpected. What we find new and interesting, and what we would like to stress again, is the abrupt change in the activation energy of bacterial reduction of the azo bond first reported here. The similar behavior of two strongly differing organisms and their practically identical reaction to temperature changes demonstrated by the similarity of their Arrhenius break point temperatures is also of particular interest. The authors hope that this observation could contribute to the elucidation of the mechanisms of temperature effect on the microbial physiological parameters having both scientific and practical impact.
References
Arrhenius S (1889) Über die Reaktionsgeschwindigkeit bei der Inversion von Rohrzucker durch Säuren. Z Phys Chem 4:226–248
Avramova T, Stefanova L, Angelova B, Mutafov S (2007) Bacterial decolorization of acid orange 7 in the presence of ionic and non-ionic surfactants. Z Naturforsch 62c(1–2):87–92
Benyahia F, Polomarkaki R (2005) Mass transfer and kinetic studies under no cell growth conditions in nitrification using alginate gel immobilized Nitrosomonas. Process Biochem 40(3–4):1251–1262
Ceuterick F, Peeters J, Heremans K (1978) Effect of high pressure, detergents and phospholipase on the break in the Arrhenius plot of Azotobacter nitrogenase. Eur J Biochem 87(2):401–407
Chang J-S, Kuo T-S (2000) Kinetics of bacterial decolorization of azo dye with Escherichia coli NO3. Bioresour Technol 75(2):107–111
Chistyakova TI, Minkevich IG, Eroshin VK (1983) Growth of the thermotolerant yeast, Candida valida, on ethanol: dependences of maximal growth rate and cell biomass yield on temperature. Eur J Appl Microbiol Biotechnol 18(4):225–228
Clark A, Anliker R (1980) Organic dyes and pigments. In: Hutzinger O (ed) The handbuch of environmental chemistry. Part A, Antropogenic compounds, vol 3. Springer, Berlin, Heidelberg, New York, pp 181–215
dos Santos AB, Cervantes FJ, van Lier JB (2004) Azo dye reduction by thermophilic anaerobic granular sludge, and the impact of the redox mediator anthraquinone-2,6-disulfonate (AQDS) on the reductive biochemical transformation. Appl Microbiol Biotechnol 64(1):62–69
dos Santos AB (2005) Reductive decolourisation of dyes by thermophilic anaerobic granular sludge, Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, ISBN 90-8504-134-1, 176, p 74–75
Ingraham JL (1958) Growth of psychrophilic bacteria. J Bacteriol 76(1):75–80
Keck A, Klein J, Kudlich M, Stolz A, Knackmuss H-J, Mattes R (1997) Reduction of azo dyes by redox mediators originating in the naphthalenesulfonic acid degradation pathway of Sphingomonas sp. strain BN6. Appl Environ Microbiol 63(9):3684–3690
Kuhn HJ, Cometta S, Fiechter A (1980) Effects of growth temperature on maximal specific growth rate, yield, maintenance, and death rate in glucose-limited continuous culture of the thermophilic Bacillus caldotenax. Eur J Appl Microbiol Biotechnol 10(4):303–315
Minkevich IG, Satroutdinov AD, Dedyukhina EG, Chistyakova TI, Kaparullina EN, Koshelev AV, Okunev ON (2006) The effect of temperature on bacterial degradation of EDTA in pH-auxostat. World J Microbiol Biotechnol 22(11):1205–1213
Mutafov S, Avramova T, Stefanova L, Angelova B (2006) Decolorization of acid orange 7 by bacteria of different tinctorial type: a comparative study. World J Microbiol Biotechnol 23(3):417–422
Mutafov SB, Minkevich IG (1986) Temperature effect on the growth of Candida utilis VKM-Y-2332 on ethanol. Compt Rend Acad bulg Sci 39(1):71–74
O’Neill C, Hawkes FR, Hawkes DL, Lourenço ND, Pinheiro HM, Delée W (1999) Colour in textile effluents- sources, measurement, discharge consents and simulation: a review. J Chem Technol Biotechnol 74(11):1009–1018
Stolz A (2001) Basic and applied aspects in the microbial degradation of azo dyes. Appl Microbiol Biotechnol 56(1–2):69–80
Verrips CT, Kwast RH (1977) Heat resistance of Citrobacter freundii in media with various water activities. Eur J Appl Microbiol 4(3):225–231
Yeager DP, Ultsch GR (1989) Physiological regulation and conformation: a BASIC program for the determination of critical points. Physiol Zool 62(4):888–907
Yu J, Wang X, Yue PL (2001) Optimal decolorization and kinetic modeling of synthetic dyes by Pseudomonas strains. Water Res 35(15):3579–3586
Acknowledgment
The study was supported by the Foundation for Scientific Investigations of the Bulgarian Ministry of Education, Science and Technology, according to the contract B-1311/2003.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Angelova, B., Avramova, T., Stefanova, L. et al. Temperature effect on bacterial azo bond reduction kinetics: an Arrhenius plot analysis. Biodegradation 19, 387–393 (2008). https://doi.org/10.1007/s10532-007-9144-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10532-007-9144-4