
Abstract
Preserving landscape heritage elements and indigenous and local knowledge (ILK) is an increasingly popular approach in conservation. We focus on a globally very contentious practice, silvopastoral livestock raising, which along with other peasant practices, is slated for elimination according to projected Chilean conservation policy. We used ecological surveys to ask how central Chilean semi-arid woodlands in the locality of Alhué have responded to past human livelihoods practices, including silvopastoralism. Using interviews, we examined local ecological knowledge and uses of forest plants. We also conducted surveys on current agricultural practices. Many residents maintain a diversified, smallholder subsistence agricultural strategy. Residents identified 113 plants with 73 uses. They also demonstrated a good knowledge of woodland regulations. We found that woodlands recover well from historical disturbances over 50–100 year time scales. In fact, the presence of cattle year-round in the woodlands was associated with greater tree regeneration. We find that despite the conservation discourse, there is no evidence of a degradation problem, and we hypothesize based on our findings that eradicating peasants’ silvopastoralism and other practices could increase degradation. We recommend that the Conservation Landscape programme be used to save key traditional practices, which should be studied further to determine optimal management. We show how conflicts and misalignments within and between ILK, data, and environmental discourse can signal complex socio-ecological issues where a closer look at how the evidence fits together is necessary.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The valorisation of heritage elements related to the exploitation of resources, climate adaptation, and other relations to the environment, such as indigenous and local knowledge (ILK) is increasingly integrated with conservation (Reser and Bentrupperbäumer 2005; Hill et al. 2011; Xu et al. 2009). ILK is complex to define but refers to “the knowledge of Indigenous and local people which often pertains to social–ecological systems” (Wheeler and Root-Bernstein 2020). Category V protected areas been defended as the best way to do conservation in contexts of traditional socio-ecological systems (Dudley et al. 2010). A Category V protected area is defined as one where “ the interaction of people and nature over time has produced an area of distinct character with significant ecological, biological, cultural and scenic value and where safeguarding the integrity of this interaction is vital to protecting and sustaining the area and its associated values” (Dudley et al. 2010). Other conservation approaches, which may not form legally protected areas, such as Man and Biosphere Reserves, the Globally Important Agricultural Heritage sites of the FAO, or the Conservation Landscape scheme in Chile (see below), also try to manage landscapes to maintain multiple values derived from or dependent on traditional practices and knowledge. While it is relatively easy to recognise the importance of traditional socio-ecological features or heritage elements like sacred groves and sites of geological interest, other ILK and heritage elements are more slippery, due to incomplete information, multiple interepretations, or controversies.
Globally, interest in incorporating ILK into both ecological research and environmental management is growing (Benyei et al. 2020; Wheeler and Root-Bernstein 2020). In the context of environmental management, inclusion of ILK can be part of participative mechanisms and adaptive management (Mistry and Berardi 2016). It remains uncommon to use ILK as a source of hypotheses or models that may be further explored with scientific methods (but see Molnár et al. 2020).
The interplay between scientific inattention and attention, and the interpretation and valorization not only of ILK, but also of traditional practices and the landscapes they produce, has a long and complex history. For example, silvopastoral systems have benefited from ecological and conservation research, leading to the contemporary creation and adoption of many new versions of silvopastoralism now applied throughout Latin America and the world (Peri et al. 2016). Not all silvopastoral systems are equally valued or researched. A specific example of complex social and scientific valuations of silvopastoralism is in Central Chile. This region, composed largely of shrubland, open woodlands, and closed woodlands, as a whole has long been treated as a wasteland-in-the-making, emptied of value (Root-Bernstein 2014). In particular, the silvopastoral Acacia caven woodlands (“espinal”), where cattle and horses are grazed at low densities, have long been considered to have no ecological value: they were described as unproductive degradations of native sclerophyllous woodlands (Aronson et al. 1993; Schulz et al. 2010). Recently, however, research has suggested that espinal silvopastoral habitats have ecological importance and value: they have been shown to be a resilient pioneer habitat that establishes after disturbance and is the first stage in succession towards sclerophyllous forest (Hernández et al. 2016; Root-Bernstein et al. 2017). The espinal silvopastoral system was seen as having no social, cultural or economic value either: this area currently lacks self-identified indigenous populations, and the livelihoods practices of mestizo peasants have long been considered non-adaptive and environmentally destructive (Aronson et al. 1998). More recently, a large body of agronomical research has shown that silvopastoralism in espinal can be efficient and productive, and thus has socioeconomic value (Olivares 2016). Nevertheless, Chilean conservation officials do not appear to have accepted the ecological and socioeconomic revaluations of these habitats suggested by recent research. Consequently, the espinal silvopastoral habitats and their associated management practices and knowledge have not been understood as elements of heritage, knowledge or practice informing socio-ecological conservation. Conservation officials remain strongly opposed to silvopastoralism in native woodlands (pers. comm. C. Ravanal 2019), despite there being no research specifically evaluating silvopastoralist practices and their contributions to the ecological or socioeconomic values of these habitats.
This anti-livestock view, despite the lack of evidence, takes its cue from the international conservation discourse about livestock. Over the years, pastoralism and transhumance have gained and lost policy support (Hoffman and Boerma 2014; FAO 2011; FAO 2001; Seré et al. 1995). Global discourse tends to emphasize that on average globally, cattle contribute to global warming, drought, erosion and degradation of rangeland quality (Herrero et al. 2013; Mekonnen and Hoekstra 2012; Ripple et al. 2014). Positive effects of livestock are in fact production-system- and context-dependent, and may include fire reduction, maintenance of habitat mosaics, and seed dispersal (Mazzini et al. 2018; Gregory et al. 2010; Bruun and Fritzbøger, 2002; Brown and Archer 1988). Livestock production practices also serve as a medium for social relations, systems of value, and relationships to the environment (Barragan López and Linck 1994; Manzano Baena and Casas 2010). In particular we know little about the impacts of livestock raising practices of non-indigenous peoples living in arid, semi-arid or seasonally dry forest environments of the Americas (Barragan López and Linck 1994).
As part of an effort to contribute research to understanding how contextual environmental and socio-economic factors affect the environmental impacts of traditional livestock raising, here we focus on ILK and traditional livelihoods practices related to the native silvopastoral and schlerophyllous woodlands of central Chile, and their impacts on woodland degradation, restoration and succession. The recent history of Chile involves multiple shifts in agricultural strategies, from large fundos owned by a family and worked by their tenants, to smallholdings and cooperatives created during the agricultural reform of the mid-20th century, to partial reversal of agricultural reform and development of industrial export agriculture, which altered the scales, distributions and intensities of resource use and land use change (Wright 1982). Little is known about how the transition from fundos to smallholdings in the mid-century period impacted anthropogenic disturbance patterns in woodlands. This legacy is likely to still be shaping remnant woodlands, even as land-use change from agricultural modernization has reduced total woodland cover (Schulz et al. 2010).
Plans being considered by the Chilean government to meet their obligations under the Convention on Biological Diversity and the United Nations Framework Convention on Climate Change propose to eradicate remaining traditional peasant livelihood and cultural activities in forests and other habitats, by paying rural people to give up some practices and folklorize others for tourism (personal comm. C. Ravanal 2019). This project is in apparent conflict with a project that developed the “Conservation Landscape” concept in Chile. A “Conservation Landscape” here refers to a land-use planning framework for municipalities designed by a GEF (Global Environment Fund) project to improve the conservation of heritage elements and the environment (e.g. PLADECO Alhué 2014). In theory, Conservation Landscapes could help to preserve any ILK and traditional practices that may have currently unrecognised environmental benefits—such as silvopastoralism.
We combine various research methods to identify which elements of traditional livelihoods practices may be harmful or helpful to promoting woodland regeneration and conservation. We combine an ecological study of tree regeneration following anthropogenic disturbances, with a study of ethnobiological knowledge of woodlands and threats to them, and an assessment of local agricultural livelihood strategies. We ask how locals believe that agricultural and forest-use practices should change, compared to data on their current state and its impacts, and compare this to our data and the dominant conservation discourse.
Methods
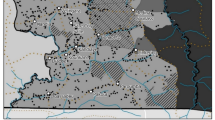
Study site. A map is shown in Fig. 1. Alhué is an interesting area with a rich cultural and ecological heritage due to its relative isolation within the valleys of the Cordillera de Cantillana in the Coastal Mountain Range of central Chile. The Alhué municipality has a population of 6,444 inhabitants and a rural population of 3,660 inhabitants (56,8 %) (INE 2017). The main socioeconomic activities are agriculture, livestock production and mining (CED 2014), with both an important gold mine and the tailings dam from the largest underground copper mine in the world within the district. Alhué municipality, with a total area of 88.476 ha, contains a number of well-conserved native sclerophyllous forests and woodlands (3.091 ha of mature native forest and 52.378 ha of secondary native forest), including those in San Juan de Pichi Natural Sanctuary (1,614 ha), and El Membrillo (16.000 ha), the compensation site for the gold mine. Most forests and woodlands in the area are private property and consist of former fundos, or latifundia. Alhué contains part of the Roblería Cobre de Loncha National Reserve, the only public protected area in the district. There are also woodlands that are owned by smallholders, as well as a woodland owned and managed by a cooperative. Local smallholders have traditional use rights in many former fundos, which their families, former tenants on the fundos, retain from before the agrarian reform in the mid twentieth century.

Map of the study sites, with major land cover types, with data from CONAF, 2013. Catastro vegetacional Región Metropolitana. Also shown, the 42 transect sites in Alhué
Fieldwork. Our sampled area was the entire Alhué municipality, which is characterized by native woodlands and other habitats including matorral shrublands especially on north-facing hillslopes (Badano et al. 2005), espinal woodlands—a kind of silvopastoral woodland dominated by Acacia caven (Donoso 1982)—and sclerophyllous woodlands containing many Chilean endemic species (Donoso 1982). Fieldwork was carried out in austral spring 2017. We repeated the methods of Root-Bernstein et al. (2017). We made transects consisting of 15 trees (> 40 cm) each (thus, of variable length), and 4 m wide. This sampling approach captures an equal number tree-tree associations (or non-associations) regardless of tree density. Transect locations were chosen in order to gain a representative sample of both typical woodland habitats with typical anthropogenic histories, and unique habitat formations or sites with unique anthropogenic histories. Unique formations included, for example, areas with rare endemic Chilean palms (Jubaea chilensis). This representative sample was created according to the advice of locals using snowballing, in the sense that at each site we asked locals (woodland users, managers, or owners) where else we should sample that was either typical of the area, or unusual. Permission to enter property and collect data was obtained from site managers or owners as relevant.
We sampled 42 transects, including one double transect (30 trees) (see Fig. 1). The double transect (no. 35) occurred at a site where the tree density was much higher than anticipated, due to a large number of trees being nursed by a single tree, which prevented sampling the formation. A double transect seemed more transparent than deleting the original sample and resampling: we correct for the doubled number of data points where it can affect statistics. At each site where we placed a transect, we oriented the transect in an arbitrary direction that allowed walking access and that crossed the vegetation type we wanted to sample. In each transect, we recorded the position of each of the 15 trees with a GPS device, as well as the species, tree height, trunk base diameter (since many trees are multi-stemmed following resprouting from the base), canopy area, and tree-tree facilitation (nurse tree) interactions, as in Root-Bernstein et al. (2017). Species identification follows Hoffmann (1979). We also recorded environmental variables, including presence of cattle or horses (using their feces as a proxy), presence of Oryctolagus cuniculus (using their feces as a proxy), presence of irrigation, and presence of a natural water source (e.g. a river or gully adjacent to the transect). Altitude data was collected by inputting the GPS position of each transect into Geoplaner.com (accessed 2018). We also recorded an oral history of anthropogenic uses of the site covering approximately the past 50 years (being the period over which most people had a detailed memory), obtained from a local user, manager or owner depending on the site. These uses or anthropogenic disturbances included house gardens, agricultural fields, livestock grazing, restoration planting and fire. Following Root-Bernstein et al. (2017), we coded the oral history, description and observation of current land-uses, along with environmental data, to create a set of explanatory variables (Table 1).
We selected sites based on local perceptions and knowledge of variation in woodland formations. While we assert that local environmental knowledge about local woodland formations is reliable and replicable, we make no assumptions about whether local perceptions correspond to scientifically and statistically assessed habitat formations. We used statistical tests to scientifically characterize the structures and compositions of our woodland sample (following Root-Bernstein et al. 2017). To assess habitat structuring, we used the Wald–Wolfowitz test. It was implemented with the ‘wawotest’ command in the adehabitat package in R version 3.1.0. This is a non-parametric categorical test that assesses the significance of “runs” or series of the same category (Siegel 1956). Each species was treated as a category, and tree number along the transect was treated as series order. Significance occurs when a given species of tree is more likely to be followed by a tree of the same species than by a tree of another species. As in Root-Bernstein et al. (2017), we defined “patchy” as runs of trees where the species were mixed in small clusters or alternating, and “gradients” as runs of trees that switched from one species at the beginning of the transect to another species at the end. These terms do not refer to spacing of the trees. We sorted transects into three habitat structures: random or “unstructured” series of tree species (including monospecfic transects), patchy habitats, and gradients. To determine whether the transects could be categorized into different habitat types in terms of species composition, we used NMDS (Non-metric Multi Dimensional Scaling) analysis. We used the Dist, metaMDS, Tree, and Group commands in the vegan package in R version 3.1.0. We used Jaccard distances since they are suitable for analysis of species composition, and the “average” command for forming clusters. We chose a height cutoff for the cluster dendrogram based on visual analysis of the tree. Both structure and species composition categories were incorporated in the set of explanatory variables.
To assess current woodland conservation status and successional capacity to overcome disturbances, we created a regeneration index for each sampled site. There are two processes that allow tree regeneration and thus woodland resilience to disturbances: tree establishment with, and without, facilitation. Tree establishment occurs in pulses in this region, following rain events (Gutiérrez et al. 2007). Tree-tree facilitation is a mechanism for habitat composition change or succession (facilitation occurs between species) (Root-Bernstein et al. 2017). Where the overall index is low, the woodland is neither reproducing itself, nor changing in composition, and therefore shows no evidence that it could recover (or is recovering) from a disturbance. Where the overall index is high, mechanisms for woodland regeneration are functioning. The regeneration index was defined as the sum of tree seedlings < 40 cm and trees > 40 cm being facilitated by a nurse tree, in each transect. For the double-length transect, we corrected the regeneration index by dividing it by 2.
We then used a zero-inflated regression model to assess whether disturbance and environmental factors explain the regeneration index. We used a zero-inflated regression because many transects (15 transects, or 36 %) had regeneration indices of 0. A zero-inflated regression produces two models. The first, the count model, uses a Poisson distribution with log link. The second, the zero-inflation model, uses a binomial distribution with log link. The count model explains what happens in sites with non-zero data, while the zero inflation model gives the odds of having 0 s. We then further explored patterns in the data using χ2 tests. All statistical tests were run in Rstudio version 1.1.453.
Ethnobiology methods. The ethnobiological study was focused on the Pichi community within Alhué (see Fig. 1), which is locally reputed to have maintained considerable traditional botanical knowledge. The profile for the sample was adults over 18 years old who lived in Pichi and had experience in the woodlands, possessed knowledge about plants, and were influential in the community. Participants were found first by consulting a list of “key actors” in environmental issues compiled by a community group (“junta de vecinos”) and the PRODESAL smallholder outreach office; further individuals were found using snowballing. Snowballing is a common method used in social sciences to obtain a sample of people to interview, in which participants comprising an initial target or convenience sample recommend other people to be interviewed, which continues until no new contacts or no new information is obtained (Mendieta 2015; Rust et al. 2017). The sample included 31 adults, of which 15 were men and 16 women, were interviewed between February and March 2017. Participants were interviewed in their homes, or while carrying out agricultural tasks. Participants were given a written explanation of the study and signed an informed consent form. Each participant was given a questionnaire of closed questions focussing on socio-demographic information. Each participant was then interviewed to determine knowledge of sclerophyllous woodland plants and their uses. Participants were then asked open questions about the problems affecting the native woodlands in Pichi in the context of the Conservation Landscape initiative. Uses of plants were assorted by similarity and coded. Finally, a participative mapping workshop including a subset of available participants (3 women and 1 man) was carried out to understand the changes over time in the environment and landscape of Pichi.
Agricultural livelihoods surveys. Agricultural models in central Chile have shifted considerably during the 20th century—for a more detailed account for this field site, see Root-Bernstein et al. (2020). The pattern of agricultural strategies is thus the outcome of several different political and economic processes. We set out to capture the diversity of existing agricultural strategies in Alhué with a survey instrument. The survey instrument was integrated into a larger questionnaire and project, which is described in Root-Bernstein et al. (2020). Here, we report only the data relevant to this study and from Alhué. The questionnaire was applied to farmers, including both smallholders and large landowners, who were located through snowballing and with the support of the Alhué Smallholder Development Program PRODESAL. Our sample was stratified by farm size, using three categories: < 10 ha, 10–100 ha, > 100 ha. Interviews were carried out in Alhué between February- August 2017. Farmers were interviewed at their homes. They were orally informed that their participation was voluntary and anonymous, consent to participate was obtained orally, and they were given a printed form of the contact information of the research team member carrying out the survey. Here we report survey data from closed and open questions and Likert scale questions about farm production, livelihoods, environmental values, knowledge of forest regulations put in place by CONAF (the National Forestry Service), and socio-demographic questions. The surveys were read aloud to participants, and answers were recorded by a research team member.
We created indices to summarize farmers’ agricultural and livelihood strategies, socioeconomic profiles and knowledge about forest regulations. The ranges of values that went into the indices were set by the total dataset from the larger study that included farmers in two other municipalities to the north and south of the Cordillera de Cantillana (see Root-Bernstein et al. 2020), Pintué and Doñihue (see Fig. 1). The indices thus implicitly provide a view of how Alhuino farmers compare to the observed range of regional farming strategies, socioeconomic conditions and knowledge. The formulas for these indices are given in Table 2. In order to detect trends in the studied population, we tested each index against normality using the Kolmogorov-Smirnov test as implemented by the command “lillie.test” in the nortest package in Rstudio version 1.1.453.
Ethical note: The ethnographic study, carried out by B.H., obtained approval from the ethics committee of the University Alberto Hurtado, because it was designed within the framework of the “Anthropology of the Forest” project led by Dr. Juan Carlos Skewes of that university. The agricultural questionnaire, carried out by A.B. with assistance from M.G.G. under the supervision of M.R.-B. who worked at INRA, was not required to be submitted to an ethics committee. At INRA, the ethics committee does not pronounce on particular research projects. Researchers are guided by a “Deontological Charter”.
Results
Ecological results. Across the 42 transects combined, 33 tree species were recorded, of which 23 are endemic to Chile. Segovia et al. (2013) map on a 10 km x 10 km scale an expected 35 species present in the local area where the study took place. Hoffmann (1979), our own source for identification, lists 42 tree species of tree found in central Chile, of which 34 are described as endemic to Chile. Of the 33 species that we recorded, we found several non-native but non-invasive trees. Robinia pseudoacacia, Ailanthus altissima, and Crategus oxicanta, all ornamental trees, and a Prunus sp., a fruit tree, appeared to have been planted and were found only in two very specific anthropogenic locations including adjacent to an abandoned house and at the site of a shrine in some rocks (see below). We also found walnut planted as part of a walnut plantation intermingled within the native woodland, and eucalyptus planted as part of an afforestation project from the 1990 s. This leaves 27 non-planted species in our sample, which includes between 77 % and 64 % of the possible species in the local area or central Chilean region, respectively (Segovia et al. 2013; Hoffmann 1979).
Of a total of 626 trees > 40 cm sampled, L. caustica was the most common (117 trees), followed by P. boldus, A. caven, C., tebo/trevo (Retanillo trinervia) and Q. saponaria, all of which species are endemic or native trees typical of early to intermediate successional-stage woodlands linking espinal to schlerophyllous forest formations. Specifically, L. caustica and A. caven can be described as pioneers that establish post-disturbance in open areas, with R. trinervia a pioneer especially common after fire, while P. boldus and Q. saponaria are frequently facilitated by A. caven to form what can be considered tree associations typical of a second step in succession post-disturbance (Root-Bernstein et al. 2017). A rare but notable species present in the sample was the endangered endemic Chilean palm (Jubaea chilensis), in both natural and planted populations. According to ILK, various populations of this species have persisted in Alhué at least in part because their seeds are dispersed by cattle (see Fleury et al. 2015). All accessible areas of Alhué appear to have experienced extensive anthropogenic disturbance in the form of charcoal production, collection of leaf litter, bark and leaves, and agriculture. A typical landscape formation throughout Alhué corresponds to a now-abandoned historical pattern of clearing mountain slopes with fire, followed by a rotation of wheat, beans or potatoes, and fallows grazed by livestock (Fig. 2).

Observed woodland structure along mountainsides in Alhué. The single-trunk Acacia caven in espinals in former agricultural fields have never been cut or burned. The “dense woodland” and “high closed-canopy forest” are typical sclerophyllous forest formations
The MDS analysis of tree co-occurrence in transects, with a cut-off of h = 0.65, yielded 9 distinct clusters. These corresponded to two groups of silvopastoral espinals, espinals entirely dominated by A. caven (suggesting chronic heavy disturbance), gully habitat, reforested areas near a river, another set of reforested areas including planted J. chilensis, the two planted anthropogenic sites (abandoned house and shrine), and dense sclerophyllous forest at high elevation. The remaining sites, sclerophyllous forest with a mix of non-schlerophyllous native species corresponding to early and later successional stages, formed the largest group (Fig. 3). This grouping does not correspond exactly to how locals identified woodland habitats. Locals did not spontaneously distinguish between different forms of espinal, and they were more likely to distinguish different schlerophyllous formations based on site history or the presence of particular species, although these sites form a single group according to our MDS analysis.

Dendogram and plot of clusters from MDS analysis of tree composition. The dotted line on the cluster dendogram indicates the cluster cutoff of h = 0.65. The main cluster of schleropyllous woodlands is indicated on the cluster plot
The range of values for the regeneration index was 0–18. In Table 3, the count model explains what happens in sites with regeneration (regeneration index > 0) while the zero inflation model gives the odds of having zero regeneration. We found that among sites with regeneration (count model), both water and cattle presence are the two variables that explain the observed regeneration index; both increase regeneration. The measure of cattle presence was the only significant variable found for the zero-inflation model: the absence of cattle is associated with an increase in the odds to exp(− 1.64) = 0.19, equivalent to a 16 % probability, of having a regeneration index of 0.
Because the results that livestock presence is good for woodland regeneration, and its absence leads to lack of regeneration at least a sixth of the time, may be surprising, we examined this result in more detail. Using χ2 tests, we looked at the relationship between spring-only vs. year-round livestock pasturing on seedling number, rather than the regeneration index. We focus on seedlings because one possibility is that livestock are seed dispersers for certain tree species, in addition to being seedling predators and agents of soil erosion and trampling: they may either increase or decrease seedlings (Bruun et al. 2002; Mazzini et al. 2018). There were fewer seedlings in areas with spring-only cattle grazing (χ2 = 14.119, df = 7 p = 0.0491), but there was no difference in seedling number between sites with year-round cattle grazing and the rest of the sites (χ2 = 6.4762, df = 7, p = 0.4854).
Ethnobiology results. Of the 31 adults interviewed, 16 were female and 15 male, and 39 % were over 60. They represented 24 households out of the 100 in Pichi. 61 % had not finished high school, but 19 % had a university degree. 64 % of participants were born and raised in Pichi, and 70 % had lived there for more than 20 years. 45 % made a living from agriculture, apiculture or reforestation activities, 19 % worked in public administration, and 29 % were retired and lived off a pension. However, many pensioners did not have enough money to live on from their pension alone and supplemented it with agricultural production and forest products for their own consumption.
In total, 128 names were given for 113 plant species, 80 of which were native species. To put this in context, a compilation of ethnobotanical records for all of Chile found 995 native “useful plant species” (Díaz-Forrestier et al. 2019). Given that our study area in theory contains close to 50 % of native plant species found in Chile (Segovia et al. 2013), we might expect 400–500 plants to be named as useful, which seems to indicate that existing local knowledge is a small subset of potential knowledge of local plant uses. The four species most commonly named were the four endemic sclerlophyllous trees boldo (Peumus boldo), quillay (Quillaja saponaria), peumo (Cryptocarya alba) and litre (Lithrea caustica), typical of relatively dense woodlands in a successional stage following open espino (Acacia caven) woodlands, and of shaded south-facing mountain slopes. The next most commonly named species, patagua (Crinodendron patagua), roble (Nothofagus sp.), lingue (Persea lingue) and canelo (Drimys winteri), are endemic trees typical of riverbanks and very moist, shaded, dense woodlands. Canelo is sacred in the indigenous Mapuche culture. Finally, the next most named plant was the espino (A. caven), typical of open, early-succession woodlands. 37 % of named species were trees, 40 % were shrubs, 15 % were herbs, and the rest were ferns, creepers, cactus, and bromeliads.
73 uses of plants were recorded, which were grouped into 14 classes (Fig. 4). The majority (66 %, 73 species) had medicinal uses. 4 % (4 spp) were recorded as having no use, and 4 spp had syncretic ceremonial uses. “No use” included one species, Prosopsis chilensis, which is endangered and rare and thus reported to be protected by the community (we could not confirm that this species was present). The plant species named as having the greatest importance for woodland conservation were the endemic sclerophyllous trees quillay (Q. saponaria), boldo (P. boldus), and litre (L. caustica), because they “produce leaf litter” and grow quickly, and one tree species named because of its association with water, the endemic patagua (Crinodendron patagua). There was little agreement on changes over time in tree species. When naming trees that used to be more numerous in the past, 23 % of participants named C. patagua, 23 % named Q. saponaria, and 10 % named C. alba. However, 10 % thought Q. saponaria was more numerous today than in the past, and 16 % identified C. alba as more numerous today than in the past. When thinking of species no longer seen in the forest, 28 % of participants identified canelo (Drimys winteri), and 21 % said the same for C. patagua.

Distribution of recorded uses of native flora in Alhué
Participants identified a set of threats to the woodlands in Pichi. These were classified post-hoc as related to fire, water scarcity, tree felling, poor management and bad behaviours, mining, floods, road construction, and invasive species and plagues. 87 % of participants identified wildfires as a princpal threat, while 58 % identified a water scarcity factor as a principal threat (threats were not ranked, so percentages do not sum to 100 %). Water scarcity was frequently blamed on climate change. Participants described a clear reduction in rainfall, snow, running water, volume in local rivers, and flood events during their lifetimes. When asked to look to the future of the sclerophyllous forest, the majority of participants expected the forest to diminish in extent, and a few even thought it would totally disappear (Fig. 5). Reasons given for decline included fires, drought, and perceptions of devalorization and poor management of the woodlands. Proposed solutions for woodland conservation during the participatory mapping activity included zonification of the woodlands with fences and exclusion of livestock, education and outreach around forest management and conservation, and training in sustainable agricultural and forest exploitation practices.

Perceptions of the future extent of the sclerophyllous forest by participants in the participatory mapping exercise. Numbers indicate the illustrated percent agreement with each answer
Agricultural survey results. We surveyed 34 farmers in Alhué. Alhué has a mixed profile of industrial fruit production and smallholders producing both for their own consumption and for the market. However, the distribution of landowners across farm sizes is highly skewed towards smallholders, with 52 % having farms < 10 ha, and 31 % with farms 10–100 ha (INE 2007). We attempted to reflect this in our stratification, although we under-sampled farmers with landholdings > 10 ha. We interviewed 33 farmers who held title to < 10 ha, 0 between 10 and 100 ha, and 1 with > 100 ha. The mean farm size, considering land to which the farmer held land title, was 7.6 ± 5.99 ha. This mean farm size includes 6 individuals in the sample who owned woodland or matorral (shrub habitat). These 6 woodland and matorral areas had a mean size of 3.75 ± 2.9 ha. Mean extent of agricultural fields or non-wooded pastures per farm was just 3.9 ± 2.9 ha. Only five of the 34 sampled individuals rented land in addition to the land to which they held title, ranging from 0.05 ha to 3000 ha. We recorded 70 distinct crops and 16 animals being raised. Livestock, which included cattle, horses and sheep, are raised in a free-range, low-input manner on private lands, communally managed lands, or on former fundos via traditional access rights. Generally livestock production is not market-oriented but used for subsistence and as a form of savings or capital.
In Fig. 6 we show the data from the indices of agricultural diversity, agricultural model, agricultural capital and lifestyle. The Lifestyle index, the Agricultural Diversity index, the Agricultural Model index and the Knowledge of Forest Regulations index were all significantly different from normal in distribution (K-S test: Life-style index, D = 0.17003, p-value = 0.01394; Agricultural Diversity, D = 0.32247, p-value = 8.663e-10; Agricultural Model, D = 0.21299, p-value = 0.0005716; Knowledge of Forest Regulations, D = 0.33184, p-value = 2.113e-10). Only the Agricultural Capital index was not significantly different from a normal distribution (D = 0.12757, p-value = 0.1727). A non-normal distribution suggests that a non-random process, here for example some structural features of the socio-economic context, has biased the distribution. The large number of non-normal distributions among the indices illustrates the socio-economic inequality in this region (Fig. 6).

Indices summarizing farmer livelihood profiles and strategies
Discussion
The ecological evidence suggests that the Alhué area has a diverse set of endemic woodland tree species, which as we show here, mainly assort into a single habitat type characterized by a mix of species typical of a successional transition between espinal and mature schlerophyllous forest, as found at other sites in central Chile (Root-Bernstein et al. 2017). Unsurprisingly in this semi-arid habitat (Gutiérrez et al. 2007), the regeneration index was highest where there was water available in the form of a river or gully. The presence of irrigation, in planted woodland restoration sites, was not associated with natural regeneration. Restoration sites may differ from other sites in that they generally lack adult trees that provide shade, which is important for tree-tree facilitation in this region (Root-Bernstein et al. 2017). Currently, it is illegal in Chile to use plant-plant facilitation in restoration projects. Adult trees’ shade and branches are also important for attracting livestock and perching birds, which are dispersers for several native trees (Olivares 2016). Restoration plantings currently appear inadequate to jumpstart natural regeneration processes.
Perhaps more surprisingly, the regeneration index was also highest in the presence of extensively grazed livestock. When we looked closer just at abundance of tree seedlings under 40 cm, we found that year-round cattle presence by itself did not explain seedling presence. This suggests either that cattle neither benefit (e.g. through seed dispersal) nor harm (e.g. through seedling herbivory or trampling) tree regeneration, or that these two effects balance out. The general benefit of extensive livestock grazing must be via a mechanism or interaction that does not directly affect seedling abundance. Rather, livestock might affect, for example, the spatial distribution of those seedlings, and thus their ability to benefit from tree-tree facilitation (Root-Bernstein et al. 2017), or their competitive advantage (see below). However, spring-only cattle pasturing was harmful to tree seedlings: there were more seedlings in areas with no cattle and with year-round cattle, compared to areas with spring-only cattle. One mechanism that may explain this would be selective foraging of grasses and herbs but not tree seedlings by experienced livestock, as opposed to unselective foraging by naïve livestock. Under this hypothesis, which was suggested to us by a local farmer during a session where we presented our study results, keeping cattle out of woodlands during most of the year prevents livestock from learning to forage selectively. Tree seedlings are generally lacking in sugars, and so may be non-preferred as food sources (Wohlleben 2015). Effects of social learning and experience on cattle foraging preferences have indeed been reported in rangelands research (Provenza and Ralph 1988). As suggested by another interlocutor, cattle and horses may also selectively eat the grass under tree canopies at the end of winter, which could reduce tree seedling-grass competition and facilitate seedling establishment if they emerge after winter grass grazing (Smit et al. 2015). Spring-only pasturing, starting after grasses and herbs have been allowed to accumulate biomass, may occur too late to provide an opening for trees to germinate under canopies.
Cattle might nevertheless be assumed to lock woodlands into early-successional states that reproduce themselves without allowing mature schlerophyllous forest to form. The existence of facilitation of schlerophyllous trees and succession by nucleation, in the presence of the same cattle raising practices, does not support this interpretation (Root-Bernstein et al. 2017). In addition, schlerophyllous forest is also reported to reform from its own fragments, meaning that it is not necessarily dependent on the successional trajectory from espinal (Hernández et al. 2016).
Before discussing the congruence between the ILK we report and our ecological results, we consider the implications of the observation that Alhuino ILK is less than what might be expected. There are no indigenous groups in central Chile whose knowledge of plant uses can be compared to this rural mestizo population, but Mapuche communities further south are reported to identify, e.g. 47 edible wild plants (Barreau et al. 2016), or 89 medicinal plants of which 47 are native (Estomba et al. 2006). These numbers are also not as high as one might expect. This may represent a loss of social relationships and relationships to the woodlands, related to the history of changing socioeconomic conditions, policies, and land uses (Barreau et al. 2016; Parra et al. 2019). These residual or relictual ethnobiological knowledges are reduced in quantity but not in quality (Barreau et al. 2016; Parra et al. 2019). In our interpretation, Alhuinos’ working knowledge of their local woodlands is superior in many respects to the scientific literature on central Chilean woodlands, which itself is underdeveloped compared to that in other countries (e.g. Rackham 2006), due in part to the scientific biases discussed in the Introduction.
There were some areas where reported ILK corresponds well to the scientific literature and our ecological results. The four trees most cited in the ethnographic interviews were also four of the five most common tree species in the local ecological transects (Q. saponaria, P. boldus, C. alba, and L. caustica). Respondents gave reasons for their expectation of decline or even disappearance of woodlands (Fig. 5) related to climate change and management practices. Whether cattle pasturing is a form of poor management is contentious, as many claim that such practices may be necessary for adaptive management to climate change, e.g. fire control (see below). Local reports of a reduction in rainfall and water volume, and the risks of drought and fire, are echoed in research demonstrating that a long mega-drought has taken place in central Chile, and that fires are increasing (Garreaud et al. 2017; Bowman et al. 2018). Although Alfonso et al. (2017) suggest that rural Chileans are not aware of the magnitudes of climate change impacts, we show that Alhuinos percieve climate change as a threat, as do smallholders globally (e.g. Ayanlade et al. 2017).
By contrast, another set of issues was more ambiguous and points to areas where further research is needed to clarify socio-ecological dynamics. These include reported beliefs about cattle management, leaf litter collection, and agricultural training for development. These are all highly pertinent and unresolved local issues that draw on global discourses about best practices.
The ethnobiological results show that local residents believe that cattle should be zoned out of forests in order to conserve forests better. Exclusionary zoning of different kinds is a globally common conservation intervention (e.g. Cotroneo et al. 2018), which, however, is increasingly questioned in different contexts due to the socio-ecological impacts it can have (e.g. Brown and McDonald 1995). The idea of keeping livestock out of woodlands was also contested by other local individuals, outside the ethnographic study. Local farmers claimed that the cattle are good for the forest because they eat all the grass that will otherwise dry up and feed wildfires (pers. comms. to MG-G, MR-B; see also discussion of leaf litter and forest fires below). It is difficult to assess whether the claim that cattle should be excluded from forests represents Alhuinos’ ILK, or whether it represents an attempt to position themselves as “environmentalist” (as they believe themselves to be, see Root-Bernstein et al. 2020) by aligning with CONAF and PRODESAL, which in turn are aligned with the dominant global discourse against livestock. Alternatively, by “zonation” they may be refering to traditional local practices of community-managed rotational harvest of bark, leaves, and leaf litter, and transhumance and the spring-only pasturing discussed above. Rotational zonation and transhumance can lead to landscape-level ecological connections (Manzano et al. preprint), but timing issues may be important.
The ethnobiological results also highlight a claim that leaf litter/humus formation is important to maintain the forest. Our knowledge of decomposition rates of schlerophyllous hardwood litter, and their interaction with mediterranean or South American habitat weather and climate patterns, is incomplete (Bani et al. 2018). The interactions between leaf litter, soil, and nutrient cycling are complex, and it is not necessarily the case that a greater accumulation of leaf litter points to richer soil, if the rate of decomposition is particularly slow under drought or species composition change, for example (Hobbie 2015; Krishna and Mohan 2017; Bani et al. 2018). Leaf litter traps soil moisture, and can improve the germination rate of species such as the endangered endemic Bielschmiedia miersii (Becerra et al. 2004), and probably other species as well. However, leaf litter might also prevent the germination of trees with other requirements. During fieldwork we also encountered a view held by local arrieros (cowboys) that woodlands should be kept “clean” through a combination of livestock grazing and leaf litter collection. Leaf litter collection presumably removes nutrients from the system, although the traditional 10-year rotations should allow some nutrient leaching (Krishna and Mohan 2017; Bani et al. 2018). Livestock grazing of the understory should relatively increase nutrient cycling rates, especially as wild herbivores that might have done so in the past are absent (Schmitz 2008). Critically, a ‘clean’ forest may have a lower risk of intense forest fires (Gómez-González and Cavieres 2009). We predict that the timing and patchiness of leaf litter collection and cattle grazing will modulate between possible benefits to fire control vs. tree establishment.
Finally, some local residents expressed the view that training in sustainable agricultural and woodland exploitation practices would secure forest conservation. With this claim, local residents are clearly aligned with global best practice in adaptive and community-based conservation (e.g. Staiger-Rivas et al. 2012). However, if we take this alignment as proof that their opinion is justified, we would be ignoring our empirical evidence. The agricultural results point to a set of agricultural and exploitative practices with notable diversity and apparent flexibility, related to past and current rural poverty, as suggested by our Agricultural Capital and Lifestyle indices. Our data shows that Alhuino smallholders tend to have an independent, isolated lifestyle that is moderately integrated with services and infrastructures. This lack of integration can act to maintain dependency on woodland resources (e.g. firewood) and to maintain (some) ILK and management knowledge, while also representing a lack of integration with the logics and capitals that allow intensified agricultural exploitation. Residents also showed good knowledge of forest regulations. Our own evidence indicates that the historical and current traditional anthropogenic disturbances throughout the valley of Alhué have not resulted in widespread degradation. The patchy legacies of historical and present land uses in Alhue (e.g. Fig. 2) may allow the preservation of seed sources for woodland regeneration, as elsewhere (Armesto et al. 1991). Small-scale, rotational and diversified agricultural livelihoods produce patchy disturbances, and are globally associated with better forest conservation outcomes than alternative agricultural strategies (Allen et al. 2014; Perz 2004).
Why then do locals ask for training in sustainable agriculture and woodland exploitation? It would be naïve to read this as a real lack of know-how. Some locals may believe that their own knowledge is illegitimate (which might reinforce its loss, see above). Capacity building, to promote entrepreneurial values, is also an important feature of the Chilean approach to reducing poverty (Mayol et al. 2013). The rural poor are thus likely to have an expectation that training leads to social mobility and economic development. In addition, the recorded perception of climate change risks may contribute to the desire for adaptive management techniques in order to face an uncertain and changing climate, although we did not specifically ask about this.
There may be forms of sustainable agriculture in which locals could be trained that could lead to economic development while also legitimizing low-input diversified agricultural models (e.g. Bucher and Huszar 1999, Ponnusamy and Pachaiyappa 2018), which might thus conserve local woodlands. In contrast, the currently proposed interventions aimed at altering or removing anthropogenic uses of woodlands may, as we discuss above, reduce tree recruitment and increase fire intensity. These recommendations should be tested before people are trained in them.
Category V-type protected areas and programmes are a kind of normative tool for identifying what is and should be valued about landscapes. We find that in this context, despite the conservation discourse, there is no evidence of a degradation problem that could be fixed by eradicating peasant smallholder livelihoods. We recommend that the Conservation Landscape programme be used to save key traditional practices, which should be studied further to determine optimal management. We show how conflicts and misalignments within and between ILK, data, and environmental discourse can signal complex socio-ecological issues where a closer look at how the evidence fits together is necessary. Using ILK and ecological data, we have demonstrated clear possible mechanisms by which livestock raising in particular, as well as other traditional anthropogenic woodland uses, could play key roles, when managed properly, in woodland maintenace and recovery. If these forms of management can be developed, then land-sharing in heritage landscapes should be sustainable (Zorondo-Rodríguez et al. 2019).
Availability of data and material
The agricultural data and tree data will be made available on ResearchGate.
Code availability
Not applicable.
References
Alfonso A, Zorondo-Rodríguez F, Simonetti JA (2017) Perceived changes in environmental degradation and loss of ecosystem services, and their implications in human well-being. Int J Sustain Dev World Ecol 24(6):561–574
Allen T, Prosperi P, Cogill B, Flichman G (2014) Agricultural biodiversity, social—ecological systems and sustainable diets. Proc Nutr Soc 73(4):498–508
Armesto JJ, Pickett ST, McDonnell MJ (1991) Spatial heterogeneity during succession: a cyclic model of invasion and exclusion. Ecological heterogeneity. Springer, New York, NY, pp 256–269
Aronson J, Le Floc E, Ovalle C, Pontanier R (1993) Restoration and rehabilitation of degraded ecosystems in arid and semi-arid lands II Case studies in southern Tunisia, Central Chile and Northern Cameroon. Restor Ecol 1:168–187
Aronson J, Pozo A, Ovalle C, Avendaño J, Lavin A, Etienne M (1998) Land use changes and conflicts in Central Chile In: Rundel PW, Montenegro G, Jaksic F (eds) Landscape disturbance and biodiversity in Mediterranean-type ecosystems, Springer Verlag Berlín Heidelberg, Berlin/Heidelberg, Germany, pp 155–168
Ayanlade A, Radeny M, Morton JF (2017) Comparing smallholder farmers’ perception of climate change with meteorological data: A case study from southwestern Nigeria. Weather Clim Extremes 15:24–33
Badano EI, Cavieres LA, Molina-Montenegro MA, Quiroz CL (2005) Slope aspect influences plant association patterns in the Mediterranean matorral of central Chile. J Arid Environ 62(1):93–108
Bani A, Pioli S, Ventura M, Panzacchi P, Borruso L, Tognetti R, Brusetti L (2018) The role of microbial community in the decomposition of leaf litter and deadwood. Appl Soil Ecol 126:75–84
Barragan López E, Linck T (1994) Rancheros y sociedades rancheras: quinientos y un años de conquista ordinaria. In:Caravelle, n°63, 501 ans plus tard: Amérique Indienne 93, pp 11–27
Barreau A, Ibarra JT, Wyndham FS, Rojas A, Kozak RA (2016) How can we teach our children if we cannot access the forest? Generational change in Mapuche knowledge of wild edible plants in Andean temperate ecosystems of Chile. J Ethnobiol 36(2):412–432
Becerra PI, Celis-Diez JL, Bustamante RO (2004) Effects of leaf litter and precipitation on germination and seedling survival of the endangered tree beilschmiedia miersii. Appl Veg Sci 7(2):253–257
Benyei P, Arreola G, Reyes-García V (2020) Storing and sharing: A review of indigenous and local knowledge conservation initiatives. Ambio 49(1):218–230
Bowman DM, Moreira-Muñoz A, Kolden CA, Chávez RO, Muñoz AA, Salinas F, Borchers N (2018) Human–environmental drivers and impacts of the globally extreme 2017 Chilean fires. Ambio 4:350–362
Brown JH, McDonald W (1995) Livestock grazing and conservation on southwestern rangelands. Conserv Biol 9(6):1644–1647
Brown JR, Archer S (1988) Woody plant seed dispersal and gap formation in a North American subtropical savanna woodland: the role of domestic herbivores. Vegetatio 73(2):73–80
Bruun HH, Fritzbøger B (2002) The past impact of livestock husbandry on dispersal of plant seeds in the landscaprthwestern South America. Ecol Soc 19(4)
Bucher EH, Huszar PC (1999) Sustainable management of the Gran Chaco of South America: ecological promise and economic constraints. J Environ Manage 57(2):99–108
CED, Centro de Estudio del Desarrollo (2014) Desarrollo e Implementación Inicial del Paisaje de Conservación de la Comuna de Alhué, Región Metropolitana de Santiago SDP No 288/2013. Informe Final, Santiago de Chile
Cotroneo SM, Jacobo EJ, Brassiolo MM, Golluscio RA (2018) Restoration ability of seasonal exclosures under different woodland degradation stages in semiarid Chaco rangelands of Argentina. J Arid Environ 158:28–34
Díaz-Forestier J, León-Lobos P, Marticorena A, Celis-Diez JL, Giovannini P (2019) Native useful plants of Chile: a review and use patterns. Econ Bot 73(1):112–126
Donoso C (1982) Reseña ecologica de los bosques mediterraneos de Chile. Bosque 4:117–146
Dudley N, Parrish JD, Redford KH, Stolton S (2010) The revised IUCN protected area management categories: the debate and ways forward. Oryx 44(4):485–490
Estomba D, Ladio A, Lozada M (2006) Medicinal wild plant knowledge and gathering patterns in a Mapuche community from North-western Patagonia. J Ethnopharmacol 103(1):109–119
FAO (2011) World Livestock 2011—Livestock in food security. Rome, Italy
FAO (2001) Pastoralism in the new millennium. FAO animal production and health paper 150. Rome, Italy
Fleury M, Marcelo W, Vásquez RA, González LA, Bustamante RO (2015) Recruitment dynamics of the relict palm, Jubaea chilensis: intricate and pervasive effects of invasive herbivores and nurse shrubs in central Chile. PLoS ONE 10(7):e0133559
Garreaud RD, Alvarez-Garreton C, Barichivich J, Pablo Boisier J, Christie D, Galleguillos M, Zambrano-Bigiarini M (2017) The 2010–2015 megadrought in central. Impacts on regional hydroclimate and vegetation, Chile
Gómez-González S, Cavieres LA (2009) Litter burning does not equally affect seedling emergence of native and alien species of the Mediterranean-type Chilean matorral. Int J Wildland Fire 18(2):213–221
Gregory NC, Sensenig RL, Wilcove DS (2010) Effects of controlled fire and livestock grazing on bird communities in East African savannas. Conserv Biol 24(6):1606–1616
Gutiérrez JR, Holmgren M, Manrique R, Squeo FA (2007) Reduced herbivore pressure under rainy ENSO conditions could facilitate dryland reforestation. J Arid Environ 68(2):322–330
Hernández A, Miranda MD, Arellano EC, Dobbs C (2016) Landscape trajectories and their effect on fragmentation for a Mediterranean semi-arid ecosystem in Central Chile. J Arid Environ 127:74–81
Herrero M, Havlík P, Valin H, Notenbaert A, Rufino MC, Thornton PK, Blümmel M, Weiss F et al. (2013) Biomass use, production, feed efficiencies, and greenhouse gas emissions from global livestock systems. Proc Natl Acad Sci 110(52):20888–20893
Hill R, Cullen-Unsworth LC, Talbot LD, McIntyre-Tamwoy S (2011) Empowering Indigenous peoples’ biocultural diversity through World Heritage cultural landscapes: a case study from the Australian humid tropical forests. Int J Herit Stud 17(6):571–591
Hobbie SE (2015) Plant species effects on nutrient cycling: revisiting litter feedbacks. Trends Ecol Evol 30(6):357–363
Hoffmann J (1979) Flora silvestre de Chile. Zona central. Una guía para la identificación de las especies vegetales más frecuentes. Fundación Claudio Gay, Santiago, Chile
Hoffman T, Boerma D (2014) Ecosystem services provided by livestock species and breeds, with special consideration to the contributions of small-scale livestock keepers and pastoralists FAO Commission on genetic resources for food and agriculture Background study paper 66(1) https://www.faoorg/3/a-at598epdf
INE, Instituto Nacional de Estadistica (2017) Resultados censo de población y vivienda 2017. Santiago, Chile
Krishna MP, Mohan M (2017) Litter decomposition in forest ecosystems: a review. Energy Ecol Environ 2(4):236–249
Manzano Baena P, Casas R (2010) Past, present and future of transhumancia in Spain: Nomadism in a developed country. Pastoralism 1:72–90
Manzano P, Galvin KA, Cabeza M preprint. A global characterization of pastoral mobility types. https://www.openanthroresearch.org/doi/abs/10.1002/oarr.10000335.1
Mayol Miranda A, Azócar Rosenkranz C, Azócar Ortiz C (2013) El Chile profundo: Modelos culturales de la desigualidad y sus resistencias. Liberalia Ediciones, Santiago
Mazzini F, Relva MA, Malizia LR (2018) Impacts of domestic cattle on forest and woody ecosystems in southern South America. Plant Ecol 219(8):913–925
Mekonnen MM, Hoekstra AY (2012) A global assessment of the water footprint of farm animal products. Ecosystems 15:401–415
Mendieta G (2015) Informantes y muestreo en investigaci?n cualitativa. Investigaciones Andina 17(30):1148–1150
Mistry J, Berardi A (2016) Bridging indigenous and scientific knowledge. Science 352(6291):1274–1275
Molnár Z, Kelemen A, Kun R, Máté J, Sáfián L, Provenza F, Vadász C (2020) Knowledge co-production with traditional herders on cattle grazing behaviour for better management of species‐rich grasslands. J Appl Ecol 57(9):1677–1687
Olivares AE (2016) El espinal: Manejo silvopastoril de un recurso ignorado. Editorial Universitaria, Santiago
Parra SA, Folchi M, Simonetti JA (2019) Knowledge of native edible plants in a monoculture plantation-dominated landscape. J Ethnobiol 39(4):567–583
Peri PL, Dube F, Varella A (eds) (2016) Silvopastoral systems in Southern South America. Advances in Agroforestry, vol 11. Springer, New York
Perz SG (2004) Are agricultural production and forest conservation compatible? Agricultural diversity, agricultural incomes and primary forest cover among small farm colonists in the Amazon. World Dev 32(6):957–977
PLADECO (Development Plan for the Municipality of Alhué) (2014) Paisaje de Conservación, 2014–2020 Municipality of Alhué, Chile
Ponnusamy K, Pachaiyappan K (2018) Strengthening extension research in animal husbandry: review of issues and strategies. Indian J Anim Sci 88(2):137–143
Provenza FD, Ralph DF (1988) Development of dietary choice in livestock on rangelands and its implications for management. J Anim Sci 66(9):2356–2368
Rackham O (2006) Woodlands. Collins, London
Ravanal C (2019) Personal communication to Meredith Root-Bernstein. Fieldnotes, 18/11/2019, Meredith Root-Bernstein
Reser JP, Bentrupperbäumer JM (2005) What and where are environmental values? Assessing the impacts of current diversity of use of ‘environmental’and ‘World Heritage’values. J Environ Psychol 25(2):125–146
Ripple WJ, Smith P, Haberl H, Montzka SA, McAlpine C, Boucher DH (2014) Ruminants, climate change and climate policy. Nat Clim Change 4:2–5
Root-Bernstein M (2014) Nostalgia, the fleeting and the rare in Chilean relationships to nature and non human animals. Soc Anim 22:560–579
Root-Bernstein M, Bondoux A, Guerrero-Gatica M, Zorondo- Rodriguez F (2020) Tacit working models of human behavioural change II: Farmers’ folk theories of conservation programme design. Ambio 49, 1658–1675
Root-Bernstein M, Valenzuela R, Huerta M, Armesto J, Jaksic F (2017) Acacia caven nurses endemic sclerophyllous trees along a successional pathway from silvopastoral savanna to forest. Ecosphere 8(2):e01667
Rust NA, Abrams A, Challender DW et al. (2017) Quantity does not always mean quality: The importance of qualitative social science in conservation research. Soc Nat Resour 30(10):1304–1310
Segovia RA, Hinojosa LF, Pérez MF, Hawkins BA (2013) Biogeographic anomalies in the species richness of C hilean forests: Incorporating evolution into a climatic–historic scenario. Austral Ecol 38(8):905–914
Schmitz OJ (2008) Herbivory from individuals to ecosystems. Annu Rev Ecol Evol Syst 39:133–152
Schulz JJ, Cayuela L, Echeverria C, Salas J, Benayas JMR (2010) Monitoring land cover change of the dryland forest landscape of Central Chile (1975–2008). Appl Geogr 30(3):436–447
Seré C, Steinfeld H, Goenewald J (1995) World Livestock Production Systems: Current status, issues and trends. FAO Animal Production and Health Paper 127, Rome, Italy
Siegel S (1956) Nonparametric statistics for the behavioral sciences. McGraw-Hill, New York
Smit C, Ruifrok JL, van Klink R, Olff H (2015) Rewilding with large herbivores: The importance of grazing refuges for sapling establishment and wood-pasture formation. Biol Conserv 182:134–142
Staiger-Rivas S, Álvarez S, Ashby JA, Lundy M, Muthoni Andriatsitohaina R, Victoria PA, Russell N (2012) Strengthening capacity to achieve eco-efficiency through agricultural research for development. Centro Internacional de Agricultura Tropical (CIAT)
Wheeler HC, Root-Bernstein M (2020) Informing decision‐making with Indigenous and local knowledge and science. J Appl Ecol 57(9):1634–1643
Wohlleben P (2015) La vie secrète des arbres: Ce qu’ils ressentent, comment ils communiquent. Les Arènes, Paris
Wright T (1982) Landowners and reform in Chile: The Sociedad Nacional de Agricultura, 1919–1940. University of Illinois Press, London, United Kingdom
Xu J, Lebel L, Sturgeon J (2009) Functional links between biodiversity, livelihoods, and culture in a Hani swidden landscape in southwest China. Ecol Soc 14(2):20
Zorondo-Rodríguez F, Díaz M, Simonetti-Grez G, Simonetti JA (2019) Why would new protected areas be accepted or rejected by the public?: Lessons from an ex-ante evaluation of the new Patagonia Park Network in Chile. Land Use Policy 89:104248
Acknowledgements
BH thanks Juan Carlos Skewes and Victoria Castro for their supervision and assistance. We are grateful to numerous people who shared their knowledge and provided assistance and support, including the park rangers at the National Reserve Roblería Cobres de Loncha, the San Juan de Pichi Nature Sanctuary, Gerardo Schlack of Fundo El Membrillo, the PRODESAL Alhué office, the municipal staff of Alhué, Bernardita Castro, Benjamín Riveros, Luna Chiang, Ángela Danbach, Marta Bello, Flavia Bustos, and everyone who shared their experience and knowledge with us in the field. We miss the expertise, boundless curiosity and speculations of Rafael Valenzuela, who died in 2018.
Funding
Marie Curie FP7 COFUND Agreenskills Plus Fellowship; CONICYT PIA/BASAL FB000; Fondecyt-Iniciación 11160672; FONDECYT No. 1140598
Author information
Authors and Affiliations
Contributions
MR-B conceived of the paper idea with FZ-R, MG-G, AB, and BH. Methods for the different parts were designed respectively by BH and MR-B. BH, AB, MG-G, MR-B, MH, RV, and AVB collected the material and data. RV and AVB made particular contributions to data interpretation. BH, AB and MR-B carried out data analysis. BH and AB wrote masters theses; on the basis of these and the other material MR-B wrote the paper. FZ-R and MG-G provided editorial comments.
Corresponding author
Ethics declarations
Ethics approval
Ethics committee of the University Alberto Hurtado.
Conflicts of interest/Competing interests
None declared.
Consent to participate
For the ethnographic study, this was obtained through a signature. For the agricultural strategies study, this was obtained orally.
Consent for publication
Not applicable.
Additional information
Communicated by Lilly O Rodriguez.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article belongs to the Topical Collection: Forest and plantation biodiversity.
Rights and permissions
About this article

Cite this article
Root-Bernstein, M., Vargas, B.H., Bondoux, A. et al. Silvopastoralism, local ecological knowledge and woodland trajectories in a category V- type management area. Biodivers Conserv 31, 543–564 (2022). https://doi.org/10.1007/s10531-021-02349-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-021-02349-7