
Abstract
Understanding the patterns of biodiversity is central to species conservation planning. Here we illustrate chiropteran diversity and distribution pattern of the hill ranges of the Eastern Ghats of Odisha state, India using roost survey and mist net survey techniques. The present study recorded 23 bat species including two newly recorded species for the study site i.e., Hipposideros galeritus and Megaderma spasma. The bat capture rate was the highest in the moist deciduous forest describing higher bat populations compared with the places of human habitation. Utilities of 21 climate, three physiographic and two disturbance variables and species distribution modelling could reveal that the north-eastern part was more suitable for bat distributions than the southern part of the region. The climate, physiographic and disturbance variables were crucial in their distributions, where elevation and potential evapotranspiration were highly significant. Unlike other species, Taphozous melanopogon showed negative correlation with these variables. The mean diurnal temperature and precipitation seasonality have positive influence on bat distributions, indicating their resilience to seasonal precipitation changes and day time temperature fluctuations. The negative effect of temperature seasonality on the distribution of Pteropus giganteus could attribute to its vulnerability to climate change effects. The study provides inputs for monitoring the future spatio-temporal changes, suggesting long term conservation measures. It could generate basic information on the impacts of climate change on bat distribution in the future climate change scenario.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Understanding the patterns of biodiversity at the ecosystem, species or genetic levels, within a landscape, is essential for a wide range of fundamental and applied purposes in species conservation planning (Moreno and Halffter 2001; Paliwal and Mathur 2012). Although climate has always been a prominent factor in species distributions (Francis and Currie 2003), it is difficult to assess changes in the ecology and behaviour of any individual species to a single cause. Habitat heterogeneity and local scale environments greatly influences community structure and composition (Xu et al. 2014; Kent et al. 2014). Bats are one of the most diverse mammalian order and significant contributor to global mammalian diversity with more than 1300 species worldwide (Fenton and Simmons 2014). For smooth functioning of the ecosystem, they also play important role in pollination, seed dispersion, insect pest control as well as transfer and redistribution of essential nutrients between habitats (Kunz et al. 2011). Bats have long life span and need sufficient energy to manage their sustained power flight (Mello et al. 2009; Voigt and Lewanzik 2011) and are classic examples of biodiversity indicators. Their reproduction depends on climatic suitability and the temporal availability of food resources (Crichton and Krutzsch 2000) and they are efficient in reading climatic signals for foraging, roosting, parturition, migration, breeding and hibernation (Sherwin et al. 2013). However, their slow rate of reproduction has increased their proneness to environmental stress and steady decline in population (Jones et al. 2009). In general, insect populations vary with climatic conditions, where low temperature retards their growth (Anthony et al. 1981; Burles et al. 2009). More rainfall helps dipterans and lepidopterans to multiply rapidly (Frick et al. 2009). On the contrary, insect availability to bats is regulated due to reduced insect flight during heavy rainfall (Anthony et al. 1981). It has been reported that changes in seasonal temperature and precipitation has great control on flowering and fruiting of food plants, which influences foraging behaviour of fruit bat Eidolon hevlum by altering reproductive potential of the species (Richter and Cumming 2008). Some studies also highlight the differences in bat species responses to changes in temperatures owing to disparities in foraging, habitat preferences and reproductive needs. Myotis daubentonii is reported to be less sensitive to temperature changes owing to its efficient hunting strategy and availability of insects in plenty (Boonman et al. 1998; Ciechanowski et al. 2007), whereas aerial-hawking bats are more likely to depend on climate changes due to their vulnerability to food supply in time and space. Bats are prone to evaporation due to their large naked flight membranes (Webb et al. 1995); and need constant water supply near roosting sites (Adams and Hayes 2008). The lactating females and roosts with high solar gain are more prone to water stress (Adams 2010).
Although bats are one of the most successful and highly adaptive animals, loss and degradation of roosting and foraging habitats, emerging diseases, hunting, persecution as well as effects of climate change have emerged as major factors for declining bat population worldwide (Mickleburgh et al. 2002). The negative influences of climate change on behaviour of European and north-west African bats have been reported (Sherwin et al. 2013). Lasiurus borealis, Rhinolphus ferrumequinum, Tadarida brasiliensis and Eidolon hevlum show temporal changes in their foraging behaviour in response to availability of food resources (Park et al. 2000; Dunbar and Tomasi 2006; Newson et al. 2008; Richter and Cumming 2008). The declination of bat population is more intense in the tropical countries (Kingston 2010) and it has been forecasted to exceed by 40% in coming decades solely owing to habitat related threats (Lane et al. 2006). Therefore, adequate information on diversity and status of bats is essential for region-specific conservation measures (Bates 2013).
Species distribution models (SDM) are extrapolation techniques used to map species distributions and the probability of habitat use (Hirzel et al. 2006). Several modelling protocols are employed to enumerate relationships between species and the environment. Maximum Entropy (Maxent) is a popular SDM technique, very useful for mapping species distributions with presence only data (Phillips and Dudík 2008). Its popularity is attributed to its excellent predictive ability compared with other SDMs (e.g. Elith et al. 2006). Maxent is reported to be a preferred technique in distributions of bats especially for small sample sizes (Wisz et al. 2008) and owing to their nocturnal behaviour that impedes their identification in flight (Razgour et al. 2016). Maxent finds the probability distribution, subject to the constraints imposed by the information available from the observed occurrence of the species and environmental conditions across the study area and maps depicting the species’ expected range. It has advantages of clamping and regularisation, with abilities to examine whether projections lie within environmental space or geographic space (Murray et al. 2011).
The Eastern Ghats is one of the oldest, non-contiguous and phyto-geographically important mountain ranges in India that lies parallel to the east coast from Tamil Nadu in southern India to Odisha in the eastern India, spreading over a length of around 1750 km. Recent herpeto-faunal, mammalian and avian inventories along the Eastern Ghats suggests that its forests harbour relict population of once widely distributed wet zone species (Mohapatra et al. 2010, 2014; Agarwal et al. 2013; Nayak et al. 2014; Purohit et al. 2017). Dutta and Mohapatra (2017) reviewed the faunal composition of the Eastern Ghats and reported 32 bat species from eight families to the region. They observed that the northern parts of Eastern Ghats, particularly the Odisha state, have not been adequately explored despite its biological importance. Moreover, the Eastern Ghats landscape has become vulnerable from the rapid land cover changes in recent decades through deforestation, agricultural encroachments and infrastructure development (Reddy et al. 2014) and it might be affecting the bat population occurring in this region.
The present study detailed the inventory of bat fauna of the Eastern Ghats along the hill ranges of Odisha; and explores the underlying causes of their distributions. The habitat suitability maps of some top selected bat species are derived using Maximum Entropy model (Phillips et al. 2006). The influences of elevation, climate and anthropogenic disturbance on bat distributions are highlighted. The findings of this study could provide meaningful insights to monitor the future spatio-temporal changes and suggest long term conservation measures.
Methods
Study area
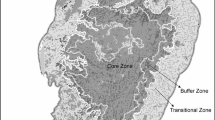
The study was carried out along the Eastern Ghats hill range in Odisha, eastern India (Fig. 1). The area lies between 17°48′ to 22°33′N and 81°23′ to 86°53′E spreading over an area of 73,700 km2. The hill ranges of the Eastern Ghats are not contiguous and highly fragmented with hilly and undulating terrain with plain areas. The maximum elevation reaches up to 1672 m, where the southern part has a higher elevation than the northern part. The Eastern Ghats experiences tropical climatic condition with three distinct seasons i.e., summer, monsoon and winter. It receives an annual rainfall of 1200–1500 mm from both southwest and northeast monsoon. The temperature varies from a minimum of 12 °C during winter to a maximum of 38 °C during summer (Ray 2005). The forest types of Eastern Ghats hill ranges of Odisha are broadly classified into the tropical moist deciduous, tropical dry deciduous and few patches of semi evergreen forests (Champion and Seth (1968). However, the forest cover of Eastern Ghats has been declined due to agricultural expansion, timber harvesting, shifting cultivation and development of industries (Saranya and Reddy 2016).

The study area showing different land cover pattern and bat sampling grids along the Eastern Ghats boundary of Odisha state, India. (Color figure online)
Sampling sites
A base map of the study area (Fig. 1) encompassing the known forest cover zone following Champion and Seth (1968) was developed using the Geographic Information System in ArcGIS version 9.3. The detailed land use pattern within each forest zone was then classified as natural forest, agricultural land and human habitation by digitizing over latest Google Earth imagery. 20 km × 20 km grids were overlaid throughout the study area covering all the habitat types (strata); and sampling sites were selected following Guthrie (2012). A total of 30 grids (20 in moist deciduous forest, nine in dry deciduous forest and one in semi evergreen forest) were selected for sampling (Fig. 1). Tropical moist deciduous forest, characterised by moderately dense vegetation with heterogeneous canopy layer is widely distributed throughout the study area. Its major plant species are Shorea robusta, Syzygium cumuni, Mangifera indica, Terminalia tomentosa and Anogeissus latifolia. Tropical dry deciduous forest has comparatively open vegetation; and distributed along the central and western parts of the study area. Its dominant plant species are Tectona grandis, Bombax ceiba, Madhuca indica and Dendrocalamus strictus. Tropical semi evergreen forest is found to occur in the foot hills and valleys > 600 m elevation i.e., in few patches Similipal Biosphere Reserve, Kerandimal hills and Deomali hills along northernmost, central and southern parts of the study area, respectively. The dominant plant species of this forest type are Terminalia arjuna, Mangifera indica and Diospyros embryopteris. Agricultural lands include some tree species i.e., Ficus benghalensis, Ficus religiosa, Mangifera indica, Bombax ceiba, Borassus flabellifer. Non-cultivable areas are dominated by Lantana camara and Chromolaena odorata. Human habitation are with minimal natural vegetation cover, but some seasonal fruiting trees like Mangifera indica, Ziziphus mauritiana, Psidium guajava and Musa sp. are witnessed in the backyard.
Bat survey
The bat fauna survey was carried out for three years during May 2015 to June 2018. The roost survey and mist net survey techniques were adopted following the guidelines and field protocols of Bat Conservation Trust (2007). In each sampling grid, information on occurrence of bat roosting sites were collected by interviewing the local people and Forest Department staffs owing to their adequate knowledge on local biodiversity. Subsequently, the identified bat roosting sites were surveyed involving them. Additionally, caves, crevices, tree holes, deserted buildings and old temples situated nearby the surveyed roosts were searched. The roost survey was carried out during the day. At each active roost, some bats were captured using an entomological net directly or mounted over an extendable pole to identify species with a minimum disturbance. Species were identified following Bates and Harrison (1997) and Srinivasulu et al. (2010) adopted for the Indian bats. For the present study, species with confirmed identification were considered. For identification of those preferring to roost in crevices, tree holes and other inaccessible locations, we used mist nets. In each 30 grids selected for sample survey, three different locations were considered on the basis of their natural vegetation, agricultural land and human habitation. Total 90 locations covering 20 in moist deciduous vegetation, nine in dry deciduous vegetation, one in semi evergreen vegetation, 30 in agricultural lands and 30 in human habitation were sampled (Table 1). Sampling sites were revisited for at least three times during the study period. Netting activities at each location was carried out for three hours per sampling night just after sunset.
For mist netting, one 15 m2 net with 13 mm mesh size and four sleeves (Avinet USA) was used and deployed across forest trails, streams and near water bodies along flyways of bats (Barlow 1999). Before mist netting, a preliminary investigation on availability of space, surrounding vegetation, availability of water holes, suspected flight path and kind of obstructions was carried out during daytime. Nets were set below canopy level either by stalking them with bamboo poles or with tree canopies following the techniques described by Humphrey et al. (1968). One additional mist net was kept for reserve to replace the damaged ones as bats damage nets after entanglement. Whenever nets were placed over water bodies or streams, intensive care was taken not to keep the lowest edge of the net near water surface as far as possible ensuring that it is high enough to prevent any bats that are captured in the bottom of the mist net from hanging in the water (Kunz 1988). After deployment, nets were checked at every 15 min interval to ensure that all the captured bats are removed quickly and prevent any of them from escaping. Bats captured in the net were immediately released with utmost care without any physical injury. If any pregnant or lactating bat was caught in the mist net, it was immediately released without further examination. All captured bats were kept in hung up position in cloth bags, and none were kept for more than one hour of capture. The number of individuals captured or located in roosts, habitat type, elevation and geographic location of sampling site were recorded.
Data analysis
The mist netting sampling effort (m2 nh) was calculated by multiplying the net area (m2) with total duration of sampling (nh) following Kingston (2009). The capture rate of individual species and individual bats were calculated by dividing the total number of species (S) and individual bats (N) by each sampling effort, respectively. Overall species richness of bats considered both roost survey and mist net data, whereas mist net data was used to enumerate diversity and abundance. To quantify the species diversity of bats between different habitats, the Shannon Diversity Index (H′), Dominance (D) and Simpson’s index of diversity were calculated.
The Shannon Diversity Index (H′) is calculated by the following equation (Shannon 1948):
where, ni is the number of individuals in a particular habitat and n is the total number of species.
The dominance index is computed using the formula (Hammer et al. 2001):
where, ni is the number of individuals of taxon i.
Simpson’s index of diversity is computed using the formula (Simpson 1949):
where, n is the total number of individuals of a species and N is the total number of individuals of the species.
The relative abundance was calculated by dividing the number of individuals of a species by the total number of all species. To investigate the species abundance distributions between habitats, a rank abundance plot of bats in the sampled habitats were developed in BioDiversity Professional version 2 (McAleece et al. 1997). To test the significant differences in observed species richness between habitats, Chi square (χ2) test was performed. The t test (t) was carried out to evaluate the Shannon and Simpson diversity indices; and Kolmogorov–Smirnov test was used to assess the significant difference in abundance distributions between habitats. Calculations of diversity indices, χ2 test and t test and preparation of graphs were done using PAST version 3.20 software (Hammer et al. 2001).
To understand the influence of variables on species responses, a total of 26 variables were considered from three correlate types: 21 climate, three physiographic and two disturbance variables. Nineteen climate variables were procured from WorldClim database (http://www.worldclim.org; Hijmans et al. 2005); and two other climate variables (potential evapotranspiration and aridity index) were obtained from CGIAR_CSI database (Trabucco and Zomer 2009). We acquired elevation and aspect from the GMTED 2010 database (https://lta.cr.usgs.gov), and derived the terrain ruggedness index from the differences of the elevation values of adjacent cells relative to the central cell (Riley et al. 2000). Two socio-ecological indicators: the human appropriation of net primary productivity and global human footprint were obtained from WCS_CIESIN database (WCS_CIESIN 2005). Each variable was assigned a common coordinate system of WGS84 datum and conformed to 1 km2 spatial resolution. The selection of environmental variables for this study was done as per Schoeman et al. (2013). Organisation and pre-processing of environmental data layers were done using ArcGIS 10 and R version 3.5. For species with ≥ 20 sample point locations were considered for analysis. We examined the correlation between variables by using the package ‘GGally’ (Schloerke et al. 2014); and a threshold of 0.75 correlation was assigned to select variables to avoid overestimation in model performances owing to high correlation. To evaluate the importance of variables, the principal component analysis, which involved simultaneous centring and scaling was performed by the package ‘factoextra’ (Kassambara and Mundt 2017). We prepared the habitat suitability maps using Maximum Entropy (Maxent) model (Phillips et al. 2006). Maxent was performed using the package ‘dismo’ by setting default parameters (Hijmans et al. 2017). Validation was done by k-fold partitioning with 80% training and 20% test data. We used area under (receiver-operator) curve (AUC) to assess the performances of Maxent. A model above random (AUC > 0.50) was nominated for mapping species distributions.
Results
Species richness and composition
During the study 470 individual bats from 21 species were captured in mist nets. The individual species capture rate was highest (S/m2nh = 0.0047) in moist deciduous forest and lowest (S/m2nh = 0.0029) in semi evergreen forest. Similarly, the capture rate of individual bats was highest (N/m2nh = 0.0345) in moist deciduous forest and lowest (N/m2nh = 0.0128) in both agricultural land and human habitation (Table 1). A total of 23 species of bats (21 from mist net capture) under seven different families were recorded. They comprised of three frugivore species and 20 insectivore species (Table 2). The family Vespertilionidae was well represented by six species, followed by Hipposideridae (5 species), Pteropodidae and Emballonuridae (3 species each), Rhinolophidae, Megadermatidae and Rhinopomatidae (2 species each) (Table 2). Moreover, during the study two species of insectivorous bats: Hipposideros galeritus and Megaderma spasma were reported to occur in Odisha for the first time. Among these, 17 species were recorded from their roosts and foraging sites, four species exclusively from their foraging sites and two species exclusively from their roosting sites (Table 2).
Species diversity and abundance
The Shannon diversity index (H′) was highest in moist deciduous forest (2.540) followed by dry deciduous forest (2.250), agricultural land (2.062), human habitation (2.053) and semi evergreen forest (2.003). The dominance (D) was highest in semi evergreen forest (0.214) followed by dry deciduous forest (0.178), agricultural land (0.161), human habitation (0.160) and moist deciduous forest (0.124). The Simpson index of diversity was recorded highest in moist deciduous forest (0.875) followed by agricultural land and human habitation (0.839 each), dry deciduous forest (0.821) and semi evergreen forest (0.785). However, the diversity between natural and anthropogenic habitats showed statistically significant differences with Shannon index (t test = 4.55, p < 0.05), but statistically insignificant diversity with Simpson index (t test = - 0.49, p > 0.05).
Rhinolophus lepidus was the most abundant species with a relative abundance of 24.68, followed by Cynopterus sphinx (15.11%), Scotophilus heathii (13.19%), Pipistrellus ceylonicus (6.81%), Scotophilus kuhlii (5.96%), Hipposideros lankadiva (4.68%), Pipistrellus tenuis (3.83%), Hipposideros ater (3.62%), Rousettus leschenaultii (3.40%), Megaderma lyra and Taphozous melanopogon (2.98% each), Saccolaimus saccolaimus (1.91%), Hipposideros speoris, M. spasma and Rhinopoma microphyllum (1.70% each), Pipistrellus coromandra (1.06%), Rhinopoma hardwickii (0.85%), Taphozous longimanus (0.64%), Hipposideros fulvus and H. galeritus (1.49% each) and Rhinolophus rouxii (0.21%) (Table 2).
The rank abundance plot exhibited R. lepidus as the most abundant species in moist deciduous forest, dry deciduous forest and semi evergreen forest with the relative abundances of 30.22, 38.10 and 42%, respectively. S. heathii was the most abundant species in agricultural land with relative abundance of 25.51%. C. sphinx and S. heathii were the most abundant species in human habitation (Fig. 2). The Kolmogorov–Smirnov test indicated no significant difference in abundance distribution of bats between habitats (p > 0.05) except when moist deciduous forest was compared with that of semi evergreen forest (Kolmogorov–Smirnov test = 0.5714, p < 0.05) and human habitation (Kolmogorov–Smirnov test = 0.4285, p < 0.05).

Rank abundance plots for bats recorded in five sampled habitats in the Eastern Ghats. Species abbreviations are given in Table 2. (Color figure online)
Co-linearity among variables
After multi co-linearity test, we considered ten least correlated variables for the analysis. This included elevation, global human footprint, human appropriation of net primary productivity, mean diurnal temperature range, mean annual temperature, mean annual precipitation, precipitation of driest quarter, potential evapotranspiration, precipitation seasonality and temperature seasonality (Fig. 3). The mean diurnal temperature range is found to have weak correlation with disturbance variables and mean annual precipitation and temperature. Its correlation with potential evapotranspiration and two seasonal climate variables (precipitation- and temperature- seasonality) was good (r = 0.70). Elevation showed weak and negative correlation with other variables, except in case of precipitation seasonality, where a weak and positive correlation existed. Both disturbance variables are weakly correlated. Similarly, climate variables (except mean diurnal temperature range) are weak, but positively correlated. The mean annual temperature and potential evaporation showed a good correlation (r = 0.70).

Correlation plot of selected variables. DTR mean diurnal temperature range, ELV elevation, HF global human footprint, HNP human appropriation of net primary productivity, MAT mean annual temperature, PDR precipitation of driest quarter, PAN annual precipitation, PET potential evapotranspiration, PS precipitation seasonality, TS temperature seasonality. (Color figure online)
Description of variables
The elevation range for the study area was 23–1672 m. Elevation is found to have maximum variation (277.4 SD). Aspect is low with a mean value of 178.3°. The terrain ruggedness showed variations (163.4 SD); and its value ranged between 6 and 777.2. The aridity index ranged between 0.07 and 0.11, with a mean of 0.09. The standard deviation and standard error was negligible. The mean annual precipitation ranged between 1194 and 1608 mm, with a mean of 1415.7 mm. The precipitations during the coldest and driest quarters were low (34.8 ± 8.7 mm). The precipitation of the driest month was almost dry with 4.7 mm rainfall. The mean precipitation of wettest and warmest quarters match (mean 311 mm) and their value ranged between 230 and 381 mm. The mean precipitation seasonality value was 98.75 CV. It ranged between 87 and 124 CV. The mean diurnal temperature range was 10 °C (8.1–11.3 °C). The mean isothermality value was 42.85. The mean annual temperature ranged between 20.9 °C and 27.5 °C, with the mean at 25.56 °C. Its standard deviation was low at 1.41 SD. The mean annual temperature range was 23.16 °C and ranged between 20.9 °C and 27.5 °C. The minimum and maximum temperatures of the coldest and warmest months were 13.2 °C and 40.8 °C, respectively. Temperature seasonality showed a greater mean (354.9 CV). The mean global human footprint was 30.28, whereas the mean human appropriation of net primary productivity was nearly double this value i.e. 59.7. The standard deviation and standard error were higher in human appropriation of net primary productivity than the global human footprint (Table 3).
Potential distribution
Out of 23 species identified, 11 species were represented with ≥ 20 sample location points. Therefore, predictions were made for 11 species using Maxent. The habitat suitability maps of nine species showing AUC > 0.5 prediction accuracy have been represented (Fig. 4). In general, the north-eastern part is found to be suitable places for chiropterans compared with southern part. C. sphinx is observed to be a well distributed species (Fig. 4a). H. ater is predicted to be distributed in the upper most eastern parts of the region (Fig. 5b). M. lyra is found to occur in upper most western parts of study area, in contrast to occurrences of C. sphinx (Fig. 4c). P. ceylonicus was confined to mid-upper eastern parts of the region (Fig. 4d). P. giganteus is observed to be more prominent in eastern regions, and additionally in southern parts of the study area (Fig. 4e). P. tenuis is predicted to exist in mid to upper eastern parts of the region (Fig. 4f). R. leschenaultii showed a similar habitat preference with that of P. tenuis (Fig. 4g). S. heathii was well distributed over the entire study area. Unlike C. sphinx, its distribution was not suitable in mid-south region of the study site (Fig. 4h). T. melanopogon exhibited a similar distribution structure like that of M. lyra (Fig. 4i).

Potential distribution maps of aCynopterus sphinx, bHipposideros ater, cMegaderma lyra, dPipistrellus ceylonicus, ePteropus giganteus, fPipistrellus tenuis, gRousettus leschenaultii, hScotophilus heathii and iTaphozous melanopogon in eastern parts of the Eastern Ghats. Maps were derived by maximum entropy distribution modelling (Phillips et al. 2006). (Color figure online)

Biplots showing coefficients of variables on principal components aCynopterus sphinx, bHipposideros ater, cMegaderma lyra, dPipistrellus ceylonicus, ePteropus giganteus, fPipistrellus tenuis, gRousettus leschenaultii, hScotophilus heathii and iTaphozous melanopogon. Abbreviations are same as in Fig. 3. (Color figure online)
Significance of variables
The principal component analysis of respective species was evaluated to assess the significance of variables (Fig. 5). In general, elevation was predicted to be a significant determinant for their distributions. Its influence was positive except in case of T. melanopogon, where elevation showed negative effect. The mean annual temperature is found to have negative influence on their distributions, but in case of T. melanopogon, its relationship was positive. Potential evapotranspiration showed a significant effect in their distributions. The impact of this variable is less in T. melanopogon. The precipitation in driest quarter exhibited a negative influence on bat distributions except in case of P. giganteus, where the variable showed a positive influence. The mean annual precipitation is found to have positive influence on distributions of C. sphinx, P. ceylonicus, P. tenuis, R. leschenaultii and T. melanopogon. However, the mean annual precipitation exhibited negative correlation with distributions of H. ater, M. lyra, P. giganteus and S. heathii. The temperature seasonality showed a general positive correlation with bat distributions except in the distribution of P. giganteus, where it showed a negative correlation. The mean diurnal temperature and precipitation seasonality were predicted along the 2nd principal component axis and both exhibited positive correlation with bat distributions.
Discussion
The present study identified 23 bat species existing in the study area. This includes 12 species newly recorded along the Eastern Ghats in Odisha, and two species i.e., H. galeritus and M. spasma newly recorded for the Odisha state. The bat fauna of the hill ranges of Eastern Ghats of Odisha is more than 90% of the total bats of Odisha (Debata et al. 2016); and around 69% of the total bats of the Eastern Ghats (Dutta and Mohapatra 2017). Earlier studies reported 11 bat species in the Eastern Ghats of Odisha i.e., C. sphinx, P. giganteus, R. leschenaultii, R. lepidus, R. rouxii, M. lyra, T. longimanus, T. melanopogon, P. coromandra, P. tenuis and S. heathii (Das et al. 1993; Ramkrishna et al. 2006). The dominant presence of two bat families: Vespertilionidae and Hipposideridae corroborates with the earlier reports from the Western Ghats (Korad et al. 2007), Andaman and Nicobar Islands (Aul et al. 2014; Srinivasulu et al. 2017), and the Himalayas (Saikia et al. 2011). The dominance of both families has also been reported in tropical countries like Borneo (Struebig et al. 2012), China (Luo et al. 2013), Malaysia (Kingston et al. 2003), Thailand (Phommexay et al. 2011) and Vietnam (Furey et al. 2010). The dominant presence of insectivore species is in congruence with the tropical bat communities (Mickleburgh et al. 1992; Hutson et al. 2001).
The frequency of bat capture rate was higher in the natural forest than that in anthropogenic habitats. It could be attributed to greater heterogeneity in natural habitats (moist deciduous forest, dry deciduous forest and semi evergreen forest) owing to multiple layers of stratification providing wide ranges of foraging niches for different species (Struebig et al. 2008; Deshpande 2012). The availability of food resources and availability of preferable roosting sites greatly influence bats and increase their presence in similar landscapes (Kingston 2010; Furey et al. 2010; Phommexay et al. 2011; Struebig et al. 2012). Loss of natural habitats due to human induced activities is a major threat to bats (Mickleburgh et al. 2002). It is in congruence with earlier studies that many bat species from Hipposideridae, Megadermatidae, Emballonuridae and Vespertilionidae families are true forest dwellers, and seldom occur in places of human habitation (Bates and Harrison 1997). However, the relative abundance of C. sphinx, R. leschenaultii and S. heathii was maximum in anthropogenic habitats. The availability of shrubs and bushes in the home gardens and across roadsides are preferred foraging sites for R. lepidus, a forest dwelling species.
R. lepidus, C. sphinx, S. heathii, P. coromandara and P. tenuis are highly abundant in the study area. Maxent predictions also reveal the same, i.e., C. sphinx and S. heathii are well distributed for the region (Fig. 5). These species are common to south Asia (Srinivasulu and Srinivasulu 2012) and in disturbed habitats (Bates and Harrison 1997). The availability of flowering and fruiting trees and agricultural crops attract insect pests, and it may attribute to assemblage of these bats in places of available food resources (Faria et al. 2006). In contrast, S. saccolaimus, H. galeritus, H. lankadiva and T. melanopogon showed less relative abundance indicating that they are habitat specialists preferring forests. T. melanopogon has comparatively less habitat suitability to the region. On the other hand, P. tenuis, R. lepidus, T. melanopogon and M. lyra exhibited similar distribution pattern. It may ascribe to their common feeding habits and climatic dependence. Probably due to the same reason, H. ater and M. lyra have different niche. However, further investigations are necessary to substantiate feeding habits and climatic dependence on their habitat preferences (Fig. 5).
In general, elevation is a significant determinant for chiropteran distributions. It indicates physiography has greater significance in their distributions. The availability of rocks and crevices could provide them to roost safely at elevated surfaces. The availability of forests might support their preferences at elevated regions. The low human interference at higher elevation also helps them grow safe. These findings corroborate with the earlier studies that elevation and habitat characteristics are reported to be significant determinants for chiropteran diversity (Arita 1993; McCain 2007). However, elevation has negative influence on the distribution of T. melanopogon, indicating its dependency on temperature. Unlike other species, the prediction showed its positive correlation with the mean annual temperature. Due to the same reason, potential evapotranspiration, which showed significant effects in distributions of other species, was less effective in the distribution of T. melanopogon. Overall, precipitation in the driest quarter showed a negative association with their species distributions, indicating significance of water availability during driest period. The positive influence of this variable in the distribution of P. giganteus might attribute to its efficiency to manage water stress better compared with other species. The positive influence of mean annual precipitation on distributions of C. sphinx, P. ceylonicus, P. tenuis, R. leschenaultii and T. melanopogon describe their water dependencies. However, the negative effects of the mean annual precipitation exhibited on distributions of H. atar, M. lyra, P. giganteus and S. heathii suggests their greater ability to manage water stress or less dependent on water. A general positive correlation between temperature seasonality and species distributions indicates their less vulnerability to fluctuations in temperature. The negative correlation between temperature seasonality and the distribution of P. giganteus could attribute to its inefficiency to regulate thermal stress due to fluctuations in temperature regime and climate change effects. The mean diurnal temperature and precipitation seasonality are positive determinants of species distributions, attributing to their resilience to seasonal precipitation changes and day time temperature fluctuations.
Conclusions
The study recorded 23 bat species. It includes the first distribution record of two species i.e., H. galeritus and M. spasma for the region. The bat capture rate was the highest in the moist deciduous forest describing higher bat diversity than in the places of human habitation. Use of Maxent could reveal the north eastern part of the hill ranges of Eastern Ghats of Odisha is more suitable for chiropterans. Elevation and potential evapotranspiration are significant determinants for their distributions except in the distributions of T. melanopogon, which showed negative correlation with both. The mean diurnal temperature and precipitation seasonality are positive determinants of bat species distributions, which may be attributed to their resilience to seasonal precipitation changes and day time temperature fluctuations. The negative effect of temperature seasonality on the distribution of P. giganteus could indicate its vulnerability to climate change effects. However, global studies on the impacts of climate change on bats are still at a preliminary stage and need further investigations. Inclusion of H. galeritus and R. rouxii as Nationally Threatened species by the Conservation Assessment and Management Plan of south Asian bats (Molur et al. 2002) is a major concern for conservation. The effects of climate change on the distribution of P. giganteus are also significant from conservation point of view. The present study generated baseline information on bat species distributions along the Eastern Ghats of Odisha; and could provide useful inputs for monitoring the future spatio-temporal changes and long term conservation measures.
References
Adams RA (2010) Bat reproduction declines when conditions mimic climate change projections for western North America. Ecology 91:2437–2445
Adams RA, Hayes MA (2008) Water availability and successful lactation by bats as related to climate change in arid regions of western North America. J Anim Ecol 77:1115–1121
Agarwal I, Wilkinson M, Mohapatra PP, Dutta SK, Giri VB, Gower DJ (2013) The first terestomatan caecilian (Amphibia: Gymnophiona) from the Eastern Ghats of India—a new species of Gegeneophis Peters, 1880. Zootaxa 3696:534–546
Anthony ELP, Stack MH, Kunz TH (1981) Night roosting and the nocturnal time budget of the little brown bat, Myotis lucifugus: effects of reproductive status, prey density, and environmental conditions. Oecologia 51(2):151–156
Arita HT (1993) Conservation biology of the cave bats of Mexico. J Mammal 74:693–702
Aul B, Bates PJJ, Harrison DL, Marimuthu G (2014) Diversity, distribution and status of bats on the Andaman and Nicobar Islands, India. Oryx 48:204–212
Barlow K (1999) Expedition field techniques—bats. Royal Geographical Society, London
Bat Conservation Trust (2007) Bat surveys—good practice guidelines. Bat Conservation Trust, London
Bates PJJ (2013) Bats. In: Johnsingh AJT, Manjrekar N (eds) Mammals of South Asia, vol 1. Universities Press, Hyderabad, pp 68–93
Bates PJJ, Harrison DL (1997) Bats of the Indian sub-continent. Harrison Zoological Museum Publication, Sevenoaks
Boonman AM, Boonman M, Bretschneider F, van de Grind WA (1998) Prey detection in trawling insectivorous bats: duckweed affects hunting behaviour in Daubenton’s bat, Myotisdaubentonii. Behav Ecol Sociobiol 44:99–107
Burles DW, Brigham RM, Ring RA, Reimchen TE (2009) Influence of weather on two insectivorous bats in a temperate Pacific Northwest rainforest. Can J Zool 87(2):132–138
Champion HG, Seth SK (1968) A revised study of the forest types of India. Government of India Press, New Delhi
Ciechanowski M, Zajac T, Bilas A, Dunajski R (2007) Spatiotemporal variation in activity of bat species differing in hunting tactics: effects of weather, moonlight, food abundance, and structural clutter. Can J Zool 85(12):1249–1263
Crichton EG, Krutzsch PH (2000) Reproductive biology of bats. Academic Press, London
Das PK, Lal JP, Agrawal VC (1993) Mammalia. In: Alfred B (ed) State Fauna series 1: Fauna of Orissa, Part 4. Zoological Survey of India, Calcutta, pp 143–180
Debata S, Palita SK, Behera S (2016) Bats of Odisha—a pictorial handbook. Odisha Biodiversity Board, Bhubaneswar
Deshpande K (2012) Assessing diversity and distribution of bats in relation to land use and anthropogenic threats in the southern Western Ghats, India. Rufford Small Grant Foundation, London. https://www.rufford.org/rsg/projects/kadambari_deshpande
Dunbar MB, Tomasi TE (2006) Arousal patterns, metabolic rate, and an energy budget of eastern red bats (Lasiurus borealis) in winter. J Mammal 87:1096–1102
Dutta SK, Mohapatra PP (2017) Eastern Ghats: Faunal composition and conservation. In: Pratihar S, Clark HO Jr (eds) Defaunation and conservation. Tucson Herpetological Society, Tucson, pp 149–183
Elith J, Graham CH, Anderson RP, Dudík M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, McC Overton J, Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberón J, Williams S, Wisz MS, Zimmermann NE (2006) Novel methods improve prediction of species’ distribution from occurrence data. Ecography 29:129–151
Faria D, Laps RR, Baumgarten J, Cetra M (2006) Bat and bird assemblages from forests and shade cacao plantations in two contrasting landscapes in the Atlantic forest of southern Bahia, Brazil. Biodivers Conserv 15:587–612
Fenton MB, Simmons NB (2014) Bats: a world of science and mystery. University of Chicago Press, Chicago
Francis AP, Currie DJ (2003) A globally consistent richnessclimate relationship for angiosperms. Am Nat 161:523–536
Frick WF, Reynolds DS, Kunz TH (2009) Influence of climate and reproductive timing on demography of little brown myotis Myoti slucifugus. J Anim Ecol 79(1):128–136
Furey NM, Mackie IJ, Racey PA (2010) Bat diversity in Vietnamese limestone karst areas and the implications of forest degradation. Biodivers Conserv 19:1821–1838
Guthrie G (2012) Basic research methods. SAGE Publications India Pvt Ltd, New Delhi
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electronica 4:1–9
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25(15):1965–1978
Hijmans RJ, Phillips S, Leathwick J, Elith J, Hijmans MRJ (2017) Package ‘dismo’. Circles 9:1
Hirzel AH, Le Lay G, Helfer V, Randin C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 199:142–152
Humphrey P, Bridge D, Lovejoy TE (1968) A technique for mist netting in the forest canopy. Bird Banding 39:43–50
Hutson AM, Mickleburgh SP, Racey PA (2001) Microchiropteran bats: global status survey and conservation action plan. IUCN/SSC Chiroptera Specialist Group, IUCN, Gland
Jones G, Jacobs DS, Kunz TH et al (2009) Carpe noctem: the importance of bats as bioindicators. Endanger Species Res 8:93–115
Kassambara A, Mundt F (2017) Factoextra: extract and visualize the results of multivariate data analyses. R package version 1.0.4
Kent R, Bar-Massada A, Carmel Y (2014) Bird and mammal species composition in distinct geographic regions and their relationships with environmental factors across multiple spatial scales. Ecol Evol 4:1963–1971
Kingston T (2009) Analysis of species diversity of bat assemblages. In: Kunz TH, Parsons S (eds) Ecological and behavioural methods for the study of bats, 2nd edn. Johns Hopkins University Press, Washington, pp 195–215
Kingston T (2010) Research priorities for bat conservation in Southeast Asia: a consensus approach. Biodivers Conserv 19:471–484
Kingston T, Francis CM, Zubaid A, Kunz TH (2003) Species richness in an insectivorous bat assemblage from Malaysia. J Trop Ecol 19:67–79
Korad V, Yardi K, Raut R (2007) Diversity and distribution of bats in the Western Ghats of India. Zoos Print J 22(7):1752–2758
Kunz TH (1988) Ecological and behavioural methods for the study of bats. Smithsonian Institution Press, Washington
Kunz TH, de Torrez EB, Bauer D, Lobova T, Fleming TH (2011) Ecosystem services provided by bats. Ann N Y Acad Sci 1223:1–38
Lane DJW, Kingston T, Lee BPY-H (2006) Dramatic decline in bat species richness in Singapore, with implications for Southeast Asia. Biol Conserv 131:584–593
Luo J, Jiang T, Lu G, Wang L, Wang J, Feng J (2013) Bat conservation in China: should protection of subterranean habitats be a priority? Oryx 47:526–531
McAleece N, Gage JD, Lambshead J, Patterson GLJ (1997) Biodiversity professional. Jointly developed by the Scottish Association for Marine Science and the Natural History Museum London. http://www.sams.ac.uk/science/outputs
McCain CM (2007) Could temperature and water availability drive elevational species richness patterns? A global case study for bats. Glob Ecol Biogeogr 16:1–13
Mello MAR, Kalko EKV, Silva WR (2009) Ambient temperature is more important than food availability in explaining reproductive timing of the bat Sturniralilium (Mammalia: Chiroptera) in a montane Atlantic Forest. Can J Zool 87:239–245
Mickleburgh SP, Hutson AM, Racey PA (1992) Old world fruit bats. An action plan for their conservation. International Union for Conservation of Nature and Natural Resources, Chiroptera Specialist Group (IUCN/SSC), Gland
Mickleburgh SP, Hutson AM, Racey PA (2002) A review of the global conservation status of bats. Oryx 36:18–34
Mohapatra PP, Das A, Dutta SK (2010) Psammodynastes pulverulentus, range extension from Eastern Ghats, Andhra Pradesh. Herpetol Rev 41(1):111
Mohapatra PP, Palei HS, Hussain SA (2014) Occurrence of Asian small-clawed otter Aonyx cinereus (Illiger, 1815) in Eastern India. Curr Sci 107:367–370
Molur S, Marimuthu G, Srinivasulu C et al (2002) Status of South Asian Chiroptera: Conservation Assessment and Management Plan (C.A.M.P.) Zoo Outreach Organisation, Conservation Breeding Specialist Group South Asia, and Wildlife Information and Liaison Development Society, Coimbatore
Moreno CE, Halffter G (2001) Spatial and temporal analysis of α, β and γ diversities of bats in a fragmented landscape. Biodivers Conserv 10:367–382
Murray KA, Retallick RWR, Puschendorf R, Skerratt LF, Rosauer D, McCallum HI, Berger L, Speare R, VanDerWal J (2011) Assessing spatial patterns of disease risk to biodiversity: implications for the management of the amphibian pathogen, Batrachochytrium dendrobatidis. J Appl Ecol 48:163–173
Nayak AK, Nair MV, Mohapatra PP (2014) Stripe-necked Mongoose Herpestesvitticollis in Odisha, eastern India: a biogeographically significant record. Small Carniv Conserv 51:71–73
Newson SE, Mendes S, Crick HQP, Dulvy NK, Houghton JDR, Hays GC, Hutson AM, Macleod CD, Pierce GJ, Robinson RA (2008) Indicators of the impact of climate change on migratory species. Endanger Species Res 7:101–113
Paliwal A, Mathur VB (2012) Predicting potential distribution of Gaur (Bosgaurus) in Tadoba-Andhari Tiger Reserve, Central India. J Life Sci 6:1041–1049
Park KJ, Jones G, Ransome RD (2000) Torpor, arousal and activity of hibernating greater horseshoe bats (Rhinolophus ferrumequinum). Funct Ecol 14:580–588
Phillips SJ, Dudίk M (2008) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161–175
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190(3–4):231–259
Phommexay P, Satasook C, Bates PJJ, Pearch M, Bumrungsri S (2011) The impact of rubber plantations on the diversity and activity of understorey insectivorous bats in southern Thailand. Biodivers Conserv 20:1441–1456
Purohit S, Nair MV, Palita SK (2017) On the occurrence of Black BazaAvicedaleuphotes Dumont, 1820 (Aves: Falconiformes: Accipitridae) in the Gupteswar forests of the Eastern Ghats, Odisha, India. J Threat Taxa 9:10964–10967
Ramkrishna Siddique SZ, Sethy P, Dash S (2006) Faunal resources of Similipal Biosphere Reserve, Mayurbhanj, Orissa. Zoological Survey of India, Kolkata
Ray GC (2005) Geography of Orissa. Kitab Mahal, Cuttack
Razgour O, Rebelo H, Febbraro MD, Russo D (2016) Painting maps with bats: species distribution modelling in bat research and conservation. Hystrix Ital J Mamm. https://doi.org/10.4404/hystrix-27.1-11753
Reddy CS, Jha CS, Dadhwal VK (2014) Spatial dynamics of deforestation and forest fragmentation (1930-2013) in Eastern Ghats, India. Int Arch Photogram Remote Sens Spat Inf Sci 40(8):637–644
Richter HV, Cumming GS (2008) First application of satellite telemetry to track African straw-coloured fruit bat migration. J Zool 275:172–176
Riley J, Hoppa GV, Greenberg R, Tufts BR, Geissler P (2000) Distribution of chaotic terrain on Europa. J Geophys Res Planets 105(E9):22599–22615
Saikia U, Thakur ML, Bawri M, Bhattacherjee PC (2011) An inventory of the chiropteran fauna of Himachal Pradesh, northwestern India with some ecological observations. J Threat Taxa 3:1637–1655
Saranya KRL, Reddy CS (2016) Long term changes in forest cover and land use of Similipal Biosphere Reserve of India using satellite remote sensing data. J Earth Syst Sci 125(3):559–569
Schloerke B, Crowley J, Cook D, Hofmann H, Wickham H, Briatte F, Marbach M, Thoen E (2014) Ggally: extension to ggplot2. R package version 0.5. 0
Schoeman MC, Cotterill FPD, Taylor PJ, Monadjem A (2013) Using potential distributions to explore environmental correlates of bat species richness in southern Africa: effects of model selection and taxonomy. Curr Zool 59:279–293
Shannon C (1948) A mathematical theory of communication. Bell Syst Tech J 27(3):379–423
Sherwin HA, Montgomery WI, Lundy MG (2013) The impact and implications of climate change for bats. Mammal Rev 43:171–182
Simpson EH (1949) Measurement of diversity. Nature 163:688
Srinivasulu C, Srinivasulu B (2012) South Asian mammals—their diversity, distribution, and status. Springer, New York
Srinivasulu C, Racey PA, Mistry S (2010) A key to the bats (Mammalia: Chiroptera) of South Asia. J Threat Taxa 2:1001–1076
Srinivasulu C, Srinivasulu A, Srinivasulu B, Gopi A, Dar TH, Bates PJJ, Rossiter SJ, Jones G (2017) Recent surveys of bats from the Andaman Islands, India: diversity, distribution, and echolocation characteristics. Acta Chiropterol 19:419–437
Struebig MJ, Kingston T, Zubaid A, Modh-Adnan A, Rossiter SJ (2008) Conservation value of forest fragments to Paleotropical bats. Biol Conserv 141:2112–2126
Struebig MJ, Bozek M, Hildebrand J, Rossiter SJ, Lane DWJ (2012) Bat diversity in the lowland forests of the heart of Borneo. Biodivers Conserv 21:3711–3727
Trabucco A, Zomer RJ (2009) Global aridity index (global-aridity) and global potential evapo-transpiration (global-PET) geospatial database. CGIAR Consortium for Spatial Information, Montpellier
Voigt CC, Lewanzik D (2011) Trapped in the darkness of the night: thermal and energetic constraints of daylight flight in bats. Proc R Soc Lond B 278:2311–2317
Webb PI, Speakman JR, Racey PA (1995) Evaporativewater loss in two sympatric species of vespertilionid bat, Plecotusauritus and Myotisdaubentoni: relation to foraging mode and implications for roost site selection. J Zool 235:269–278
Wildlife Conservation Society—WCS, and Center for International Earth Science Information Network—CIESIN—Columbia University (2005) Last of the wild project, version 2, 2005 (LWP-2): global human footprint dataset (IGHP). NASA Socioeconomic Data and Applications Center (SEDAC), Palisades. https://doi.org/10.7927/h4gf0rfq
Wisz MS, Hijmans RJ, Li J, Peterson AT, Graham CH, Guisan A, Elith J, Dudík RPM, Ferrier S, Huettmann F, Leathwick JR, Lohmann L, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton JM, Phillips SJ, Richardson KS, Scachetti-Pereira R, Schapire RE, Soberón J, Williams SE, Zimmermann NE (2008) Effects of sample size on the performance of species distribution models. Divers Distrib 14:763–773
Xu C, Huang ZYX, Chen BJW, Zhang M, Liu M (2014) Can local landscape attributes explain species richness patterns at macroecological scales? Glob Ecol Biogeogr 23:436–445
Acknowledgements
The study was carried out with financial support from University Grants Commission, India to the first author. Authors would like to thank the Principal Chief Conservator of Forests and Chief Wildlife Warden, Odisha for the required study permission (permission No. 396/4WL). We acknowledge the State Forest Department staffs and local people from different regions of Odisha for their help and support during the field study. We would like to thank M.D. Behera for his suggestions in manuscript preparation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by David Hawksworth.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Debata, S., Panda, R.M. & Palita, S.K. Chiropteran diversity and the key determinants of their distribution in Eastern Ghats, India. Biodivers Conserv 28, 2385–2404 (2019). https://doi.org/10.1007/s10531-019-01715-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-019-01715-w