
Abstract
Ancient forests are of considerable interest for strategies for biodiversity conservation. However, in European forest landscapes fragmented and harvested for a long time forest continuity might be no longer a key driver for flying organisms such as saproxylic beetles. In a study based on paired samples (n = 60 stands, p = 180 traps) of ancient and recent forests, we investigated the effects of forest continuity on saproxylic beetle assemblages in two French regions. Mean species richness was significantly related with deadwood volume in ancient forests, but not in recent forests. This loss of relationship between assemblages and their environment suggests that dispersal limitation is at work, at least for some species. Forest continuity had a significant effect on mean species richness and on the mean number of common species, but not on rare species. Forest continuity had a significant effect on assemblage composition in one out of the four cases tested. In both regions, we identified species associated with either recent or ancient forests. Finally, mean body size of species was significantly smaller in recent forests compared with ancient ones, as was their tree diameter preference, despite a higher volume of large deadwood in recent forests. These results lend support to using forest continuity as a criterion to identify sites of conservation importance, even in highly fragmented landscapes.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Ancient forests are considered to be “hotspots” or refuges in many strategies for biodiversity conservation (Goldberg et al. 2007; Hermy and Verheyen 2007; Lachat and Butler 2009; Patru-Stupariu et al. 2013). Here is important to point out that the concept of ancient forest refers only to the temporal continuity of the forest cover regardless of human disturbances (Cateau et al. 2015). Given their continuous tree cover for several centuries (i.e. forest continuity), ancient forests may harbor higher levels of biodiversity, or at least specific assemblages (Cateau et al. 2015). This has been well documented for vascular plants, and to lesser extent for other groups such as bryophytes or invertebrates (Norden et al. 2014). For deadwood-dependant (named after “saproxylic”) species, continuity of deadwood supply is assumed to be more important for species persistence than current deadwood amounts (Similä et al. 2003; Sverdrup-Thygeson et al. 2014). Though some saproxylic species have been suggested as bio-indicators of ecological continuity (Alexander, 2004; Müller et al. 2005; Norden and Appelqvist 2001), this assumption has not been tested. Indeed, there are very few references concerning the effect of continuity per se on saproxylic beetles (Bouget et al. 2014a; Buse 2011; Gossner et al. 2008; Horák et al. 2013; Irmler et al. 2010). Most of the times, the effects of continuity are confounded with the effects of habitat quality (i.e. old-growth attributes for saproxylic species) (Grove 2002; Siitonen and Saaristo 2000; Nilsson and Baranowski 1997).
Studying the effects of continuity is challenging since the effects of dispersal limitation (continuity effects per se) must be disentangled from habitat limitations (Norden et al. 2014). In recent forests (i.e. disturbed habitats), if dispersal limitation is still at work, the community is dominated by good dispersers. The relationship between habitat attributes and assemblages might then be weakened as only a fraction of the community is present. This pattern has been observed for plants by Vellend et al. (2007). For saproxylic species the prediction could be translate into the loss of the relationship between deadwood amount and species richness for example.
In the two existing studies on the effects of forest continuity on saproxylic beetle assemblages, habitat quality (i.e. deadwood amount or diversity) was either not estimated (Irmler et al. 2010) or investigations were restricted to young stands in recent forests adjacent to ancient forests (Gossner et al. 2008). Understanding the importance of continuity for the occurrence of saproxylic beetles remains a research challenge (Norden et al. 2014; Stokland et al. 2012; Sverdrup-Thygeson et al. 2014) with implication in biodiversity conservation policies.
The long history of forest fragmentation and exploitation in most European landscapes offers the opportunity to explore the effects of continuity per se and to challenge the concept of “ancient forest” as a relevant criterion for saproxylic species conservation policies. Indeed, in those landscapes, less mobile and more specialized species have declined due to fragmentation and exploitation (Vandekerkhove et al. 2011). And, due to exploitation, most of ancient forests are not mature or old-growth forests (Cateau et al. 2015). Despite evidence for a time lag between habitat modification and species extirpation, Kuussaari et al. (2009) suggested that the extinction debt may have already been paid off in landscapes with small, isolated patches. For patches of small size and isolated for a long time, forest continuity might be no longer a key driver for saproxylic beetles assemblages. The concept of maturity that refers to the stage of natural development of a stand (Cateau et al. 2015) would then be more interesting for biodiversity conservation policies in highly and long-standing fragmented landscapes.
Using species traits to characterize communities provides interesting insights into the processes that are at work (Gossner et al. 2013; Pausas and Verdú 2010). This approach bypasses the analytical problems related to a single-species approach, which suffers from insufficient data for modeling. Preferences for deadwood diameter and decay stages are two interesting life traits to take into account when investigating the effects of forest continuity on saproxylic beetle assemblages. Indeed, following the habitat templet hypothesis (Southwood 1977), species associated with long-lasting microhabitats (i.e. large logs and/or late decay stages) are likely to be poor dispersers. Whereas species associated with ephemeral habitats (branches and/or early decay stages) should be better dispersers and therefore less affected by local continuity (Norden et al. 2014). For saproxylic species, Nilsson and Baranowski (1997) provided some evidence for a lower dispersal ability of species living in stable microhabitat (hollow trees) compare with species living in more ephemeral microhabitat (standing or lying dead trees). Southwood (1977) also highlighted the importance of spatial heterogeneity of habitat distribution in shaping the ecological strategies of species. A negative effect of the reduction of habitat availability on dispersal rate has been showed by simulation work (Travis and Dytham 1999). Species inhabiting small-diameter deadwood are generally the smallest in size (Brin et al. 2011). As the diameter preference of most saproxylic beetle species is not precisely known, body size can be used as a proxy for this preference (Seibold et al. 2015). Body size may also be a predictor of fragmentation sensitivity (Henle et al. 2004). Lower abundance and rate of recovery of larger species make them more prone to extinction following fragmentation (Cotgreave 1993; Lawton 1994). But this tendency may be offset by the negative correlation and population fluctuation (Pimm 1991).
Assemblages can be characterized not only by the mean value of several traits but also by the degree of similarity among species regarding each trait (Pausas and Verdú 2010). By comparing the observed distribution of species trait values to the distribution of trait values in the available species pool, we can detect patterns of over-dispersion, random or clustered trait values suggesting respectively three mechanisms: competition, random or habitat filtering effects (Pausas and Verdú 2010). For clustering patterns, the deviation from random community composition can help evaluate the importance of habitat filtering effect. In line with the hypothesis of weaker species-environment relationships in recent forests, we would expect a lower, if any, habitat filtering effect in recent than in ancient forests.
Landscape connectivity or landscape continuity may compensate for local discontinuity (Inclán et al. 2014; Norden et al. 2014; Rösch et al. 2013). Indeed, mobile species like flying insects easily recolonize new habitat patches as soon as they become suitable. It is therefore important to control for landscape connectivity when we want to compare ancient and recent forests. Using a sampling design with paired samples of ancient and recent forests replicated in two regions, we investigated the effects of forest continuity on assemblages of flying saproxylic beetles. More specifically, we tested the following hypotheses:
-
Due to the weakening of the species-environment relationships caused by dispersal limitation local amount of deadwood should only be a good predictor of species richness in ancient forests.
-
Forest continuity should have an effect on assemblage composition due to differences in colonization ability of species.
-
In recent forests compared with ancient forests, species should be smaller and mostly associated with small-diameter deadwood and early decay stages.
-
Due to weaker species-environment relationships in recent forests, the habitat filtering effect, if any, should be less important in those forests.
Materials and methods
Study areas and sampling design
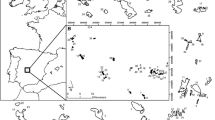
Fragmented woodlots were selected in two 30 × 40 km agricultural landscapes, one located in Northern France (Gatinais-Puisaye, names hereafter “Gatinais”) and one in South-Western France (Gascogne) (Fig. 1). In each region, we selected 15 pairs of woodlots (mean area of 3 ha), separated by a mean distance of 1000 m. Sites were selected in a twostep process: by using GIS we first identified paired sites made of 1 recent and 1 ancient forest, similar in size (3 ha on average) and not too convoluted in shape, with a maximum distance of 1.5 km between them. We then checked for the tree species composition by simple visual inspection of each stand in the field so as to include only oak-dominated stands in our samples as this is the predominant forest formation in our study regions. We also limited variations in topography, soil conditions and mean stand age in a very narrow range, and excluded the recently harvested woodlots. The first step gave us about 20 pairs of sites in Gascogne, and 29 pairs of sites in Gatinais. We stopped the second step once we had reached our goal of 15 pairs of site within each region.

Location of study regions and study sites (filled black triangle: ancient forests, +: recent forests)
Ancient forests are those present on the “Map on France”, a military map drawn in the middle of the nineteenth century (Dupouey et al. 2007) at the moment when forest cover in France reached its lowest level in the last two millennia. By definition, ancient forests are those which have been established for at least 200 years, i.e. their existence was already attested in the middle of the nineteenth century on state maps. Recent forests were identified from digitalized maps based on aerial photographs taken in 2010 by the National Institute of Geographic and Forest Information (© IGN-BD Topo® edition 2010) and defined as those created by natural succession on former agricultural land, i.e. these sites had not yet been transformed into forests in the middle of the nineteenth century.
All the sites are managed, mainly by coppicing with standards with fine-scaled logging events (<1 ha) (De Warnaffe et al. 2006; Andrieu et al. 2011).
Beetle sampling
Flying saproxylic beetles were captured in alcohol-baited cross-pane flight interception traps (polytrap™). Three of such traps were set out in each woodland at least 30 m apart. The traps were suspended roughly 1.5 m above the ground, and specimens were collected from April to July in 2012.
All saproxylic beetles were identified to the species level following the nomenclature of the Fauna Europaea Web Service (de Jong et al. 2014). They were assigned to a trophic group according to the nomenclature proposed by Bouget et al. (2005).
Commonness and rarity were defined using the species patrimoniality values (= IP) defined in Bouget et al. (2010) with a four-level scale. The geographic scale under consideration for distribution is the French metropolitan territory. As we had only one “very rare” species, we used a simplified classification. The category “common” species combines the classes IP1 (common and widely distributed species) and IP2 (not abundant but widely distributed species, or only locally abundant species). Species that are not abundant and only locally distributed (IP3) and those known in less than 5 localities or in a single “county” in France (IP4) were classified as “rare” species.
Environmental data
Deadwood was measured around each trap during spring 2012. Downed woody debris was quantified using the Line Intersect Sampling (LIS) method (De Vries 1973; Marshall et al. 2000). Pieces with a diameter of at least 2.5 cm were assessed along two perpendicular transects of 50 m each.
All snags and dead trees with a diameter at breast height (dbh) greater than 7.5 cm were measured in a circular plot with a 20 m radius (giving a 1256 m2 area). We only measured the dbh for trees and snags higher than 4 m; for shorter trees, the diameter was taken at mid height.
Stumps were quantified in three circular sub-plots whose radii varied depending on stump’s diameter: 6-m-radius plots (giving a 113 m2 area) for stumps with diameters from 7.5 to 12.5 cm, 10-m-radius plots (giving a 314 m2 area) for stumps with a diameter between 12.5 and 22.5 cm, and 20-m-radius plots (giving a 1256 m2 area) for stumps with diameters greater than 22.5 cm. We followed the same sampling procedure for living trees but added a fourth circular sub-plot (radius = 4 m for 50 m2) for trees with a dbh from 2.5 to 7.5 cm.
For stumps, snags and dead trees with a height of up to 4 m, the volume was calculated with the cylinder formula. For snags and dead trees taller than 4 m, the volume was assessed according to wood volume tables based on the dbh.
Four variables were used to describe deadwood: tree species, diameter (six classes: 5, 10–15, 20–25, 30–40, 50–65, >70 cm), position (lying on the ground, standing, stump), and decay stage. We adapted nine decay stages classes adapted from Sippola et al. (1998) and Larjavaara and Muller-Landau (2010) by crossing three classes of remaining bark cover (from 95 % of the stem still covered by attached bark to missing bark from the whole stem) with three classes of inner wood hardness assessed by ‘knife penetration test’ (from hard outer wood to deeply disintegrated and soft inner wood). We then calculated a deadwood diversity index to reflect the number of observed deadwood types, that is the number of combinations of the above four variables (tree species × diameter class × decay stage × position), as suggested by Siitonen et al. (2000).
We used landscape forest cover (LFC) as a measure of connectivity. The LFC was calculated in a circular buffer zone centered at the barycenter of the three traps with the ArcGIS 10.2 for Desktop ® software ver. 2012 [Esri (Environmental Systems Research Institute), Redlands, CA] and French vegetation maps (© IGN-BD Topo® edition 2010) at two spatial scales: 78 ha (radius 500 m) and 1256 ha (radius 2 000 m). These spatial scales were considered adequate to differentiate beetle species with different dispersal abilities (ranging from ten to several hundreds of meters; e.g. Sauvard 2004; Ranius 2006) and to demonstrate possible isolation effects. Using digitalized versions of historic maps from the nineteenth century, we also computed the ancient landscape forest cover (ALFC) at the same scales (see “Study sites and sampling design” for more details about the historic maps).
Data analysis
For all analyses, we pooled the data from the three traps within each forest site into one “observational unit”. For deadwood attributes, we calculated the mean for each observational unit.
To test for effects on species richness of the local deadwood resource (deadwood volume and deadwood tree species diversity), and for forest continuity (current vs ancient), we fitted generalized linear mixed Poisson models. We introduced an interaction term between deadwood volume and forest continuity to test our first hypothesis about the weakening of species-environment relationships in recent forests. Since deadwood volume and deadwood diversity were highly correlated (r = 0.78, P < 0.0001), we only used the volume variable in our models. Site and region were random factors. LFCs—current and past- were also introduced into the models as covariates to account for differences in landscape connectivity. Total species richness, number of rare species and number of common species were dependent variables.
Due to differences in regional species pools, we explored the individual responses of species at the regional scale. We considered beetle species occurring in at least 10 % of the sites and with a total count of more than 25 individuals. We fitted generalized linear mixed models with either a Poisson distribution or a binomial, for abundance and occurrence respectively. We used the volume of deadwood, the forest continuity and the LFCs as fixed factors and the site and region as random factors.
To test for an effect of forest continuity on beetle assemblage composition, we used the multivariate generalization of generalized linear models proposed by Warton (2011). This method consists in fitting separate models to each species, summing the test statistics and then assessing the significance of these multivariate test statistics through a resampling procedure. Compared with distance-based multivariate analyses or canonical correspondence analyses, this approach is more appropriate for mean–variance relationships, which are critical in count data (Warton et al. 2012). We fitted the GLMs with a negative binomial distribution where mean abundance was the dependent variable and forest continuity the predictor. To calculate the multivariate test statistics, we relaxed the assumption of independence for the species response variables. We used the parametric bootstrap resampling method to estimate the P value associated with the test statistics (n = 1000).
To characterize beetle assemblages, we computed two metrics for each trait (body size, decay-class preferences and diameter preferences): the community-level weighted mean (CWM) and the effect size of the mean pairwise distances. The former indicates the average of the niche positions and the latter indicates the range extent or the dispersion around the average. For categorical traits (decay and diameter classes), we estimated the niche position of each species following Gossner et al. (2013). These authors used the occurrence of a given species across categories and weighting scores (0.5 = very rarely used; 1 = rarely used; 2 = commonly used; 3 = preferred) to end up with a niche position value. Effect size of the mean pairwise distances for each trait was estimated by randomizing (n = 999) the community data matrix within samples. The species richness of samples was maintained in the null models. Values > 0 indicate over-dispersion, possibly due to competition, while values < 0 indicate clustering due to habitat filtering (Pausas and Verdú 2010). We used the data provided by Gossner et al. (2013) and experts for the decay and diameter preferences. We have to acknowledge that data were not available for all the species. However, there were no bias in the distribution of species with missing data (i.e. 31 and 29 % in ancient and recent forests respectively). Body size information come from the French Saproxylic Database FRISBEE (Bouget et al. 2010).
To test for a forest continuity effect on each trait metric, we used linear mixed-effect models with the volume of deadwood as co-variable (fixed factor) to account for the quality of the habitat. Region and pair were used as random factors to account for the spatial structure of our data set.
All analyses were carried out with the R 3.1.0 (R Core Team 2013) software, using the picante (Kembel et al. 2014) and FD (Laliberté et al. 2014) packages for trait analyses, the lme4 (Bates et al. 2014) and multcomp (Hothorn et al. 2014) packages for the GLMMs, and the mvabund package (Wang et al. 2015) for multivariate analyses.
Results
Recent forests had a higher volume of deadwood on average (Table 1). This characteristic allowed us to disentangle the habitat limitation effect from the effect of forest continuity per se. Landscape Forest Cover was not significantly different between ancient and recent forests; this was true at both spatial scales (Table 1). This confirms the ability of our sampling design to control for this factor. Density and basal area of living trees were also not significantly different between the two forest continuity categories (Table 1). This suggests that our comparisons are not biased by microclimatic differences.
A total of 492 saproxylic species were trapped and identified: 425 and 310 species in the Gascogne region and the Gatinais region, respectively (see Supplementary Appendix for the species list). Of these species, 208 (42 %) were observed in both regions. None of the species are threatened according to the European Red List of saproxylic beetles (Nieto and Alexander 2004).
Forest continuity had a significant effect on mean species richness and on the mean number of common species, but no effect on rare species (Table 2). The mean global species richness, as well as common and rare species richness, were all significantly influenced by the local deadwood volume (Table 2). In line with our first hypothesis, there was significant interaction between deadwood volume and forest continuity, both for the global mean number of species, and for common and rare species richness (Table 2). When we fitted a GLMM for each forest continuity category (ancient vs recent) separately, deadwood volume was still significantly related to mean species richness in ancient forests (z value = 2.389, P = 0.017), though not in recent forests (z value = −1.057, P = 0.291).
In line with our second hypothesis, forest continuity had a significant effect on assemblage composition in Gascogne (Wald test = 24.73, P = 0.002), but not in Gatinais (Wald test = 17.46, P = 0.352) and as far as species abundance is concerned. No significant effect was detected in either region when considering species occurrence (Gascogne: Wald test = 21.53, P = 0.189; Gatinais: Wald test = 11.7, P = 1).
We found almost twice as many species associated with ancient forests (21) than with recent ones (11) (Table 3). Forest continuity had a significant effect either on species abundance or occurrence or sometimes on both (Table 3). In Gascogne and Gatinais respectively, the species associated with ancient forests represent 12.4 and 6.7 % of the testable species (i.e. which occurred in more than 6 sites and were represented by at least 25 individuals).
In line with our third hypothesis, the species observed in recent forests had a significantly smaller mean body size than the species observed in ancient forests (Table 4). Their mean diameter preference was also significantly smaller. For both forest continuity categories, diversity of the three traits (body size, diameter and decay preferences) were globally smaller than expected by chance suggesting habitat filtering effect in all habitat types (Fig. 2). Only the diversity of body size was significantly influenced by forest continuity (Table 4). Species tended to be more similar in terms of body size in recent than in ancient forests despite a higher diversity of deadwood in recent forests.

Diversity of each trait (diameter preference (a), decay preference (b), body size (c)) for each forest continuity class (A: ancient versus R: recent). Diversity was computed as the standardized effect size (n = 999 runs) of mean pairwise distances for each trait
Discussion
Given our study design and the a posteriori control for deadwood amount in our analysis, our results provide strong evidence that a forest continuity effect on flying saproxylic beetle assemblages does indeed exist, in line with results from the few previous studies (Gossner et al. 2008; Irmler et al. 2010).
Species richness was significantly influenced by deadwood volume in ancient forests, but not in recent ones. This loss of relationship between assemblages and their environment is congruent with the pattern observed for plants by Vellend et al. (2007) and corroborates the dispersal limitation hypothesis, for some species at least. In a companion paper (Bouget et al. 2014a), we found a micro-evolutive response that also corroborates this hypothesis: wing loading (used as a proxy for dispersion capacity) was lower in recent than in ancient forest plots for Melandrya barbata males. This suggests that only the best dispersers were able to colonize the recent forest plots. In line with the dispersal limitation hypothesis, the species occurring in recent forests in the present study were on average smaller and had smaller diameter preferences than those occurring in ancient forests. They also tended to be of similar body size. This lack of large species in recent forests even though there are more deadwood and especially large deadwood (Table 1) can be interpreted in light of the habitat templet hypothesis (Southwood 1977). Indeed, the largest species breed on large diameter pieces (Brin et al. 2011) which decayed more slowly (Brown et al. 1996; Vanderwel et al. 2006). Large species might therefore be less mobile than the smallest ones, and they might be among the latest to colonize a new habitat. Furthermore, because of higher population levels and faster reproductive cycles, smaller species might be less prone to extinction in fragmented landscapes (Henle et al. 2004; Stokland et al. 2012). The ability to colonize a new patch depends not only on the dispersal capacity, but also on the number of dispersing individuals (Hanski 1999). In the studied fragmented landscapes, low level of populations of the largest species may hamper their colonization rate of new patch. Species observed in ancient forests but missing in recent woodlots may relate to what is called the “species credit” (Hanski 2000), or the “colonization credit” (Jackson and Sax 2010; Nagelkerke 2010; Nagelkerke et al. 2002). These concepts refer to species that either come back after being regionally extinct or increase their populations following an improvement of habitat quality. Our results provide evidence for such transient dynamics in highly fragmented forests.
Contrary to Bouget et al. (2014b), we did not find any significant effect of deadwood tree species diversity on beetle species richness, even though this is one of the four parameters commonly used to describe deadwood diversity. Deadwood tree species diversity has been acknowledged as a key driver of saproxylic assemblages (Lassauce et al. 2012; Stokland et al. 2012). Our results may be due to the phylogenetic proximity of the tree species in our study. Indeed, at our sites, the deadwood originated from deciduous trees only. Whereas insects assemblages associated with broadleaved trees are clearly different from assemblages associated with conifers trees, patterns are far less obvious within each branch (Brändle and Brandl 2006; Gossner et al. 2009; Milberg et al. 2014).
Forest continuity had a significant effect on assemblage composition with some beetle species clearly more abundant and/or frequent in ancient forests. Our results are congruent with the few references available about saproxylic beetles and forest continuity. Two species, Dryocoetes villosus and Mycetochara maura, were among the least mobile species according to the classification by Irmler et al. (2010). The occurrence of the later species was significantly and strongly associated with the density of large or hollow oak density at small spatial scale (i.e. 52 m) (Bergman et al. 2012). Three others species (Prionus coriarius, Trypodendron signatum and Microrhagus pygmaeus) “are known to have occurred in recent times only in areas believed to be ancient woodland” (Alexander 2004). But we also found species that were more abundant and/or frequent in recent forests. This pattern may be caused by a trade-off between dispersal and competitive ability (Leibold et al. 2004). Species having high colonization capacity are poor competitors and are then more abundant or frequent in recent forests compare with ancient ones. Tritoma bipustulata appeared to be significantly more abundant but not more frequent in recent forests (Table 3). According to Alexander (2004) this species is restricted to ancient forests. Our contradictory finding may be explained by the differences observed between recent and ancient forests in terms of habitat quality (Table 1). Notably, deadwood volume was much higher in recent forests. This in turn may have consequences on the availability of fungi such as Polyporus spp., Trametes spp. or Daedalea spp. which T. bipustulata feeds on.
The effect of forest continuity was significant at the community level only in the Gascogne region. This may be due to a higher proportion of species influenced by continuity in this region, as reflected by results from the individual species models. Furthermore, spatial landscape features may interact with temporal discontinuity to contribute to extinction/colonization and diversity patterns (Brunet et al. 2011). The proportion of the landscape covered by hedgerows is lower in the Gatinais region (mean 3.22 ± 0.30 SE) than in the Gascogne region (mean 7.22 ± 0.42 SE). If such landscape elements do indeed contribute to woodlot connectivity, as they do for forest plants (Verheyen et al. 2003; Wehling and Diekmann 2009), the woodlots in the Gatinais region would have experienced a higher degree of isolation. This could have been detrimental to many poorly mobile species. In the Gascogne region, the higher forest landscape connectivity through hedgerow networks may compensate for the negative effects of temporal discontinuity on saproxylic beetle assemblages. Clustering patterns for all trait values, regardless of forest continuity, suggest habitat-filtering effects on the assembly process. The lower diversity of body sizes in recent forests can also be a sign of biotic homogenization due to disturbance (i.e. deforestation) (Devictor et al. 2008; Mori et al. 2015; Smart et al. 2006).
In conclusion, this study gives support to using forest continuity as a relevant criterion to identify sites of conservation importance, even in highly fragmented landscapes. Overall, our results may even be rather conservative, for two reasons. First, our recent forests had a higher volume of deadwood on average than our ancient forests. This may have reduced the difference between the two habitats. Second, some old-growth attributes such as large or highly decayed logs were rare in our ancient-forest study sites (Table 1). As these substrates harbor the most specialized and least mobile species (Gossner et al. 2013), the effect of forest continuity way have been under-estimated.
References
Alexander, K (2004) Revision of the Index of Ecological Continuity as used for saproxylic beetles. English Nature Research Reports, vol 574. English Nature, Peterborough
Andrieu E, Ladet S, Heintz W, Deconchat M (2011) History and spatial complexity of deforestation and logging in small private forests. Landsc Urban Plan 103:109–117
Bates D, Maechler M, Bolker B, Walker S, Christensen RHB, Singmann H, Dai B, Grothendieck G (2014). lme4 package: Linear mixed-effects models using ‘Eigen’ and S4 (https://cran.r-project.org/web/packages/lme4/index.html)
Bergman K-O, Jansson N, Claesson K, Palmer MW, Milberg P (2012) How much and at what scale? Multiscale analyses as decision support for conservation of saproxylic oak beetles. For Ecol Manag 265:133–141
Bouget C, Brustel H, Nageleisen LM (2005) Nomenclature of wood-inhabiting groups in forest entomology: synthesis and semantic adjustments. CR Biol 328:936–948
Bouget C, Brustel H, Zagatti P, Noblecourt T (2010) The French information system on saproxylic beetle ecology (FRISBEE): an ecological and taxonomical database to help with the assessment of forest conservation status. http://frisbee.nogent.cemagref.fr/index.php/en/
Bouget C, Brin A, Tellez D, Archaux F (2014a) Intraspecific variations in dispersal ability of saproxylic beetles in fragmented forest patches. Oecologia 177(3):1–10
Bouget C, Larrieu L, Brin A (2014b) Key features for saproxylic beetle diversity derived from rapid habitat assessment in temperate forests. Ecol Ind 36:656–664
Brändle M, Brandl R (2006) Is the composition of phytophagous insects and parasitic fungi among trees predictable? Oikos 113:296–304
Brin A, Bouget C, Brustel H, Jactel H (2011) Diameter of downed woody debris does matter for saproxylic beetle assemblages in temperate oak and pine forests. J Insect Conserv 15:653–669
Brown JH, Stevens GC, Kaufman DM (1996) The geographic range: size, shape, boundaries, and internal structure. Annu Rev Ecol Syst 27:597–623
Brunet J, Valtinat K, Mayr ML, Felton A, Lindbladh M, Bruun HH (2011) Understory succession in post-agricultural oak forests: habitat fragmentation affects forest specialists and generalists differently. For Ecol Manag 262:1863–1871
Buse J (2011) “Ghosts of the past”: flightless saproxylic weevils (Coleoptera: Curculionidae) are relict species in ancient woodlands. J Insect Conserv 16(1):1–10
Cateau E, Larrieu L, Vallauri D, Savoie JM, Touroult J, Brustel H (2015) Ancienneté et maturité: deux qualités complémentaires d’un écosystème forestier. Ancientness and maturity: two complementary qualitites of forest ecosystems. CR Biol 338:58–73
Cotgreave P (1993) The relationship between body size and population abundance in animals. Trends Ecol Evol 8:244–248
de Jong Y, Verbeek M, Michelsen V, de Place Bjørn P, Los W, Steeman F, Bailly N, Basire C, Chylarecki P, Stloukal E, Hagedorn G, Wetzel FT, Glöckler F, Kroupa A, Korb G, Hoffmann A, Häuser C, Kohlbecker A, Müller A, Güntsch A, Stoev P, Penev J (2014) Fauna Europaea–all European animal species on the web Biodivers Data J 2:e4034. doi:10.3897/BDJ.2.e4034
De Vries PG (1973) A general theory on line intersect sampling with application to logging residue inventory. Wageningen University, Wageningen
De Warnaffe GDB, Deconchat M, Ladet S, Balent G (2006) Variability of cutting regimes in small private woodlots of south-western France. Annals For Sci 63:915–927
Devictor V, Julliard R, Clavel J, Jiguet F, Lee A, Couvet D (2008) Functional biotic homogenization of bird communities in disturbed landscapes. Glob Ecol Biogeogr 17:252–261
Dupouey JL, Bachacou J, Cosserat R, Aberdam S, Vallauri D, Chapart G, Corvisier de Villèle M-A (2007) Vers la réalisation d’une carte géoréférencée des forêts anciennes de France. CFC 191:85–98
Goldberg E, Kirby K, Hall J, Latham J (2007) The ancient woodland concept as a practical conservation tool in Great Britain. J Nat Conserv 15:109–119
Gossner MM, Chao A, Bailey RI, Prinzing A (2009) Native fauna on exotic trees: phylogenetic conservatism and geographic contingency in two lineages of phytophages on two lineages of trees. Am Nat 173:599–614
Gossner MM, Engel K, Jessel B (2008) Plant and arthropod communities in young oak stands: are they determined by site history? Biodivers and Conserv 17(13):3165–3180
Gossner MM, Lachat T, Brunet J, Isacsson G, Bouget C, Brustel H, Brandl R, Weisser WW, Müller J (2013) Current near-to-nature forest management effects on functional trait composition of saproxylic beetles in beech forests. Conserv Biol 27:605–614
Grove SJ (2002) The influence of forest management history on the integrity of the saproxylic beetle fauna in an Australian lowland tropical rainforest. Biol Conserv 104:149–171
Hanski I (1999) Habitat connectivity, habitat continuity, and metapopulations in dynamic landscapes. Oikos 87:209–219
Hanski I (2000) Extinction debt and species credit in boreal forests: modelling the consequences of different approaches to biodiversity conservation. Ann Zool Fenn 37:271–280
Henle K, Davies KF, Kleyer M, Margules C, Settele J (2004) Predictors of species sensitivity to fragmentation. Biodivers Conserv 13:207–251
Hermy M, Verheyen K (2007) Legacies of the past in the present-day forest biodiversity: a review of past land-use effects on forest plant species composition and diversity. Ecol Res 22:361–371
Horák J, Vodka Š, Pavlíček J, Boža P (2013) Unexpected visitors: flightless beetles in window traps. J Insect Conserv 17:441–449
Hothorn T, Bretz F, Westfall P, Heiberger RM, Schuetzenmeister A, Scheibe S (2014) Multcomp package: simultaneous Inference in General Parametric Models (https://cran.r-project.org/web/packages/multcomp/index.html)
Inclán D, Cerretti P, Marini L (2014) Interactive effects of area and connectivity on the diversity of tachinid parasitoids in highly fragmented landscapes. Landsc Ecol 29:879–889
Irmler U, Arp H, Nötzold R (2010) Species richness of saproxylic beetles in woodlands is affected by dispersion ability of species, age and stand size. J Insect Conserv 14:227–235
Jackson ST, Sax DF (2010) Response to ‘Biodiversity “surpluses” and “deficits” are not novel issues’: we agree. Trends Ecol Evol 25:621–622
Kembel S, Ackerly D, Blomberg S, Cornwell W, Cowan P, Helmus M, Morlon H, Webb C (2014) Picante package: R tools for integrating phylogenies and ecology (https://cran.r-project.org/web/packages/picante/index.html)
Kuussaari M, Bommarco R, Heikkinen RK, Helm A, Krauss J, Lindborg R, Ockinger E, Partel M, Pino J, Roda F, Stefanescu C, Teder T, Zobel M, Steffan-Dewenter I (2009) Extinction debt: a challenge for biodiversity conservation. Trends Ecol Evol 24:564–571
Lachat T, Butler R (2009) Identifying conservation and restoration priorities for saproxylic and old-growth forest species: a case study in Switzerland. Environ Manag 44:105–118
Laliberté E, Legendre P, Shipley B (2014) FD package: measuring functional diversity (FD) from multiple traits, and other tools for functional ecology (https://cran.r-project.org/web/packages/FD/index.html)
Larjavaara M, Muller-Landau HC (2010) Comparison of decay classification, knife test, and two penetrometers for estimating wood density of coarse woody debris. Can J For Res 40:2313–2321
Lassauce A, Lieutier F, Bouget C (2012) Woodfuel harvesting and biodiversity conservation in temperate forests: effects of logging residue characteristics on saproxylic beetle assemblages. Biol Conserv 147:204–212
Lawton JH (1994) Population Dynamic Principles. Philos Trans R Soc Lond B 344:61–68
Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Law R, Tilman D, Loreau M, Gonzalez A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613
Marshall PL, Davis G, LeMay VM (2000) Using line intersect sampling for coarse woody debris, In Ecology F.R.T. Report. Forest Service, British Columbia, Vancouver Forest Region, p 34
Milberg P, Bergman K-O, Johansson H, Jansson N (2014) Low host-tree preferences among saproxylic beetles: a comparison of four deciduous species. Insect Conserv Divers 7:508–522
Mori A, Ota A, Fujii S, Seino T, Kabeya D, Okamoto T, Ito M, Kaneko N, Hasegawa M (2015) Biotic homogenization and differentiation of soil faunal communities in the production forest landscape: taxonomic and functional perspectives. Oecologia 177:533–544
Müller J, Bußler H, Bense U, Brustel H, Flechtner G (2005) Urwald relict species-Saproxylic beetles indicating structural qualities and habitat tradition Urwaldrelikt-Arten-Xylobionte Käfer als Indikatoren für Strukturqualität und Habitattradition. Waldökologie online, pp 106–113
Nagelkerke KC (2010) Biodiversity “surpluses” and “deficits” are not novel issues. Trends Ecol Evol 25:620–621 author reply 621-622
Nagelkerke KC, Verboom J, van den Bosch F, van den Wolfshaar K (2002) Time lags in metapopulation responses to landscape change. In: Gutzwiller KJ (ed) Concepts and applications of landscape ecology in biological conservation. Springer Verlag, New York, pp 330–354
Nieto A, Alexander KNA (2010) European red list of saproxylic beetles. Publications Office of the European Union, Luxembourg
Nilsson SG, Baranowski R (1997) Habitat predictability and the occurence of wood beetles in old-growth beech forests. Ecography 20:491–498
Norden B, Appelqvist T (2001) Conceptual problems of ecological continuity and its bioindicators. Biodivers Conserv 10:779–791
Norden B, Dahlberg A, Brandrud TE, Fritz O, Ejrnaes R, Ovaskainen O (2014) Effects of ecological continuity on species richness and composition in forests and woodlands: a review. Ecoscience 21:34–45
Patru-Stupariu I, Angelstam P, Elbakidze M, Huzui A, Andersson K (2013) Using forest history and spatial patterns to identify potential high conservation value forests in Romania. Biodivers Conserv 22:2023–2039
Pausas JG, Verdú M (2010) The jungle of methods for evaluating phenotypic and phylogenetic structure of communities. Bioscience 60:614–625
Pimm SL (1991) The balance of nature ?. University of Chicago Press, Chicago
R Development Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0, http://www.R-project.org/
Ranius T (2006) Measuring the dispersal of saproxylic insects: a key characteristic for their conservation. Popul Ecol 48:177–188
Rösch V, Tscharntke T, Scherber C, Batáry P (2013) Landscape composition, connectivity and fragment size drive effects of grassland fragmentation on insect communities. J Appl Ecol 50:387–394
Sauvard D (2004) General biology of bark beetles. In: Lieuthier F, Day KR, Battisti A, Grégoire J-C, Evans HF (eds) Bark and wood boring insects in living trees in Europe, a synthesis. Kluwer Academic Publishers, Dordrecht, pp 63–88
Seibold S, Brandl R, Buse J, Hothorn T, Schmidl J, Thorn S, Müller J (2015) Association of extinction risk of saproxylic beetles with ecological degradation of forests in Europe. Conservation Biology 29(2):382–390
Siitonen J, Saaristo L (2000) Habitat requirements and conservation of Pytho kolwensis, a beetle species of old-growth boreal forest. Biol Conserv 94:211–220
Siitonen J, Martikainen P, Punttila P, Rauh J (2000) Coarse woody debris and stand characteristics in mature managed and old-growth boreal mesic forests in southern Finland. For Ecol Manag 128:211–225
Similä M, Kouki J, Martikainen P (2003) Saproxylic beetles in managed and seminatural Scots pine forests: quality of dead wood matters. For Ecol Manag 174:365–381
Sippola AL, Siitonen J, Kallio R (1998) Amount and quality of coarse woody debris in natural and managed coniferous forests near the timberline in Finnish Lapland. Scand J For Res 13:204–214
Smart SM, Thompson K, Marrs RH, Le Duc MG, Maskell LC, Firbank LG (2006) Biotic homogenization and changes in species diversity across human-modified ecosystems. Proc R Soc B 273:2659–2665
Southwood TRE (1977) Habitat, the templet for ecological strategies? The Journal of Animal Ecology 46:337–365
Stokland J, Siitonen J, Jonsson BG (2012) Biodiversity in dead wood. University Press, Cambridge
Sverdrup-Thygeson A, Gustafsson L, Kouki J (2014) Spatial and temporal scales relevant for conservation of dead-wood associated species: current status and perspectives. Biodivers Conserv 23:513–535
Travis JMJ, Dytham C (1999) Habitat persistence, habitat availability and the evolution of dispersal. Proc R Soc Lond B 266:723–728
Vandekerkhove K, De Keersmaeker L, Walleyn R, Köhler F, Crevecoeur L, Govaere L, Thomaes A, Verheyen K (2011) Reappearance of old-growth elements in lowland woodlands in northern Belgium: do the associated species follow? Silva Fenn 45:909–935
Vanderwel MC, Malcolm JR, Smith SM (2006) An integrated model for snag and downed woody debris decay class transitions. For Ecol Manage 234(1–3):48–59
Vellend M, Verheyen K, Flinn KM, Jacquemyn H, Kolb A, Van Calster H, Peterken G, Graae BJ, Bellemare J, Honnay O, Brunet J, Wulf M, Gerhardt F, Hermy M (2007) Homogenization of forest plant communities and weakening of species-environment relationships via agricultural land use. J Ecol 95:565–573
Verheyen K, Guntenspergen GR, Biesbrouck B, Hermy M (2003) An integrated analysis of the effects of past land use on forest herb colonization at the landscape scale. J Ecol 91:731–742
Wang Y, Naumann U, Wright S, Warton D (2015) mvabund package: statistical methods for analysing multivariate abundance data (https://cran.r-project.org/web/packages/mvabund/index.html)
Warton DI (2011) Regularized sandwich estimators for analysis of high-dimensional data using generalized estimating equations. Biometrics 67:116–123
Warton DI, Wright ST, Wang Y (2012) Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol Evol 3:89–101
Wehling S, Diekmann M (2009) Importance of hedgerows as habitat corridors for forest plants in agricultural landscapes. Biol Conserv 142:2522–2530
Acknowledgments
The authors are grateful to F. Archaux for helpful comments during the project, and to H. Brustel, O. Courtin, S. Malaty, C. Mailliu, C. Victoire, L. Burnel, B. Nusillard, C. Moliard and A. N’Diaye for field and laboratory work. We thank O. Courtin and H. Brustel for the identification of a large part of the specimens. We thank V. Moore who checked the English language. We are also grateful to two anonymous reviewers for their helpful comments. This research was granted by the French Ministry in charge of Ecology through the “Biodiversité, Gestion Forestière et Politiques Publiques” (BGF) program (Contract Number: 10-MBGDBGF- 2-CVS-103 Distrafor).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Jens Wolfgang Dauber.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Brin, A., Valladares, L., Ladet, S. et al. Effects of forest continuity on flying saproxylic beetle assemblages in small woodlots embedded in agricultural landscapes. Biodivers Conserv 25, 587–602 (2016). https://doi.org/10.1007/s10531-016-1076-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-016-1076-z