
Abstract
Parrots (Psittaciformes) are among the most threatened bird orders with 28 % (111 of 398) of extant species classified as threatened under IUCN criteria. We confirmed that parrots have a lower Red List Index (higher aggregate extinction risk) than other comparable bird groups, and modeled the factors associated with extinction risk. Our analyses included intrinsic biological, life history and ecological attributes, external anthropogenic threats, and socio-economic variables associated with the countries where the parrot species occur, while we controlled for phylogenetic dependence among species. We found that the likelihood of parrot species being classified as threatened was less for species with larger historical distribution size, but was greater for species with high forest dependency, large body size, long generation time, and greater proportion of the human population living in urban areas in the countries encompassing the parrots’ home ranges. The severity of extinction risk (from vulnerable to critically endangered) was positively related to the per capita gross domestic product (GDP) of the countries of occurrence, endemism to a single country, and lower for species used as pets. A disproportionate number of 16 extinct parrot species were endemic to islands and single countries, and were large bodied, habitat specialists. Agriculture, hunting, trapping, and logging are the most frequent threats to parrots worldwide, with variation in importance among regions. We use multiple methods to rank countries with disproportionately high numbers of threatened parrot species. Our results promote understanding of global and regional factors associated with endangerment in this highly threatened taxonomic group, and will enhance the prioritization of conservation actions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The current worldwide decline in biodiversity requires urgent action (Butchart et al. 2010; Pimm et al. 2014; Tittensor et al. 2014), but determining appropriate actions is often impeded by poor understanding of the diverse causes of decline and extinction across different taxa and regions of the world (Spray and McGlothlin 2003). The factors that determine a species’ extinction risk fall into two main categories: (1) intrinsic biological attributes of species, and (2) exposure to external anthropogenic threats (Fisher et al. 2003). Although exposure to threats is the ultimate cause of extinction, a species’ biology often determines how sensitive it is to different threat types. Particular biological attributes can allow populations to recover rapidly from depletion and can offer a degree of resilience from external threats (Cardillo et al. 2004). Biological attributes that have been shown to decrease the risk of extinction include large geographical distribution, high population density, lower trophic level, and various life history traits including high reproductive rate, small body size, short generation length, and a low degree of habitat specialization (Owens and Bennett 2000; Purvis et al. 2000).
The order Psittaciformes (parrots) contains 398 extant species divided into 3 families (Psittacidae 374, Cacatuidae 21, Strigopidae 3 species) of which 111 (28 %) are classified by BirdLife International (2014) as threatened on the IUCN Red List [i.e. in the categories of Critically Endangered, Endangered or Vulnerable (IUCN 2014)]. Sixty species are classified as Near Threatened and the rest (227 species) are considered as Least Concern. Psittaciformes is the fourth largest bird order after Passeriformes (5913 spp., 10 % threatened), Caprimulgiformes (593 spp., 9 % threatened), and Piciformes (484 spp., 7 % threatened), but it contains the second highest number of threatened bird species after Passeriformes (containing 611 threatened species). Data from BirdLife International underpinning assessments for the IUCN Red List show that 56 % of all parrot species are in decline, 35 % are stable and only nine per cent have increasing populations (IUCN 2014).
Parrots are currently native to 124 countries between the latitudes N 35° and S 56°, but are mainly distributed in tropical and subtropical habitats of the southern hemisphere (Fig S1). They have been popular as pets throughout human history, probably due to their colourful appearance, reputedly high intelligence, and remarkable ability to mimic various sounds including the human voice (Grahl 1990). This popularity has led many species to be captured from the wild for pets. Although captive breeding has proven possible for many species (Arndt 2007; Tella and Hiraldo 2014), Psittaciformes remains the most common avian order reported in the wildlife trade (Bush et al. 2014).
Parrots exhibit many of the traits known to be associated with extinction: for example, many are large-bodied and slow-breeding (e.g. macaws and cockatoos), and 70 % are ecologically specialized (e.g. forest species) (Forshaw 2011). The main anthropogenic threats to parrots are habitat loss, degradation and fragmentation driven by agriculture, logging and commercial and residential development, and hunting and trapping (Beissinger et al. 1992; BirdLife International 2014; Laurance et al. 2002; Snyder et al. 2000). The socio-economic status of a country can determine the severity of anthropogenic threats affecting the species living there. For example, human population density was found to be closely related to the proportion of threatened bird species per country (Kerr and Currie 1995). Human activities such as poaching, hunting, and land clearing can be poverty driven in certain developing areas (Blaikie and Jeanrenaud 1997), but the effects of socio-economic drivers on the extinction risk of parrots have never been examined.
Although parrots have long been considered a group of especially high conservation concern (Pasquier 1980), in-depth quantitative analysis of the nature and trends in threats across the order has been lacking. Reviews to date have assessed particular threats (Beissinger and Bucher 1992; Harris et al. 2012; Laurance et al. 2009; Müller 2000; Newton 1994; Pain et al. 2006; Pires 2012), focused on individual species (Baker-Gabb 2011; Holdsworth and Starks 2006; Snyder et al. 2000; Webster et al. 2003) or assessed general trends in parrot conservation without comparative analysis (Collar 1997, 2000; Collar and Juniper 1992; Martin et al. 2014). Jones et al. (2006) advocated decision tree analysis for assessing extinction risk and used the IUCN Red List data of parrots from 2000 as an example group, but did not cover extrinsic extinction threats or socio-economic traits, and nor did they control for phylogenetic dependence of data. Our aim is to use comparative analyses to enhance understanding of the extrinsic, intrinsic, global and regional factors associated with endangerment and extinction in this highly threatened taxonomic group.
Methods
Red List Index
We examined trends in extinction risk during 1988–2012 for parrots (Psittaciformes) and other high profile ecological groups with similar numbers of species using Red List Indices (Butchart et al. 2007). These groups included waterbirds, seabirds, and raptors, each of which comprises multiple orders (see online supplementary methods). Other groups included pigeons (Columbiformes) and gamebirds (Galliformes). IUCN Red List criteria (e.g. absolute population size, range size, rates of decline, etc.) are used to assign species to Red List categories of relative extinction risk, and the Red List Indices are calculated from changes between these categories. Red List Indices for sets of species are based on the number of species in each category, and the number moving between categories owing to genuine improvement or deterioration in status, i.e. increases or decreases in population size, population trends, extent of occurrence, etc. that are of sufficient magnitude to cross the thresholds for lower or higher Red List categories (Butchart et al. 2004, 2005, 2007).
Database compilation
We assembled a database of the biological and geographic attributes of all 398 extant parrot species using the 2014 version of BirdLife International and IUCN’s database which underpins the IUCN Red List assessments for birds on the BirdLife Data Zone (BirdLife International 2014) and IUCN Red List website (IUCN 2014). Table 1 shows all explanatory variables (including sources) used in our statistical models of threat status. The online supplementary methods provide further details of the derivation of values in the database.
We used historical distribution size instead of current distribution sizes to test if species with originally small distributions are more prone to be endangered, and in order to avoid possible circularity as current extent of occurrence is a parameter used in the IUCN Red List criteria. Historical distribution is mapped using the same sources as for contemporary distribution, adding areas with historical records prior to 1980 for which the species is now judged to be extirpated owing to lack of recent records despite searches, lack of suitable habitat and informed by expert judgment (BirdLife International and NatureServe 2014). Population size and trend were not used as variables in our models for similar reasons. All the detailed underpinning data for Red List evaluation are available on the BirdLife Datazone (BirdLife International 2014). Because generation time was significantly positively correlated with body size (Table S1A), we calculated and used the residual values from the simple linear regression of generation time versus body size and referred to this variable as ‘residual generation time’ following Owens and Bennett (2000).
To assess socioeconomic and demographic attributes of the countries in the parrot species’ range, we used the World Economic Outlook Database (IMF 2013), the World Factbook (CIA 2013), and the database of the Food and Agriculture Organization of the United Nations (FAOSTAT 2013). We calculated the mean values of each parameter for all countries in which a species occurred (excluding vagrant records). Industrial production growth rate, unemployment rate, human population density, urban population, human population growth rate, and agriculture area were all significantly correlated with per capita gross domestic product (GDP) (Table S1B). We used residual values for these variables after regressing them against GDP.
We analyzed the traits of 16 extinct species separately comparing trends with values for extant species.
Statistical modeling procedures
We used linear logistic regression models to identify the broad covariates of whether or not a parrot species is classified as threatened. ‘Threatened’ was defined as belonging to the IUCN threat categories ‘Vulnerable’, Endangered’, and ‘Critically Endangered’, whereas ‘Non-threatened’ included the categories ‘Least Concern’ and ‘Near Threatened’. We then used ordinal regression models to evaluate in more detail the covariates of the degree of threat faced by the threatened parrot species. Both logistic and ordinal regression models were initially computed separately using each of four categories of variables (Table 1), and a final universal model was computed by combining the variables found to be significant in each of the sub-models. We used GenStat 13.7 (Payne 2009) for all modeling.
To control for phylogenetic relatedness among species we downloaded 1000 possible phylogenetic trees of Psittaciformes from birdtree.org. These were randomly selected to account for the branch lengths in addition to nodes separating species (Jetz et al. 2012). For each phylogeny, we ran a phylogenetic generalized least squares (PGLS) regression using the ‘caper’ package of R statistics (Freckleton et al. 2002). The explanatory and response variables were those from the best models per category found in the regression models. For each predictor in each model we report the modified p value accounting for phylogenetic relatedness, and we report λ (lambda transformation that improves the fit of the model to the phylogeny data) for each model. Greater values of λ indicate that the relationship between response and predictor variables is dependent on the phylogeny and trait values are more similar for closely related species. Values of λ closer to 0 indicate that the relationship is unrelated to phylogeny. Since these analyses were repeated for 1000 phylogenetic hypotheses, we report the mean and standard deviation of λ in each case. Additionally, we calculated in R the Blomberg´s K for threat status, which is a measure of phylogenetic signal. When values are closer to zero it indicates that the distribution of the trait is not related to phylogenetic relatedness.
We analyzed the specific threats associated with high extinction risk in parrots by assessing how many parrot species are affected by each threat type globally and specifically at the regional scale in the Neotropics, Afrotropics, Indomalaya, and Australasia/Oceania. We used threat impact scores estimated from timing, scope and severity for each threat for each species, and we compared the high and medium impact threats worldwide. We also considered the most important conservation actions needed to improve the status of threatened parrots. A formal prioritization analysis has not been carried out for all species, but the most urgent actions are documented by BirdLife International and therefore useful to present a broad level analysis.
We draw attention to countries with high risk of parrot conservation using two different methods. (1) We created a composite species ranking based on the sum of the ranks of the number of parrot species, number of globally threatened parrot species, and the number of endemic parrot species. We refer to these as ‘country priority’ scores. (2) We calculated the residual values of the final combined linear logistic regression model accounting for all significant extrinsic and intrinsic variables. We refer to these values as ‘unexplained extinction risk’ hereafter.
Results
Red list index for parrots and geographical differences
The Red List Index for parrots (0.825 in 2012) is lower than that for comparable groups of bird species including waterbirds (0.879), raptors (0.877), pigeons (0.868), gamebirds (0.842), and seabirds (0.828; Fig. 1). However, rates of decline from 1988 to 2012 are broadly comparable across these different groups: 1.27 % for parrots, 1.42 % for waterbirds, 1.75 % for raptors, 0.52 % for pigeons, 1.45 % for gamebirds, and 1.41 % for seabirds (Fig. 1).

Red List Indices of species survival for parrots (N = 398 non data deficient extant species; Psittaciformes), seabirds (N = 355; Anseriformes, Podicipediformes, Phaethontiformes, Gaviiformes, Sphenisciformes, Procellariiformes, Pelecaniformes, Suliformes, Charadriiformes), gamebirds (N = 307; Galliformes), pigeons (N = 350; Columbiformes), raptors (N = 320; Accipitriformes, Cathartiformes, Falconiformes), and waterbirds (N = 852; Anseriformes, Podicipediformes, Phoenicopteriformes, Gruiformes, Gaviiformes, Ciconiiformes, Pelecaniformes, Suliformes, Charadriiformes)
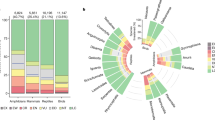
A total of 16 parrot species are considered to be Extinct, 17 are Critically Endangered (one species is Possibly Extinct in the Wild, and another is Possibly Extinct), 40 Endangered, 54 Vulnerable, 60 Near Threatened, and 227 are categorized as Least Concern. The list of threatened parrots species with their country of occurrence and the extinct parrots are shown in the supplementary Excel data file. Species density maps of all extant parrot species (Fig S1a) and of the threatened species (Fig S1b) are shown in the online supplementary material. The Neotropics have the highest proportion of threatened species (37 % of the region’s parrot species, N = 176), followed by Australasia/Oceania (22 %, N = 168 spp.), Afrotropics (19 %, N = 26 spp.), and Indomalaya (14 %, N = 28 spp.; Figure 2). Overall, there was no significant phylogenetic signal in degree of threat (K = 0.05, P = 0.36), which means that closely related species are not more likely to share the same Red List category (Fig. 3).

Proportion of parrot species in each IUCN Red List category for each region (following Croxall et al. 2012). Number of species in each category is shown

Phylogeny of parrots indicating IUCN Red List status of each species. Colors at the tip of the branches represent the IUCN Red List category of each species; grey shading inside the circle represent major genera and groups of related genera as labeled outside the circle. Images are taken with permission from the Handbook of Birds of the World online (Taylor 1996) and represent examples of species that are critically endangered. Source of phylogeny: www.birdtree.org
Threatened versus non-threatened
The probability of a parrot species being threatened decreased significantly with larger historical distribution size (combined linear logistic regression model, χ 21 = 64.89, P < 0.001; Fig S2a). The probability of being threatened increased significantly with: body size (χ 21 = 13.18, P < 0.001; Fig S2b), residual generation time (χ 21 = 16.82, P < 0.001; Fig S2c), extent of forest dependency (χ 23 = 29.47, P < 0.001; Fig S2d), and the percentage of the human population in the countries of occurrence living in urban areas (χ 21 = 28.71, P < 0.001; Fig S2e). There were no significant interactions among the 27 tested variables (all P ≥ 0.184). Table S3 lists additional variables found to be significant when analyzed separately by group (A–D). All significant variables in the combined linear logistic regression model were also significant in the PGLS model controlling for phylogeny (Table S5a).
Degree of threat
The severity of threat among threatened parrot species increased significantly with higher per capita GDP (Table 1) of the countries of occurrence (combined ordinal regression model, χ 21 = 8.47, P = 0.004; Fig S3a) and when the species is endemic to a single country (χ 21 = 5.6, P = 0.018; Fig S3b). The severity of threat was significantly less for species that are used as pets (χ 21 = 4.75, P = 0.029; Fig S3c). There were no significant interactions among the 27 tested variables (all P ≥ 0.140). Table S4 lists additional variables found to be significant when analyzed separately by group (A–D). All significant variables in the combined ordinal regression model were also significant in the PGLS model controlling for phylogeny (Table S5b).
Traits of extinct species
Sixteen parrot species in 12 genera are recorded as extinct. Five of the extinct species lived in the Neotropics (four in the Caribbean), six on islands near Africa (Mascarene Islands, Mauritius, Seychelles), and five in Australasia/Oceania (4 in Oceania and 1 in Australia). Fourteen out of 16 extinct species (88 %) inhabited islands only. By comparison one quarter of all extant parrot species (96 out of 398 species, 24 %) inhabit only islands a significant over-representation of island species amongst those now extinct (χ 21 = 28.5, P < 0.001). All but one extinct species were endemic to a single country, but distribution size was highly variable (38–557,670 km2) with the Carolina Parakeet (Conuropsis carolinensis) forming an outlier with range size of 3,167,000 km2 (mean of 240,510 km2 ± 198,000 SE for all 16 species). The mean of the extinct species’ median latitude of occurrence was −6.96 degrees ± 5.00 SE compared with −6.6 degrees ± 0.75 SE for extant species, with no significant difference (ANOVA F1,413 = 0.01, P = 0.936). Extinct species were mainly large bodied with a mean length of 39 cm ± 3.5 SE (ranging between 25 and 70 cm) compared with a mean of 28.5 cm ± 0.7 SE (ranging between 8 and 100 cm) for extant parrots (ANOVA F1,410 = 7.62, P = 0.006). Their estimated mean generation time was 8 years ± 0.7 SE compared with 7.3 years ± 0.2 SE for extant species (ANOVA F1,412 = 0.92, P = 0.339). All but one of the extinct species lived in only one habitat type, mainly in forests. Out of the 16 extinct species 38 % (6 species) were recorded as used as pets compared to 93 % today (372/398 species). A percentage of 69 % (11/16 spp.) of the extinct species were used as food compared to 23 % today (90/398 spp.).
Threat types, conservation actions, and priority countries
Based on the threat impact scores, the greatest threat to parrots worldwide is from agriculture (impacting 35 % of extant species), followed by hunting & trapping, logging, climate change & severe weather, and invasive alien species (Fig. 4a). The top three threats (agriculture, hunting & trapping, logging) appear roughly similar across regions, however residential development features as a greater threat in the Neotropics and Indomalaya, and invasive alien species is a greater threat in the Afrotropics and Australasia/Oceania (Fig. 4b). Whether species are threatened depends significantly on five major types of threat and their interactions: invasive alien species, agriculture, hunting & trapping, energy production & mining, and residential & commercial development (Table S6a). Higher categories of threat (i.e. Endangered, Critically Endangered) were especially associated with invasive alien species, even after controlling for phylogeny (χ 21 = 14.41, P < 0.001; Fig S3d, Table S6b).

Threats: a percentage of parrot species impacted by the 10 major threat types. Impact is calculated from scores for timing, scope and severity (see http://www.birdlife.org/datazone/info/spcthreat); b percentage of species in each region impacted by each threat type. Only threats impacting >15 species were plotted
The most important conservation actions as determined by BirdLife International (2014) are presented in Fig. 5. The most common actions needed in the Neotropics are site protection and management, in the Afrotropics they are legislation and ex-situ conservation, and in both Indomalaya and Australasia/Oceania they are awareness & communications, broad-scale habitat protection, and site protection (Fig. 5).

Most important conservation actions needed by region. The figure shows for each region the percentage of species for which each conservation action is considered required by BirdLife International (2014). Only conservation actions identified as required for >15 species across all regions were plotted. Order is given by total species showing the globally most important conservation actions first
Our country priority method revealed the following 10 countries with highest priority ranks: Indonesia, Brazil, Australia, Colombia, Bolivia, Ecuador, Peru, Papua New Guinea, Venezuela, and Mexico (more countries in Table 2). Examination of the unexplained extinction risk (after all significant intrinsic and extrinsic variables were accounted for in our models—see above) showed that island countries (or territories) accounted for both the lowest and highest values (Fig. 6). In total 40 countries had more threatened species than predicted (i.e. positive residuals), 17 of these were islands, 21 were in Africa. The Philippines (Indomalaya) and New Zealand (Australasia) also showed positive residuals (Fig. 6). In the Americas, all countries had less threatened species than predicted (with Brazil being closest to zero; Fig. 6).

Unexplained (residual) extinction risk of parrots by country and region: values shown are each country’s mean residuals from the combined linear logistic regression model predicting likelihood of a parrot species being threatened (VU, EN, CR). High positive values indicate high unexplained extinction risk
Discussion
Our analysis of Red List Indices revealed that parrots are more threatened than other comparable taxonomic groups (Fig. 1) with consistently negative trends in extinction risk over the last 25 years. The results show that conservation successes have been outweighed by the number of species being up-listed to higher categories of threat (IUCN 2014). Importantly, our analysis showed no significant effect of phylogeny on threatened status of parrots (Fig. 3). Extinction risk is usually non-randomly distributed with respect to phylogeny (Fisher and Owens 2004), hence the most illuminating comparative models are those like ours that focus on relatively narrow taxonomic groups (Cardillo et al. 2008).
Biological attributes
We found several biological attributes associated with extinction risk in parrots. We found that species with larger historical distributions are less likely to be threatened (Fig S2a). This probably reflects the link between historical and present distribution size, and the fact that widely distributed species are often adapted to multiple habitats and less impacted by local threats (Ewers and Didham 2006; Purvis et al. 2000).
As for other taxonomic groups, large bodied parrots are more prone to extinction risk (Fig S2b) (Bennett and Owens 1997; Cardillo 2003; Cardillo and Bromham 2001; Purvis et al. 2000). Large body size correlates with many known extinction-promoting traits, including low population densities, slower life histories (Cardillo et al. 2005), and higher risk from hunters (Cowlishaw and Dunbar 2000). The significant effect of residual generation time also indicates that parrots with slower life history (longer generation time) are more likely to be threatened, independently of body size (Fig S2c). Bennett and Owens (1997) also found that increasing body size and residual generation time were associated with increased extinction risk among birds when analyzed at the family level. Marsden and Royle (2015) showed that larger bodied parrots are predictably uncommon in the wild.
Our study confirmed that forest dependent parrot species are more likely to be threatened (Fig S2d). Most parrots are forest dependent because of their nesting and feeding habits (Snyder et al. 2000; White et al. 2005). At least 70 % of parrots are secondary tree cavity nesters (nesting in preexisting tree cavities), hence primary forest destruction decreases the number of available nest sites and reproductive success (Newton 1994). Woodpeckers, as potentially important keystone cavity excavators in Neotropical forests (Cornelius et al. 2008), can also be threatened by anthropogenic factors that ultimately affect parrot reproduction as well. Forest destruction can also lead to the loss of key food resources, as many parrots eat mostly tree seeds and fruits and rely on large areas of suitable habitat to provide year-round sources of these temporally variable food sources (Arndt 2007; Brightsmith 2005; Forshaw 2011; Juniper and Parr 2003).
Anthropogenic and socio-economic factors
Our data suggest a subtle effect of the extent of economic development of the countries where the parrots live. Whereas the extent of urbanization (percentage of human populations living in urban areas) seems to relate broadly to whether a parrot species is threatened (Fig S2e), the impact of per capita GDP (i.e. developed economies) seems to have most influence by pushing parrot species into higher categories of extinction risk (Fig S3a). Urbanization can entail far-reaching transformations in itself but is also linked to broad scale environmental degradation (Cohen 2006; McKinney 2002). As populations and economies of primarily rural countries grow, there is often a migration of people to urban areas, and this in turn can be followed by further economic expansion and higher per capita GDP (Chang and Brada 2006; Moomaw and Shatter 1996). Hence urbanization can occur at high levels before GDP is maximized (Brülhart and Sbergami 2009; Henderson 2003). Our results suggest that urbanization is a good variable accounting for the impact on parrots of the earlier stages of economic development, with the continuing impact of economic development best captured by GDP.
Parrots with distributions limited to one country are also more likely to belong to higher categories of endangerment (Fig S3b). Our analysis thus suggests that extinction risk of single country endemics (45 % of all parrot species) is often adversely affected by either the singular history or the conservation management practices (or lack thereof) of their single country of occurrence. In the latter case, specific threats may be better controlled in some nations than others (e.g. progressive forestry laws and control of pet trade) with potential advantages to species living in more than one country (Hirakuri 2003; Pires 2012; Ribot 1999; Sunderlin et al. 2005).
Our finding that threatened species used for pets tend to belong to the lower categories of endangerment may seem counterintuitive (Fig S3c) but is supported by recent studies which show that the vast majority of species in domestic and international bird trade are non-threatened species (Pires and Clarke 2012; Pires 2012) and that utilized species in general are less threatened (Butchart 2008). This finding likely speaks much more to how increased threatened status (or decreased abundance) reduces use as pets than it does to how use for pets causes increases in threatened status. Pires and Clarke (2012) report that rare parrot species are only infrequently found in illegal parrot markets, confirming that most parrot poaching is driven by species that are easier to catch rather than by commercially based poachers searching for the rarest species.
Traits of extinct species
Our analysis of the traits of extinct species provides additional insight on some of our key results concerning extinction risk of extant species. For example, a disproportionate number of extinct parrot species (88 % vs. 24 % for extant species) were insular, yet our models for extant species did not show that island endemism was a significant factor. This suggests that island endemism has been a strong factor in extinction risk but may have been masked in our analyses of extant species because many of the most susceptible species have already become extinct. In keeping with our models, extinct parrot species were larger on average than extant parrots, and most extinct species (94 %) were single country endemics (compared to 45 % for extant species).
Threat types and conservation actions
Our analysis reveals that the most important threats to parrots are agriculture, hunting and trapping, logging, climate change and severe weather, invasive alien species, and residential and commercial development (Fig. 4a, Table S6a). Invasions by non-native species are often cited as leading causes of species extinctions (Carrete and Tella 2008; Gurevitch and Padilla 2004), but our analysis suggests more complex synergistic effects in combination with other threats, including hunting and trapping, and agriculture (see interactions in Table S6a). Interestingly, ‘invasive alien species’ was the sole threat significantly associated with the higher categories of threat status (Fig S3d, Table S6b). This warrants increasing conservation attention on invasive alien species. These threats do not seem to respond to the major conservation strategies that are normally applied (e.g. country border protection, legislation, law enforcement). Rather invasive species are a threat that needs to be combatted directly and individually through regionally designed conservation action plans.
We found regional differences in the threats impacting parrots and hence the identified conservation actions required. Agriculture is a particularly important threat worldwide in terms of number of parrot species impacted (Fig. 4a), and especially in the Neotropics (Fig. 4b) where site protection is identified as the major conservation action required (Fig. 5). In Indomalaya and Australasia/Oceania, the most significant threat to parrots is logging (Fig. 4b), driving extensive deforestation in recent decades (Sodhi et al. 2004, 2010a, b). Reflecting this, the most common conservation actions needed are broad-scale habitat protection, site protection and awareness raising (Fig. 5). In contrast, the most common threat in the Afrotropics is hunting and trapping (primarily for the cage-bird trade and for use as pets; Fig. 4b), and the most important key actions include enforcing and enhancing legislation (Fig. 5).
Priority countries
Countries with the highest conservation priority were from all regions except the Afrotropics. Although Indonesia ranked the highest overall and Australia had the highest value amongst developed countries, 15 out of the top 20 parrot conservation priority countries were from the Neotropics (Table 2). These countries deserve high conservation interest given the high diversity and endemism of their parrot species, and the high proportion of their parrot species that are threatened. After accounting for all the significant variables in our model (historical distribution size, body size, generation time, forest dependency, and percentage of urban human population in the distribution ranges), the mean residual (or unexplained) extinction risk values of each country suggested some variation in country-level performances. The countries with positive residuals (on average more threatened species than expected) included many islands of different sizes (island nations, Philippines, and New Zealand; Fig. 6). Interestingly, island countries and territories also had the lowest mean residuals (less threatened species than expected; e.g. Bahamas, Comoros, Micronesia) in conservation status (Fig. 6). Considering only mainland countries, Africa showed the worst trends compared to the other regions, where capture for the pet trade is still common (Fig. 6). These results again confirm the impact on parrots of living on islands, and our method may prove important for identifying poorly performing countries (e.g. Dominican Republic, Haiti, Brazil, Burundi, Philippines, New Zealand, etc.) that need both extra conservation attention and further research on the nature of threatening processes.
Conclusion
Our study confirms that parrots are more threatened on average than comparable bird species groups, and that biological factors known to affect extinction risk in other taxa also apply to parrots. Thus parrot species with small historical distribution size, large body size, long generation time, and forest dependency are most likely to be categorized as threatened (Vulnerable, Endangered, Critically Endangered) under IUCN Red List criteria. Extinct parrot species shared most of these traits but also highlight island endemism as an important factor in the past. The extent of a country’s urbanization provides a broad proxy for the major human socio-economic drivers of extinction risk in parrots. Our models also revealed that parrots are more likely to be highly threatened if their distribution falls within a single country’s jurisdiction and in countries with higher per capita GDP, presumably because the higher levels of development that these factors are associated with, tend to drive the major threats to parrots worldwide including agriculture, hunting and trapping, and logging.
References
Arndt T (2007) Lexicon of Parrots. Arndt Verlag, Bretten
Baker-Gabb D (2011) National Recovery Plan for the Superb Parrot Polytelis swainsonii. Department of Sustainability and Environment, Melbourne
Beissinger SR, Bucher EH (1992) Can parrots be conserved through sustainable harvesting? Bioscience 42:164–173
Beissinger SR, Snyder NFR, Thomsen JB, Mulliken TA (1992) Trade in neotropical psittacines and its conservation implications. New world parrots in crisis: solutions from conservation biology. Smithsonian Institution Press, New York and London, pp 221–239
Bennett PM, Owens IPF (1997) Variation in extinction risk among birds: chance or evolutionary predisposition? Proc R Soc Lond B Biol Sci 264:401–408. doi:10.1098/rspb.1997.0057
BirdLife International (2014) The BirdLife checklist of the birds of the world: Version 7. http://www.birdlife.org/datazone/userfiles/file/Species/Taxonomy/BirdLife_Checklist_Version_70.zip
BirdLife International and NatureServe (2014) Bird species distribution maps of the world. Version 4.0. BirdLife International, Cambridge, UK and NatureServe, Arlington, USA. http://www.birdlife.org/datazone/info/spcdownload
Blaikie P, Jeanrenaud S (1997) Biodiversity and human welfare. Social change and conservation. Earthscan, London, pp 46–70
Brightsmith DJ (2005) Competition, predation and nest niche shifts among tropical cavity nesters: ecological evidence. J Avian Biol 36:74–83
Brülhart M, Sbergami F (2009) Agglomeration and growth: cross-country evidence. J Urban Econ 65:48–63. doi:10.1016/j.jue.2008.08.003
Bush ER, Baker SE, Macdonald DW (2014) Global trade in exotic pets 2006–2012. Conserv Biol 28:663–676. doi:10.1111/cobi.12240
Butchart SHM (2008) Red List Indices to measure the sustainability of species use and impacts of invasive alien species. Bird Conserv Int 18:S245–S262. doi:10.1017/S095927090800035X
Butchart SHM et al (2004) Measuring global trends in the status of biodiversity: Red List Indices for birds. PLoS Biol 2:383
Butchart SHM et al (2005) Using Red List Indices to measure progress towards the 2010 target and beyond. Philos Trans R Soc B 360:255–268. doi:10.1098/rstb.2004.1583
Butchart SHM et al (2007) Improvements to the Red List Index. PLoS One 2:e140. doi:10.1371/journal.pone.0000140
Butchart SHM et al (2010) Global biodiversity: indicators of recent declines. Science 328:1164–1168. doi:10.1126/science.1187512
Cardillo M (2003) Biological determinants of extinction risk: why are smaller species less vulnerable? Anim Conserv 6:63–69
Cardillo M, Bromham L (2001) Body size and risk of extinction in Australian mammals. Conserv Biol 15:1435–1440
Cardillo M, Purvis A, Sechrest W, Gittleman JL, Bielby J, Mace GM (2004) Human population density and extinction risk in the world’s carnivores. PLoS Biol 2:e197. doi:10.1371/journal.pbio.0020197
Cardillo M et al (2005) Multiple causes of high extinction risk in large mammal species. Science 309:1239–1241. doi:10.1126/science.1116030
Cardillo M, Mace GM, Gittleman JL, Jones KE, Bielby J, Purvis A (2008) The predictability of extinction: biological and external correlates of decline in mammals. Proc R Soc B 275:1441–1448. doi:10.1098/rspb.2008.0179
Carrete M, Tella JL (2008) Wild-bird trade and exotic invasions: a new link of conservation concern? Front Ecol Environ 6:207–211
Chang GH, Brada JC (2006) The paradox of China’s growing under-urbanization. Econ Syst 30:24–40. doi:10.1016/j.ecosys.2005.07.002
CIA (2013) Central Intelligence Agency: The World Factbook. https://www.cia.gov/library/publications/the-world-factbook/. Accessed on June 2013
Cohen B (2006) Urbanization in developing countries: Current trends, future projections, and key challenges for sustainability. Technol Soc 28:63–80. doi:10.1016/j.techsoc.2005.10.005
Collar NJ (1997) Family Psittacidae (parrots). In: Hoyo Jd, Elliott A, Sargatal J (eds) Handbook of the birds of the world, vol 4. Lynx Edicions, Barcelona, pp 280-477
Collar NJ (2000) Globally threatened parrots: criteria, characteristics and cures. Int Zoo Yearb 37:21–35
Collar NJ, Juniper AT (1992) Dimensions and causes of the parrot conservation crisis. In: Beissinger SR, Snyder NFR (eds) New World parrots in crisis: solutions from conservation biology. Smithsonian Institution Press, Washington, DC, pp 1–24
Cornelius C et al (2008) Cavity-nesting birds in Neotropical forests: cavities as a potentially limiting resource. Ornitol Neotrop 19:253–268
Cowlishaw G, Dunbar RIM (2000) Primate conservation biology. University of Chicago Press, Chicago
Croxall JP, Butchart SHM, Lascelles B, Stattersfield AJ, Sullivan B, Symes A, Taylor P (2012) Seabird conservation status, threats and priority actions: a global assessment. Bird Conserv Int 22:1–34. doi:10.1017/S0959270912000020
Ewers RM, Didham RK (2006) Confounding factors in the detection of species responses to habitat fragmentation. Biol Rev 81:117–142. doi:10.1017/S1464793105006949
FAOSTAT (2013) Food and Agriculture Organization of the United Nations. http://faostat3.fao.org/. Accessed on July 2013
Fisher DO, Owens IPF (2004) The comparative method in conservation biology. Trends Ecol Evol 19:391–398. doi:10.1016/j.tree.2004.05.004
Fisher DO, Blomberg SP, Owens IPF (2003) Extrinsic versus intrinsic factors in the decline and extinction of Australian marsupials. Proc R Soc Lond B 270:1801–1808. doi:10.1098/rspb.2003.2447
Forshaw JM (2011) Parrots of the world CSIRO Publishing. Collingwood, Australia
Freckleton RP, Harvey PH, Pagel M (2002) Phylogenetic analysis and comparative data: a test and review of evidence. Am Nat 160:712–726. doi:10.1086/343873
Grahl Wd (1990) Papageien: Lebensweise, Arten, Zucht, 9th edn. Verlag Eugen Ulmer, Germany
Gurevitch J, Padilla DK (2004) Are invasive species a major cause of extinctions? Trends Ecol Evol 19:470–474. doi:10.1016/j.tree.2004.07.005
Harris JB et al (2012) Managing the long-term persistence of a rare cockatoo under climate change. J Appl Ecol 49:785–794
Henderson V (2003) The urbanization process and economic growth: the so-what question. J Econ Growth 8:47–71. doi:10.1023/A:1022860800744
Hirakuri SR (2003) Can law save the forest?: lessons from Finland and Brazil. CIFOR, Bogor
Holdsworth M, Starks J (2006) National Recovery Plan for the Orange-bellied Parrot (Neophema chrysogaster). Department of Primary Industries and Water (DPIW), Hobart, Australia
IMF (2013) International Monetary Fund: world economic outlook database. http://www.imf.org/. Accessed on June 2013
IUCN (2014) The IUCN Red list of threatened species. Version 2014.2. http://www.iucnredlist.org
Jetz W, Thomas GH, Joy JB, Hartmann K, Mooers AO (2012) The global diversity of birds in space and time. Nature 491: 444–448. http://www.nature.com/nature/journal/v491/n7424/abs/nature11631.html - supplementary-information
Jones M, Fielding A, Sullivan M (2006) Analysing extinction risk in parrots using decision trees. Biodivers Conserv 15:1993–2007. doi:10.1007/s10531-005-4316-1
Juniper T, Parr M (2003) Parrots: a guide to parrots of the world. A & C Black, USA
Kerr JT, Currie DJ (1995) Effects of human activity on global extinction risk. Conserv Biol 9:1528–1538. doi:10.1046/j.1523-1739.1995.09061528.x
Laurance WF, Albernaz AKM, Schroth G, Fearnside PM, Bergen S, Venticinque EM, Da Costa C (2002) Predictors of deforestation in the Brazilian Amazon. J Biogeogr 29:737–748. doi:10.1046/j.1365-2699.2002.00721.x
Laurance WF, Goosem M, Laurance SGW (2009) Impacts of roads and linear clearings on tropical forests. Trends Ecol Evol 24:659–669. doi:10.1016/j.tree.2009.06.009
Marsden SJ, Royle K (2015) Abundance and abundance change in the world’s parrots. Ibis 157:219–229. doi:10.1111/ibi.12236
Martin RO et al (2014) Research and conservation of the larger parrots of Africa and Madagascar: a review of knowledge gaps and opportunities. Ostrich 85:205–233. doi:10.2989/00306525.2014.948943
McKinney ML (2002) Urbanization, biodiversity, and conservation: the impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 52:883–890. doi:10.1641/0006-3568(2002)052[0883:ubac]2.0.co;2
Moomaw RL, Shatter AM (1996) Urbanization and economic development: a Bias toward Large Cities? J Urban Econ 40:13–37. doi:10.1006/juec.1996.0021
Müller M (2000) Review of the in situ status of the Great green or Buffon’s macaw Ara ambigua and the European Endangered Species Programme (EEP). Int Zoo Yearb 37:183–190
Newton I (1994) The role of nest sites in limiting the numbers of hole-nesting birds: a review. Biol Conserv 70:265–276. doi:10.1016/0006-3207(94)90172-4
Olson DM et al (2001) Terrestrial ecoregions of the world: a new map of life on earth. Bioscience 51:933–938. doi:10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Owens IPF, Bennett PM (2000) Ecological basis of extinction risk in birds: habitat loss versus human persecution and introduced predators. Proc Natl Acad Sci 97:12144–12148. doi:10.1073/pnas.200223397
Pain DJ et al (2006) Impact of protection on nest take and nesting success of parrots in Africa, Asia and Australasia. Anim Conserv 9:322–330. doi:10.1111/j.1469-1795.2006.00040.x
Pasquier RF (1980) Conservation of new world parrots: proceedings of the ICBP Parrot Working Group Meeting, St. Lucia, 1980. Smithsonian Institution Press for the International Council for Bird Preservation, USA
Payne RW (2009) GenStat. Wiley Interdisciplinary Reviews: Computational Statistics 1:255–258. doi:10.1002/wics.32
Pimm SL et al (2014) The biodiversity of species and their rates of extinction, distribution, and protection. Science. doi:10.1126/science.1246752
Pires SF (2012) The illegal parrot trade: a literature review. Glob Crime 13:176–190
Pires S, Clarke RV (2012) Are parrots CRAVED? An analysis of parrot poaching in Mexico. J Res Crime Delinq 49:122–146
Purvis A, Gittleman JL, Cowlishaw G, Mace GM (2000) Predicting extinction risk in declining species. Proc R Soc Lond B Biol Sci 267:1947–1952. doi:10.1098/rspb.2000.1234
Ribot JC (1999) Decentralisation, participation and accountability in Sahelian forestry: legal instruments of political-administrative control. Africa 69:23–65. doi:10.2307/1161076
Snyder N, McGowan P, Gilardi J, Grajal A (2000) Parrots. Status Survey and Conservation Action Plan 2000–2004. IUCN, Gland, Switzerland and Cambridge, UK
Sodhi NS, Koh LP, Brook BW, Ng PKL (2004) Southeast Asian biodiversity: an impending disaster. Trends Ecol Evol 19:654–660. doi:10.1016/j.tree.2004.09.006
Sodhi N, Posa M, Lee T, Bickford D, Koh L, Brook B (2010a) The state and conservation of Southeast Asian biodiversity. Biodivers Conserv 19:317–328. doi:10.1007/s10531-009-9607-5
Sodhi NS et al (2010b) Deforestation and avian extinction on tropical landbridge islands. Conserv Biol 24:1290–1298. doi:10.1111/j.1523-1739.2010.01495.x
Spray SL, McGlothlin KL (2003) Loss of biodiversity. Exploring environmental challenges. Rowman & Littlefield, Lanham
Sunderlin WD, Angelsen A, Belcher B, Burgers P, Nasi R, Santoso L, Wunder S (2005) Livelihoods, forests, and conservation in developing countries: an overview. World Dev 33:1383–1402. doi:10.1016/j.worlddev.2004.10.004
Taylor B (1996) Sulu Racquet-tail (Prioniturus verticalis), White Cockatoo (Cacatua alba), New Zealand Kaka (Nestor meridionalis), Blue-throated Macaw (Ara glaucogularis), Grey-breasted Parakeet (Pyrrhura griseipectus), Puerto Rican Amazon (Amazona vittata), Orange-bellied Parrot (Neophema chrysogaster), Malherbe’s Parakeet (Cyanoramphus malherbi), Red-throated Lorikeet (Charmosyna amabilis). In: del Hoyo J, Elliott A, Sargatal J, Christie DA, de Juana E (eds) (2013). Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona. http://www.hbw.com. Accessed on 12 Nov 2014
Tella JL, Hiraldo F (2014) Illegal and legal parrot trade shows a long-term, cross-cultural preference for the most attractive species increasing their risk of extinction. PLoS One 9:e107546. doi:10.1371/journal.pone.0107546
Tittensor DP et al (2014) A mid-term analysis of progress toward international biodiversity targets. Science 346:241–244. doi:10.1126/science.1257484
Webster A, Price R, Lowe K (2003) Action statement: Swift Parrot (Lathamus discolor). Department of Sustainability and Environment, Victoria
White TH Jr, Collazo JA, Vilella FJ (2005) Survival of captive-reared Puerto Rican parrots released in the Caribbean National Forest. The Condor 107:424–432
Acknowledgments
The authors thank the many thousands of individuals and organizations who contribute to BirdLife International’s assessments of parrots for the IUCN Red List, upon which the analyses presented here are based. We thank Hannah Wheatley for the management of these data, Mark Balam and Janet Nackoney for help with GIS mapping, Rod Peakall for commenting on the manuscript, Kurt Holle and Jenö Gászpor for helping with the socio-economic variables, and Marta Romhányi, Attila Romhányi, and Braulio Poje for helping with the database construction.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Stuart Pimm.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Olah, G., Butchart, S.H.M., Symes, A. et al. Ecological and socio-economic factors affecting extinction risk in parrots. Biodivers Conserv 25, 205–223 (2016). https://doi.org/10.1007/s10531-015-1036-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-015-1036-z