
Abstract
African wild dogs (Lycaon pictus) are often the least popular large carnivore among game ranchers because of their perceived impact on prey populations. Landowner perceptions include that wild dogs greatly deplete prey during their three-month denning period, take prey that could otherwise be sold for hunting and cause prey to move away from the vicinity of their den sites. Landowners’ tolerance towards African wild dogs could thus be improved with a more rigorous understanding of the actual impact of wild dogs on prey populations during the denning period. Using impala density data and wild dog denning records from Sango Ranch in the Savé Valley Conservancy, Zimbabwe, we compared impala densities between pre-denning, denning and post-denning season and between inside and outside the denning home range. Our results indicate that wild dog denning does not cause a significant local reduction in prey around the den and does not cause prey to move away from denning areas. However prey species did occur in lower density inside the denning home ranges than outside, in all seasons. This result indicates that wild dogs select dens in areas of lower prey density, perhaps as an avoidance mechanism for lions. Accordingly, contrary to what landowners believe, wild dogs do not have a significant impact on prey populations during their denning season.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Carnivore predation behaviour has been found to have negative effects on both prey populations (directly through predators) and on predators (indirectly through human persecution) and this often has profound implications in human-wildlife conflicts (Moleo′n et al. 2011). Indeed one of the challenges facing the conservation of large carnivores today is the conflict between their conservation needs and human interests (Mills 1991; Sillero-Zubiri and Laurenson 2001). The greatest source of mortality for many large carnivore species around the world is persecution by humans (Woodroffe and Ginsberg 1998). Unfortunately wild canids have been most affected by human negative attitudes (Kellert 1985), especially the endangered African wild dog (Lycaon pictus) (hereafter wild dog). The wild dog has performed badly in the public interests compared to other species such as the lion (Panthera leo) (Fanshawe et al. 1991) and leopard (Panthera pardus) (Lindsey et al. 2005). In some parts of Southern Africa, game ranching ventures on large tracts of land have replaced cattle ranching and this is key to the long term survival of wild dogs (Lindsey et al. 2005). The establishment of large areas under wildlife enabled the re-colonisation of wild dogs into parts of their former range in Zimbabwe (Pole 1999; Rasmussen 1999) and in South Africa (Lindsey et al. 2004). However negative perceptions surrounding wild dogs remain prevalent which potentially risk the chances for its survival within these areas.
Predation of trophy species particularly in trophy hunting areas can lead to serious conservation problems when it comes to threatened and endangered predators such as the wild dog, as it often leads to serious persecution of the carnivore (Moleo′n et al. 2011). This type of conflict is particularly prevalent in southern Africa, where there is an active trophy hunting industry (Lindsey et al. 2005). Negative attitudes by wildlife ranchers towards wild dogs stem from the perceptions that potential income is impaired through the killing of prey species and that the mere presence of wild dogs, particularly during the denning seasons, causes animals to flee the area or hide, thus decreasing the likelihood of successful hunts (Fanshawe et al. 1991). Other largely inaccurate perceptions held about wild dogs include the suggestion that wild dogs regularly kill more than they need, and that they terrorise their prey (Woodroffe and Ginsberg 1999). Wild dogs have experienced the most persecution because they are not legally trophy hunted and so are perceived to impose costs on landowners without generating any financial returns (Lindsey et al. 2007). Although state-sponsored persecution, such as that which occurred in Zimbabwe between 1956 and 1975 (Childes 1985) has ceased, many of the myths and misperceptions surrounding wild dogs still persist (Lindsey et al. 2005), and this promotes continued persecution outside protected areas especially on private land (Rasmussen 1999).
For most of the year, wild dogs use large home range areas, averaging 423–1,318 km2 (range 150–2,460 km2) (Hunter 2011). However, during the denning period, wild dogs use a restricted range stretching from 50 to 260 km2 (Hunter 2011) and averaging 80 km2 (Gorman et al. 1992). During the denning period wild dog packs choose a den (often in burrows in termite mounts, originally excavated by other species), in which to give birth and raise a litter. The pups are confined to the den for about 3 months, and the adults return to the den after each hunt (Malcolm and Marten 1982). Denning is the only time in the year when the otherwise wandering wild dogs stay in one place (Fanshawe et al. 1991). Consequently the use of such a restricted range during denning increases the chances of wild dogs impacting on the behaviour and density of their prey (Romañach and Lindsey 2008).
The effect predators have on prey species are extremely difficult to measure accurately (Mills 1991) and there has been a lack of studies on the direct effects of predation as it is often difficult to obtain reliable data on prey density and distribution, and unbiased data on predation patterns. However, an extensive database on impala (Aepyceros melampus) density and distribution and denning records of wild dogs in the Savé Valley Conservancy (SVC) in south eastern Zimbabwe, enabled us to analyse the relationship between wild dog denning and prey density. Since impala are the main prey species (>74 % in diet) for wild dogs in the study area (Pole et al. 2004; Mbizah et al. 2012) the effect of wild dogs on impala density during the denning season were investigated and tested against the perception that wild dogs deplete prey in the areas surrounding den sites during the denning period. We also tested the belief that wild dog predation may force their main prey species (impala) to alter their behaviour by moving beyond the range of the den site. In addition, we tested the hypothesis that prey density around wild dog dens is low because they select to den in areas with low prey density.
Methods and materials
Study area
SVC is a privately-owned area of ~3,490 km2 in the semi-arid southeast region of Zimbabwe (20°05′S and 32°00′E) (Romañach and Lindsey 2008). The climate is characterized by a single wet season from November to March, and a long dry season from April to October with a mean annual rainfall that varies between 300 and 500 mm (du Toit 1994). The study area has abundant populations of wildlife including all indigenous large carnivores. Wild dogs re-colonized the SVC during the early 1990s, after having been extinct for about 20 years (Pole 1999). Trophy hunting is the main land use in the SVC due to the recent collapse in the ecotourism industry in Zimbabwe (Romañach and Lindsey 2008). This study was carried out in the central part of the SVC, on Sango Ranch, which is divided into 884 grids, each measuring 1.0 km2 in area (Fig 1).

Map showing the survey grids in Sango Ranch that were used for the analysis as well as the locations of wild dog dens from 2004 to 2010 and the insert map showing the location of Savé Valley Conservancy in Zimbabwe
Wild dog dens and home ranges
Wild dogs in the SVC have been intensively monitored since 1996 using spoor tracking by experienced trackers, collars and radio telemetry, photographs obtained directly or from camera traps and observations at den sites. These methods provided accurate information on population dynamics and behaviour. In the SVC, pups are usually born early in the dry season, between May and August of each year. For the purposes of this study, we used a total of 18 den sites (Fig. 1) recorded over a seven-year observation period, and including only the first den site chosen by each pack each year. Wild dogs will occasionally move to a new den during the denning season, but in this study, only three packs did a single den move. A total of ten different packs were recorded in the study area from 2004 to 2010 with an average pack size of eight adults and yearlings (range; 2–23 adults and yearlings).
Global Positioning System (GPS) collar data was only available from 2009 onwards and this was used to calculate the average denning and non-denning home range sizes for wild dog packs and then applied to all packs included in the study. Since 2009, 15 individuals from six different wild dog packs in the SVC were fitted with GPS collars programmed to take eight fixes per day. GPS fixes covered a daytime resting location (12:00 noon) a night time resting point (01:00), the morning hunting period (06:00, 07:00 & 08:00) and the evening hunting period (17:00, 18:00 & 19:00). We calculated the average denning period using the dates between which the six packs denned each year, this resulted in an average denning period of June–August. Location data from GPS fixes were then used to calculate home ranges for the denning period (June–August inclusive) and the non denning period (September–May inclusive), using the Home Range Tools and the Kernel Density Estimate (KDE) function extensions in Quantum GIS (1.7.3) (Quantum GIS Development Team 2012). Kernel isopleths were calculated using the reference smoothing factor href as recommended by Hemson et al. (2005) and a 90 % isopleth.
Impala data
Impala densities in Sango Ranch from 2004 to 2010 were determined using the grid square method (Joubert 2013), based on inputs from daily game scout patrol reports. Each game scout was allocated a map designed in ESRI ® ArcMap ™ 10 (License ArcView) which was subdivided into 1 km2 grid squares. At each animal sighting game scouts recorded the type of species, the number seen, the date and the grid reference. Relative density estimates and distribution for impala was thus calculated to a level of 1 km2. Time spent in each grid was not recorded, so the assumption is that game scouts covered each grid equally and that impala are equally detectable in all grids. We believe these assumptions did not bias our density estimates since impala are usually in big herds, making them easier to detect, and also because there was a generally comprehensive and even coverage of the study area by the game scouts. For the purpose of this study only impala data (42 024 impala observation records) were used in the analysis while the data for the other ungulates was excluded.
In this analysis we used den site location and impala densities for the period 2004–2010. We then used a generic three-month period, June–August (calculated earlier as being the average denning period) as the denning season for all wild dog packs, as this allowed us to increase sample size by including dens recorded before the use of GPS collars. This is justified because the wild dogs in the SVC tend to have a very synchronous breeding season, with relatively little variability in denning dates. The area calculated as the average denning season home range (90 km2; N = 6 packs) was used to demarcate a circular area of the same size around each den site. We calculated impala densities inside the denning home range and outside the denning home range by averaging the density in grids that fell within the denning home range and grids that fell outside the denning home range respectively. For each grid and each year, impala densities were averaged into three different seasons: pre-denning season (January–May), denning season (June–August) and post-denning season (September–December).
Impala densities in the pre-denning, denning and post-denning season
The hypothesis that wild dogs deplete prey within their denning season home range, and that they drive prey away from the denning range was tested using linear mixed models. To assess the effect of season (pre-denning, denning and post-denning) on impala densities the following linear mixed model was used:
where impala density is the response variable, season is the fixed-effect with year (2004–2010) as a random-effect. The interaction between season (pre-denning, denning and post-denning) and year was also included to test whether the effect of season on impala densities depended on the year. Finally, general linear models were used to understand the effect of season on impala densities for each individual year.
Impala density inside and outside the denning ranges during the denning season
Linear mixed models were used to test the hypothesis that wild dog denning causes prey to move away from the denning home range during the denning period. We assessed whether impala density during the denning period varied depending on whether it was inside the denning home range or outside the denning home range. Year was included as a random effect and we also included the interaction between place (inside or outside denning home range) and year to test whether the effect of place on impala densities depended on the year. General linear models were later used to understand the effect of place on impala densities for each individual year.
Impala density inside and outside the denning ranges in the three seasons
To test the hypothesis that wild dogs deliberately den in areas of low prey abundance we calculated the interaction between place (inside and outside denning area) and season (pre-denning, denning, post-denning). The changes in impala density inside and outside denning areas before, during and after denning were assessed so as to test whether impala densities remained lower inside the denning home range than outside throughout the three seasons. Statistical analyses were performed in R (version 2.15.1) (R Development Core Team 2012), all results are reported as mean ± 95 % confidence interval with a significance level of 5 %.
Results
Wild dog home ranges
The non-denning home ranges (September–May) for six pack years were on average 453 km2 (SD = 208 km2; range = 153–717 km2; median = 446 km2). The denning home range (June–August) for six pack years were on average 90 km2 (SD = 52 km2; range = 45–180 km2; median = 69 km2) which is a 5.3 km radius from the den (minimum 3.8 km, maximum of 7.5 km). Home range sizes during the denning seasons were on average smaller than during the non-denning seasons (T = −4.14; d.f = 5.61; p = 0.007).
Impala densities in the pre-denning, denning and post-denning season
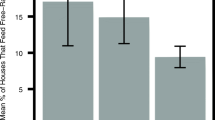
In general, when averaged across all years wild dog denning had no effect on impala densities within the denning home ranges. Impala densities did not differ between the pre-denning (17.80 ± 0.35 impala/km2); denning (18.03 ± 0.33 impala/km2) and post-denning season (17.66 ± 0.28 impala/km2) (F = 0.17; d.f = 2; p = 0.85). The effect of season on impala density depended on the year (F = 3.55; d.f = 2; p = 0.03). In six out of 7 years there was no significant difference in impala densities between the pre-denning, denning and post-denning seasons while 2010 had a significant difference, with impala densities being lowest in the pre-denning season (Fig 2).

Impala densities inside the denning home range for the pre-denning, denning and post-denning seasons in the Sango Ranch, Savé Valley Conservancy, Zimbabwe from 2004 to 2010
Impala density inside and outside the denning ranges during the denning season
During the denning period, impala density was significantly higher outside the denning home range (18.19 ± 0.22 impala/km2) than inside (16.83 ± 0.35 impala/km2) (F = 6.21; d.f = 1; p = 0.01) when averaged across all years. The effect of place on impala density slightly depended on year (F = 3.60; d.f = 1; p = 0.058). In three out of 7 years impala density was significantly higher outside the denning home range than inside, while there was no significant difference during the 2004, 2005, 2007 and 2008 denning seasons (Fig 3).

Impala densities inside and outside the denning home ranges, during the denning season, in Sango Ranch, Savé Valley Conservancy, Zimbabwe from 2004 to 2010
Impala density inside and outside the denning ranges in the three seasons
The effect of place on impala density did not depend on whether it was the pre-denning, denning or post-denning season (F = 0.4808; d.f = 2; p = 0.6184). Impala densities generally remained lower inside the denning ranges than outside for all seasons: pre-denning, denning and post-denning (Fig 4).

Impala densities inside and outside the denning home range during the denning, post-denning and pre-denning seasons, in Sango Ranch, Savé Valley Conservancy, Zimbabwe averaged across 2004 to 2010
Discussion
Wild dogs are the least popular large carnivores among game ranchers because of their perceived impact on wild ungulate populations (Lindsey et al. 2005). Contrary to the belief by some ranchers that wild dogs cause a local reduction in density of wild ungulates during the denning season (Fanshawe et al. 1991), this study has shown that wild dogs do not cause a significant depletion of prey around their den sites, nor do they drive away prey from the denning range. Results from our study also support earlier findings that wild dogs select dens in areas of lower prey density, possibly as a way of avoiding encounter with lions (Mills and Gorman 1997; Creel and Creel 2002).
The presence of a wild dog den had no effect on impala densities during the denning period, which is contrary to the perception that wild dogs deplete prey during the denning period (Fanshawe et al. 1991). This study found no difference in impala densities between the denning season and the pre-denning and post-denning seasons, within denning home ranges. A decrease in impala density during the denning season might have been explained by the fact that impala might (1) have been depleted by wild dog predation or (2) have moved out of the denning home ranges to avoid contact with wild dogs. Consequently given that impala density in the denning season did not differ from the pre-denning season we can conclude that (1) wild dogs do not deplete prey during the denning season and (2) impala could not have suddenly moved out during the denning season, also evidenced by the fact that there was no sudden increase in impala density during the post-denning season. Therefore the presence of denning wild dogs has no direct effect on impala density or movements.
There were less impala within wild dog denning home ranges than outside the denning home ranges during the denning period and this could be explained by the fact that (1) wild dogs were causing a reduction in impala during denning (2) impala moved out of the denning home range during the denning period or (3) wild dogs were selecting denning in areas of low impala density. The first and second explanations are not supported by our study as explained above. However the third explanation is supported by our study as impala densities remained lower inside the denning home range than outside, throughout all seasons (pre-denning, denning and post-denning). Earlier studies have also shown wild dogs to select habitats with low densities of their preferred prey so as to avoid contact with lions (Mills and Gorman 1997; Creel and Creel 2002). Throughout wild dogs’ range (Woodroffe et al. 2004) and indeed in SVC, lions are a major cause of wild dog mortality. In SVC lion numbers have been increasing rapidly since 2005 and pup predation by lion at den sites has been observed on several occasions (Mbizah et al. 2012). One of the main tactics available to wild dogs to decrease mortality from lions is spatial avoidance by selecting areas with lower lion densities (Creel and Creel 2002). These tend to be the areas where ungulate density is lower, because lions will monopolise the prime, prey-rich, areas (Mills and Gorman 1997; Creel and Creel 2002). Nonetheless, the long-term impact of wild dog predation on the local scale may be higher if wild dogs choose to locate their den in the same area year after year, which may be the case in smaller reserves and ranches.
Wild dogs have very little effect on prey, beyond what they actually consume. Some studies have suggested that even if wild dogs do deplete prey, they mainly select for animals in poor condition (Pole et al. 2004; Davies-Mostert et al. 2013), and subsequently, a portion of predation by wild dogs removes sick animals that would have died anyway (compensatory predation) (Lindsey et al. 2005). However, wild dogs’ impact on ungulate populations would also be expected to be greater inside smaller, fenced reserves, where populations are enclosed and where wild dogs can use the fences as an aid when hunting (van Dyk and Slotow 2003). Fence hunting increases the effect of wild dogs on prey by enabling them to capture prey of different species, age classes, sex, body condition and size than would otherwise be taken (van Dyk and Slotow 2003; Davies-Mostert et al. 2013). However, the removal of internal fences within the SVC in the early 90s (Pole 1999) reduces the chances of wild dogs using fences for hunting prey thereby minimising their effect on prey. Consequently many perceived problems associated with carnivores on private land are reduced in conservancies such as SVC where fences are absent, where there are larger prey populations, and where landowners benefit from economies of scale which make them more financially profitable and robust, and also more economically efficient (Barnes and De jager 1996). Therefore encouraging the formation of conservancies among private landowners should be a priority to facilitate coexistence between landowners and predators (Lindsey et al. 2005).
Conservation implications
The long-term survival of wild dogs and success of future conservation efforts in private land depends on the willingness of landowners to tolerate their presence. Although negative attitudes towards wild dogs by ranchers may take time to change, it is believed that research studies aiming at establishing factual evidence about wild dogs will influence the current inaccurate perceptions pertaining today. Awareness programs for both ranchers and communities outside protected areas highlighting the economic, ecological and social benefits of wild dogs and other large carnivores also assists with decreasing misperceptions and forms an important strategy for promoting carnivore conservation. Generating benefits from wild dog conservation such as through eco-tourism and promoting the tourist value of wild dogs is another way in which attitudes towards wild dogs might be improved. Apart from studies such as ours that demonstrate no significant impact of wild dogs on prey, we believe that the negative attitudes of ranchers towards wild dogs can be further reduced if researchers can demonstrate that a sustainable profit could be derived from having wild dogs on ranches. This will probably increase the number of ranchers willing to have wild dogs on their land. The information from this study is crucial in allowing managers and ranchers to make informed decisions about wild dog management. However, it is not clear to what extent our findings can be extrapolated to other areas. Therefore there is a need for more studies on wild dogs’ true impact on ungulate populations, in other ecosystems.
References
Barnes JI, De jager JLV (1996) Economic and financial incentives for wildlife use on private land in Namibia and the implications for policy. S Afr J Wildl Res 26:37–46
Childes SL (1985) The past history, present status and distribution of the hunting dog Lycaon pictus in Zimbabwe. Biol Conserv 44:301–316
Creel S, Creel NM (2002) The African wild dog: behaviour, ecology and conservation. Princeton University Press, Princeton
Davies-Mostert HT, Mills MGL, Macdonald DW (2013) Hard boundaries influence African wild dogs’ diet and prey selection. J Appl Ecol 50:1358–1366
du Toit RF (1994) The Savé Valley Conservancy. In: Booth V, de la Harpe D (eds) The Lowveld conservancies: new opportunities for productive and sustainable land-use. Price Waterhouse, Zimbabwe, pp 28–36
Fanshawe JH, Frame LH, Ginsberg JR (1991) The wild dog—Africa’s vanishing carnivore. Oryx 25:137–146
Gorman ML, Mills MGL, French J (1992) Satellite tracking of the African wild dog Lycaon pictus. In: Priede IG, Swift SM (eds) Wildlife telemetry: remote monitoring and tracking of animals. Ellis Horwood, London, pp 219–228
Hemson G, Johnson P, South A, Kenward R, Ripley R, Macdonald D (2005) Are kernels the mustard? Data from global positioning system (GPS) collars suggests problems for kernel home-range analyses with least-squares cross-validation. J Anim Ecol 74:455–463
Hunter L (2011) Carnivores of the world. Princeton University Press, Princeton
Joubert C (2013) A natural resource assessment of Chapungu Ranch, Savé Valley Conservancy, Zimbabwe. MSc dissertation, University of Pretoria
Kellert SR (1985) Public perceptions of predators, particularly the wolf and coyote. Biol Conserv 31:167–189
Lindsey PA, du Toit JT, Mills MGL (2004) Area and prey requirements of African wild dogs under varying habitat conditions: implications for reintroductions. S Afr J Wildl Res 34:77–86
Lindsey PA, du Toit JT, Mills MG (2005) Attitudes of ranchers towards African wild dogs Lycaon pictus: conservation implications for wild dogs on private land. Biol Conserv 125:113–121
Lindsey PA, Roulet PA, Romañach SS (2007) Economic and conservation significance of the trophy hunting industry in sub-Saharan Africa. Biol Conserv 134:455–469
Malcolm JR, Marten K (1982) Natural selection and the communal rearing of pups in African Wild Dogs (Lycaon pictus). Behav Ecol Sociobiol 10:1–13
Mbizah MM, Marino J, Groom RJ (2012) Diet of four sympatric carnivores in Savé Valley Conservancy, Zimbabwe: implications for conservation of the African wild dog (Lycaon pictus). S Afr J Wildl Res 42:94–103
Mills MGL (1991) Conservation management of large carnivores in Africa. Koedoe 34:81–90
Mills MGL, Gorman ML (1997) Factors affecting the density and distribution of wild dogs in the Kruger National Park. Conserv Biol 11:1397–1406
Moleo′n M, Sa′nchez-Zapata JA, Gil-Sa′nchez JM, Barea-Azco′n JM, Ballesteros-Dupero′n E, Virgo′s E (2011) Laying the foundations for a human–predator conflict solution: assessing the impact of Bonelli’s Eagle on rabbits and partridges. PLoS ONE 6:e22851
Pole AJ (1999) The behaviour and ecology of African wild dogs, Lycaon pictus in an environment with reduced competitor density. PhD thesis, University of Aberdeen
Pole A, Gordon IJ, Gorman ML, Macaskill M (2004) Prey selection by African wild dogs (Lycaon pictus) in southern Zimbabwe. J Zool Lond 262:207–215
R Development Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria
Rasmussen GSA (1999) Livestock predation by the painted hunting dog Lycaon pictus in a cattle ranching region of Zimbabwe: a case study. Biol Conserv 88:133–139
Romañach SS, Lindsey PA (2008) Conservation implications of prey responses to wild dogs Lycaon pictus during the denning season on wildlife ranches. Anim Conserv 11:111–117
Sillero-Zubiri C, Laurenson MK (2001) Interactions between carnivores and local communities: conflict or coexistence? In: Gittleman JL, Funk SM, Macdonald DW, Wayne RK (eds) Carnivore conservation. Cambridge University Press, Cambridge, pp 282–312
Quantum GIS Development Team (2012) Quantum GIS Geographic Information System. Open Source Geospatial Foundation Project
van Dyk G, Slotow R (2003) The effects of fences and lions on the ecology of African wild dogs reintroduced to Pilanesberg National Park, South Africa. Afr Zool 38:79–94
Woodroffe R, Ginsberg JR (1998) Edge effects and the extinction of populations inside protected areas. Science 280:2126–2128
Woodroffe R, Ginsberg JR (1999) Conserving the African wild dog, Lycaon pictus. I. Diagnosing and treating causes of decline. Oryx 33:132–142
Woodroffe R, McNutt JW, Mills MGL (2004) African wild dog Lycaon pictus. In: Sillero-Zubiri C, Hoffmann M, Macdonald DW (eds) Canids: foxes, wolves, jackals and dogs, IUCN status survey and conservation action plan. IUCN, Cambridge, pp 174–183
Acknowledgments
Wild dog fieldwork was conducted under permits from the Research Council of Zimbabwe, and funded by National Geographic Conservation Trust, Columbus Zoo and Rufford Small Grants Foundation. We would like to thank the Zimbabwe Parks and Wildlife Management Authority for their support, and members of the Savé Valley Conservancy, particularly Sango Ranch for access to their properties. Rueben Bote and Misheck Matari are thanked for assistance in the field.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by David L. Hawksworth.
Rights and permissions
About this article
Cite this article
Mbizah, M.M., Joubert, C.J., Joubert, L. et al. Implications of African wild dog (Lycaon pictus) denning on the density and distribution of a key prey species: addressing myths and misperceptions. Biodivers Conserv 23, 1441–1451 (2014). https://doi.org/10.1007/s10531-014-0675-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-014-0675-9