
Abstract
Terrestrial micromolluscs (snails with an adult maximum shell dimension <5 mm) constitute a considerable proportion of the land snail fauna of the Pacific. However, micromolluscs are often underestimated in biological surveys because of size bias. It has been argued that visual searches are preferable on Pacific islands because: (1) size biases are limited based on the understanding that most native Pacific island land snails are very small, and (2) amount of labor is less than other methods such as soil surveys and adequate for inventory purposes (though not for abundance assessments). To test whether visual surveys and soil surveys were accurately recording all taxa, land snail inventories were completed in three forest reserves (5 sampling sites in each) on the island of Oahu, Hawaii. Visual surveys involved 30-min visual search in a 10 m2 site; soil surveys involved sieving leaf litter and topsoil from four 0.3 m2 quadrats and extracting snails with the aid of a microscope. The data indicate a size and microhabitat bias associated with both techniques. Visual surveys consistently collected large arboreal and litter-dwelling species but missed a significant portion of micromolluscs, while soil surveys collected micromolluscs but missed larger snails. Because of such biases, employing both methods is critical for collecting all taxa at a survey location. As such, we recommend that future land snail surveys on Pacific Islands incorporate both survey techniques. Obtaining a complete inventory is critical if we are to understand species distributions and patterns of diversity and make well-informed conservation recommendations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The native Hawaiian land snail fauna is highly diverse, with more than 750 species, of which more than 99 % are endemic to the Hawaiian Islands (Cowie et al. 1995). Habitat destruction, impacts of invasive species and other anthropogenic changes (e.g. climate change) have drastically reduced the numbers of extant species (Lydeard et al. 2004). Estimates of extinction are as high as 90 % and the majority of the remaining species are now found only at higher elevations where human impact is less than at lower elevations (Solem 1990; Cowie 2001a). However, insufficient empirical data have been acquired to estimate this loss of biodiversity accurately, as is the case for land snails worldwide (Régnier et al. 2009). Effective conservation relies on accurate knowledge of the remaining species, which requires extensive survey work that uses efficient, rigorous and comprehensive sampling methods (Cameron and Pokryszko 2005).
Terrestrial micromolluscs (snail species with an adult maximum shell dimension of <5 mm) constitute a considerable proportion of the snail fauna in many locations and habitats (Liew et al. 2008). For example, in eastern North America they represent almost 75 % of land snail species (Nekola 2005), on the island of Bali 55 % (Vermeulen and Whitten 1998) and in one location in New Zealand 88 % (Barker and Mayhill 1999). However, because of their small size and for many species their soil and leaf litter habitat, micromolluscs are often under-sampled in biological surveys (Menez 2001; Rundell and Cowie 2003; Cameron and Pokryszko 2005). Of the 752 taxonomically valid land snail species in Hawaii, excluding Ellobiidae, taxa of unknown systematic placement and taxa that may not be Hawaiian, as explained by Cowie (1995), ~26 % are <5 mm; 32 % 6–10 mm and 42 % >10 mm. Among Hawaii’s ten native families of land snails, more than 50 % of the species in Endodontidae, Helicinidae and Zonitidae are micromolluscs. Three of the families, Hydrocenidae, Punctidae and Pupillidae, are composed entirely of micromolluscs. These assessments are based on measurement of type material and information from the literature.
Two methods are commonly used for sampling land snails: (1) visual surveys—timed visual searches in the field that quantify person survey hours by recording the number of people and the time each area is surveyed; and (2) soil surveys—collection of soil with associated leaf litter, either a standard volume or from a standard area, followed by sequential sieving through a series of progressively finer mesh sieves and picking the molluscs from the resulting fractions, under a microscope for the smaller fractions (Emberton et al. 1996; Menez 2001; Cameron and Pokryszko 2005; Nekola 2005; Liew et al. 2008; Fontaine et al. 2011). Recent surveys in Hawaiian habitats have employed timed visual searches, but not soil surveys (e.g. Rundell and Cowie 2003; Cowie et al. 2008; Meyer and Cowie 2010). It has been argued that visual searches alone are preferable in Hawaii and other Pacific islands for land snail species inventory (though less so for assessments of abundance) because with limited resources the far greater number of samples that can be processed and therefore the greater potential geographic coverage outweigh the disadvantages of under-sampling the very small species that are more difficult to see/collect (Cowie 2001b; Rundell and Cowie 2003). It has also been argued that because most native Pacific island land snails are very small, biases related to size are limited (Cowie and Robinson 2003). These methods have been discussed in detail by Cameron and Pokryszko (2005).
However, a rigorous comparison between the two methods has never been conducted in the Pacific. Ultimately, discerning the most effective survey approach for finding micromolluscs, critical for compiling comprehensive inventories, is crucial for understanding species distributions, and developing effective conservation management plans. Our objective in conducting this study was to examine the efficacy of both techniques (visual and soil) to accurately record all species present. Based on the results, a methodology suitable for land snail surveys in Hawaii and other Pacific islands is recommended. Our analyses complement similar analyses of methods used in Madagascar (Emberton et al. 1996) and Europe (Cameron and Pokryszko 2005) and will lead to a more generally accepted and rigorous approach to land snail inventory that will also permit more rigorous comparison among studies in different parts of the world, a need previously identified in particular by Emberton et al. (1999).
Methods
Study sites and sampling strategies
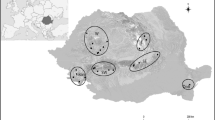
Surveys took place in three forest reserves in the Waianae mountain range of the Hawaiian island of Oahu: Mt. Kaala, Kahanahaiki and Palikea (Fig. 1). These areas harbor native and non-native snails, both macromolluscs and micromolluscs, in a variety of habitats. Sampling took place at each forest reserve from April to August 2011.

Map showing the location of the Hawaiian Islands in the Pacific, with the three surveyed forest reserves on the island of Oahu (numbered stars). From north to south: 1—Kahanahaiki, 2—Mt. Kaala and 3—Palikea
At each forest reserve five 10 m2 sampling sites, spaced at least 100 m apart along a transect, were chosen. Maximizing the number of different microhabitats sampled was the focus. The Palikea sites were visited twice and the Mt. Kaala and Kahanahaiki sites were visited three times. However, only three of the five sites at Kahanahaiki were surveyed on the third visit. At each site, sampling first involved a 2 person, 15 min visual search of all suitable microhabitats (including on plants) by experienced snail collectors. These visual searches were followed by collection of leaf litter and 1 cm of topsoil within 0.3 m2 quadrats from four haphazardly selected areas within each site, including places of deep litter accumulation and around the bases of rocks and trees. All leaf litter and topsoil was concentrated in the field using a 7 mm mesh sieve, with the material that remained in the sieve being visually searched in the field to find any larger snails or slugs (cf. Fontaine et al. 2011). The material that passed through the sieve was brought to the lab for further processing. All specimens collected, using both collection methods, were brought back to the lab for identification, with the exception of any federally listed Achatinella spp. and other species readily identified in the field, which were simply counted. All species were identified to the lowest taxonomic level possible. In some instances it was not possible to refer specimens to a particular species but only to a family, subfamily or genus.
Laboratory processing was completed by separating the concentrated samples through a series of sieves (5, 2, 0.5 mm), which facilitated faster searches and easier removal of molluscs from the different size fractions. Each size fraction was then searched for specimens with the aid of a stereomicroscope. Due to the extensive amount of time necessary to process and sort the soil/litter, it was often not possible to determine if an individual died prior to collection or during storage. As such, live and dead (shell only) counts were pooled. A subset of the live specimens was preserved in 95 % ethanol as representative vouchers and for future genetic analysis. All vouchers were cataloged and deposited in the malacological collections of the Bishop Museum, Honolulu (BPBM 277352-277535, 277543-277603).
Analyses
To determine the relative effectiveness of the visual and soil techniques for collecting all taxa at different scales (i.e. entire forest reserves and sites within forest reserves), we compared richness at each forest reserve and examined differences in the mollusc composition at each site. We also recorded the amount of time necessary for each method to evaluate better the most effective allocation of research efforts for future studies.
Three rarefaction curves were created for each forest reserve using EcoSim 7.0 (Gotelli and Entsminger 2004) to examine patterns of richness as the number of samples increased. Two curves examined patterns of taxonomic richness for each sampling method independently and the other examined patterns of taxonomic richness combining the data from both sampling methods.
To investigate statistically whether the two techniques collected different components of the snail community at each site, we used a permutation-based hypothesis test (ANOSIM analyses) implemented in PRIMER 5.2.9 (Clarke and Gorley 2001). Separate analyses were conducted for each sampling site. Prior to each ANOSIM analysis, a presence/absence matrix of all taxa by site was assembled for each sampling site. A presence/absence matrix was used because the visual search method was aimed at assessing taxonomic richness not relative abundances. Similarity matrices for each forest reserve were created using the Bray–Curtis similarity coefficient (Legendre and Legendre 1983). Because we expected differences in land snail communities among sites within each forest reserve, we used a two-way crossed ANOSIM (9,999 permutations) with sampling technique and sampling site as factors. This allowed us to test for differences between sampling techniques averaged within blocks of sites for each of the three forest reserves, an analysis that is a permutation analogue to a complete randomized block ANOVA. We report both the ANOSIM test statistic (R-values) and the permutation based P values for just the sampling technique factor. The R-statistics are the average rank dissimilarities among and within groups, scaled so that R-values vary between roughly 0 and 1 (there may be some negative values); a value of 0 indicates that there are no differences among treatments, and a value of 1 indicates that all dissimilarities between samples in different treatments are larger than the average dissimilarity among samples within each treatment. The P values test whether R-statistics differ significantly from zero (i.e. that there are significant differences among treatments). To determine the relative contribution of the various taxa to the dissimilarity between the two sampling methods, we also ran a similarity percentage analysis (SIMPER) for each forest reserve using the presence/absence matrices.
We also used ANOSIM tests to examine if the two methods reveal similar patterns of community structure (i.e. do they identify similar land snail community patterns among forest reserves). We ran two ANOSIM tests, one using only the soil data and the other using only the visual data. Analyses again used presence/absence matrices and the Bray-Curtis similarity coefficient. Pair-wise differences among sampling locations were examined only following a significant ANOSIM test and with P values adjusted for multiple testing (3 pairwise comparisons) using a Bonferroni procedure (α = 0.016).
Results
In total, 23 taxa were collected at the three forest reserves (Table 1). A substantial proportion of the taxa collected were found either only using the visual method (Euglandina rosea, Achatinella mustelina, Deroceras laeve, Kaala subrutila, Auriculella spp., Amastra sp., Limax maximus) or only the sieving technique (Helicinidae sp., Pronesopupa spp., Paralaoma servilis). In general the species collected using the visual technique were either large ground-dwelling species (e.g. Deroceras laeve, Kaala subrutila) or arboreal species (Achatinella mustelina), while those collected using the soil technique were primarily ground-dwelling micromolluscs. No snails or slugs were found in the material retained by a 7 mm sieve when collecting soil and leaf litter in the field.
The rarefaction curves indicate that using both survey techniques is the only way to insure that a complete inventory of the species present is obtained at a sampling location (e.g. a forest reserve). For example, at Mt. Kaala, combining the two methods led to recording higher taxonomic richness than using either method alone (Fig. 2). The visual method did not record Paralaoma servilis, Helicinidae spp., Pronesopupa spp. or Allopeas clavulinum, while the soil method did not recover Deroceras laeve, Kaala subrutila or Auriculella spp. (Table 1). Conversely, the rarefaction curves for Kahanahaiki and Palikea (though not Kaala) indicate that given enough effort (>8 samples) both soil and visual search techniques will collect approximately the same level of diversity and collect the majority of the taxa present at a forest reserve (Fig. 2). The rarefaction curves also indicate that many fewer sites can be visited in a sampling location (approx. 4–5 sites) if both techniques are employed at each site.

Rarefaction curves of taxonomic richness as the number of sampling sites increases using either the soil or visual technique or both techniques combined for the three sampling locations
ANOSIM analysis revealed that the land snail assemblages collected by the two sampling techniques differed among sites within all three forest reserves: Mt. Kaala (R = 0.553, P = 0.003), Kahanahaiki (R = 0.562, P = 0.006), and Palikea (R = 0.461, P = 0.03). SIMPER analysis showed that different taxa were collected with differing regularity using the two survey techniques (Table 1). Micromolluscs were more often collected in soil surveys while larger molluscs (ground-dwelling and arboreal) were collected more often in visual surveys.
The ANOSIM analyses for each of the two sampling techniques identified significant differences in land snail communities among forest reserves (soil, R = 0.387, P = 0.001; visual, R = 0.19, P = 0.02). Using the visual technique only we found significant differences among all forest reserves, while using only the soil technique significant differences were found only between Kahanahaiki and Mt. Kaala and between Kahanahaiki and Palikea but not between Mt. Kaala and Palikea (Table 2).
Discussion
The soil technique has not been used in most recent land snail surveys in the Hawaiian and Samoan islands (e.g. Cowie and Robinson 2003; Cowie et al. 2008; Meyer and Cowie 2010). This is because it has been suggested that biases related to size are limited, because most native Hawaiian and Pacific island land snails are very small (Cowie and Robinson 2003), meaning that since surveyors are focused on small snails, they are unlikely to miss small snails. Our results contradict this assumption. We have demonstrated that there is a size and habitat bias associated with both visual and soil sampling techniques. Timed visual surveys consistently find large arboreal and litter-dwelling species but miss a significant portion of micromolluscs, while soil surveys find micromolluscs but miss larger snails, particularly arboreal snails. For example, the soil collection method failed to find arboreal snails like A. mustelina and large litter-dwelling slugs like D. laeve. This is probably due to the limited area surveyed using the soil/litter collection technique, which results in missing these larger, less abundant species. Other studies have found that large species and slugs are often missed in soil surveys (Emberton et al. 1996; Cameron and Pokryszko 2005; Fontaine et al. 2011). Also, the extended processing and storage time associated with the soil/litter collection technique may have led to the slugs decomposing (Cameron and Pokryszko 2005), as slugs were documented in subsequent sampling that included storing the soil/litter at 4 °C during the laboratory processing phase.
Due to these biases, employing both methods is critical for collecting all taxa at a sampling location, and is especially important for collecting all taxa at sites within a sampling location. While rarefaction curves for Kahanahaiki and Palikea suggest that employing either method with sufficient effort (>8 sites) may result in the collection of most taxa, both the rarefaction curve for Mt. Kaala and the SIMPER results indicate that efforts that only employ one technique will often miss taxa (e.g. Pronesopupa spp. and Succineidae spp. at Palikea, despite both species being relatively common). Most micromollusc taxa were commonly found at sites when soil surveys were used, but were rare in visual surveys, and the opposite pattern was found for large arboreal mollusc species.
The soil sampling technique is not often used because of the labor required. The time required to collect and sort the soil samples in this study was 4–8 h per site. This is in sharp contrast to the 1–3 h it took for the visual survey method. Neither estimate includes the time to identify specimens, nor travel time, which would increase dramatically with the number of sites. While the additional steps involved in the laboratory processing of the soil increased the time it took to complete a soil survey compared to a visual survey, fewer sites may need to be surveyed if both techniques are employed. For example, based on the rarefaction curves and average time per site for each method (2 h visual, 6 h soil), at Mt. Kaala, collecting at just six sites using both techniques would yield more species with 48 h of effort than sampling at 14 sites using just one of the techniques, which would require 28 h (visual) or 84 h (soil). While four times as many sites could be surveyed visually in the time needed to complete six sites using both methods, there is no guarantee of recovering the same numbers of species and the added travel time would probably be prohibitive.
Determining whether or not to employ both methods in surveys will also depend on the question being addressed. For example, to examine possible differences in snail assemblages among sampling locations one technique may be sufficient. In this study both soil and visual surveys identified differences in the land snail assemblages among most sampling locations. Soil surveys identified no difference between Mt. Kaala and Palikea, while visual surveys did, although the difference was only marginally significant (P = 0.014) following Bonferroni correction (α = 0.016). This suggests that using either technique will yield similar results. Which technique will be more sensitive for addressing such questions will probably depend on both the areas being surveyed and the taxonomic level to which the specimens are identified. For instance, if the Helicarionidae spp. were identified to the species level, we may have seen significant differences among the Mt. Kaala and Palikea sampling locations in the soil survey results. Using only one sampling technique may reveal differences among sampling locations, but using a single technique will often result in an incomplete inventory of the mollusc fauna at a given location. Similarly, single technique approaches may not allow a clear understanding of the distribution of all species in an area, because of the size and habitat biases of the different techniques.
Although both methods can provide quantitative assessments (using area or volume for soil/litter, time for visual), the direct comparability among researchers and across studies arguably make soil/litter sampling more objective and therefore a key component of land snail biodiversity surveys. The importance of implementing the soil/litter collection methodology in Hawaiian land snail biodiversity surveys was made especially clear by the novel recording of Paralaoma servilis on Oahu during this study (Christensen et al. 2012). Despite being widespread, this species was not found in the multiple visual surveys that have been conducted in the Waianae mountain range prior to this study. In the current study, it was missed entirely in visual surveys and only discovered, often widespread and abundant, in the soil surveys.
As such, we recommend that future land snail surveys on Pacific Islands, especially Hawaii, incorporate both survey techniques. Using both techniques reduces biases related to size and microhabitat (e.g. arboreal species missed by soil/litter collecting). Micromolluscs constitute a significant proportion of the Hawaiian and Pacific fauna and to assess their conservation status adequately requires comprehensive surveys employing soil surveys (e.g. Gargominy 2008; Sartori et al. 2013). If researchers are reluctant to employ the soil technique because of the time required to sort through the remaining soil and leaf litter to find the smaller snails, we suggest reducing the number of sites visited but still employing both techniques. We make this recommendation because it is the only way to obtain a complete inventory of the species present at a site, which is critical if we are to understand patterns of diversity across a landscape and make species-specific conservation recommendations based on their distributions.
References
Barker GM, Mayhill PC (1999) Patterns of diversity and habitat relationships in terrestrial mollusc communities of the Pukeamaru Ecological District, northeastern New Zealand. J Biogeogr 26:215–238
Cameron RAD, Pokryszko BM (2005) Estimating the species richness and composition of land mollusc communities: problems, consequences and practical advice. J Conchol 38:529–547
Christensen CC, Yeung NW, Hayes KA (2012) First records of Paralaoma servilis (Shuttleworth 1852) (Gastropoda: Pulmonata: Punctidae) in the Hawaiian Islands. Bishop Mus Occas Pap 112:3–7
Clarke KR, Gorley RN (2001) V.5 users manual/tutorial PRIMER-E. PRIMER-E Ltd, Plymouth
Cowie RH (1995) Variation in species diversity and shell shape in Hawaiian land snails: in situ speciation and ecological relationships. Evolution 49:1191–1202
Cowie RH (2001a) Invertebrate invasions on Pacific islands and the replacement of unique native faunas: a synthesis of the land and freshwater snails. Biol Invasion 3:119–136
Cowie RH (2001b) Decline and homogenization of Pacific faunas: the land snails of American Samoa. Biol Conserv 99:207–222
Cowie RH, Robinson AC (2003) The decline of native Pacific island faunas: changes in status of the land snails of Samoa through the 20th century. Biol Conserv 110:55–65
Cowie RH, Evenhuis NL, Christensen CC (1995) Catalog of the native land and freshwater molluscs of the Hawaiian Islands. Backhuys Publishers, Leiden
Cowie RH, Hayes KA, Tran CT, Meyer WM III (2008) The horticultural industry as a vector of alien snails and slugs: widespread invasions in Hawaii. Int J Pest Manag 54:267–276
Emberton KC, Pearce TA, Randalana R (1996) Quantitatively sampling land-snail species richness in Madagascan rainforests. Malacologia 38:203–212
Emberton KC, Pearce TA, Randalana R (1999) Molluscan diversity in the unconserved Vohimena and the conserved Anosy mountain chains, southeast Madagascar. Biol Conserv 89:183–188
Fontaine B, Gargominy O, Prié V (2011) Indigenous land snails. In: Bouchet P, Le Guyader H, Pascal O (eds) The natural history of santo. Muséum national d’Histoire naturelle, Paris, pp 169–178
Gargominy O (2008) Beyond the alien invasion: a recently discovered radiation of Nesopupinae (Gastropoda: Pulmonata: Vertiginidae) from the summits of Tahiti (Society Islands, French Polynesia). J Conchol 39:517–536
Gotelli NJ, Entsminger GL (2004) EcoSim: null models software for ecology. Version 7.0. Acquired Intelligence Inc. & Kesey-Bear. http://www.together.net/~gentsmin/ecosim.htm. Accessed 3 June 2013
Legendre L, Legendre P (1983) Numerical ecology: developments in environmental modelling, 3rd edn. Elsevier, Amsterdam
Liew T-S, Clements R, Schilthuizen M (2008) Sampling micromolluscs in tropical forests: one size does not fit all. Zoosymposia 1:271–280
Lydeard C, Cowie RH, Ponder WF, Bogan AE, Bouchet P, Clark S, Cummings KS, Frest TJ, Gargominy O, Herbert DG, Hershler R, Perez K, Roth B, Seddon M, Strong EE, Thompson FG (2004) The global decline of nonmarine mollusks. Bioscience 54:321–330
Menez A (2001) Assessment of land snail sampling efficacy in three Mediterranean habitat types. J Conchol 37:171–175
Meyer WM III, Cowie RH (2010) Invasive temperate species are a threat to tropical island biodiversity. Biotropica 42:732–738
Nekola JC (2005) Geographic variation in richness and shell size of eastern North American land snail communities. Rec W Aust Mus Suppl 68:39–51
Régnier C, Fontaine B, Bouchet P (2009) Not knowing, not recording, not listing: numerous unnoticed mollusk extinctions. Conserv Biol 23:1214–1221
Rundell RJ, Cowie RH (2003) Preservation of species diversity and abundances in Pacific island land snail death assemblages. J Conchol 38:155–169
Sartori AF, Gargominy O, Fontaine B (2013) Anthropogenic extinction of Pacific land snails: a case study of Rurutu, French Polynesia, with description of eight new species of endodontids (Pulmonata). Zootaxa 3:343–372
Solem A (1990) How many Hawaiian land snail species are left? and what we can do for them. Bishop Mus Occas Pap 30:27–40
Vermeulen JJ, Whitten AJ (1998) Fauna Malesiana: guide to the land snails of Bali. Backhuys Publishers, Leiden
Acknowledgments
We thank Dylan Ressler, Patrick Curry, David Sischo, Jaynee Kim, Kelsey Coleman, Taryn Takebayashi, Celeste Yee, Vince Costello and Stephanie Joe for assistance in the field and lab. T.H. Durkan and N.W. Yeung were supported by an Oahu Army Natural Resource Program grant (PI Yeung). This work was also supported by National Science Foundation grant DEB-1120906 to K.A. Hayes and a University of Hawaii Undergraduate Research Opportunities Program grant to T.H. Durkan. We thank the Division of Forestry and Wildlife (Department of Land and Natural Resources) for permits.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Durkan, T.H., Yeung, N.W., Meyer, W.M. et al. Evaluating the efficacy of land snail survey techniques in Hawaii: implications for conservation throughout the Pacific. Biodivers Conserv 22, 3223–3232 (2013). https://doi.org/10.1007/s10531-013-0580-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-013-0580-7