
Abstract
Wind energy development is rapidly increasing within breeding and wintering ranges of many grassland birds in North America. Despite recognized environmental benefits of such development, wind farms have the potential to negatively impact bird communities. Using an area-search method, we surveyed grassland birds within a matrix of pastures, hay fields, and agricultural lands at a wind facility in north-central Texas during the winters of 2009/10 and 2010/11. We used binary logistic regression to examine the effect of distance from wind turbines on plot occupancy for species that did not show significant habitat associations. In contrast, for species with significant habitat associations, we used a two-step process to test for displacement. First, we identified suitable plots using a niche modeling approach in Maxent. Second, we used binary logistic regression to examine whether distance to turbine influenced the occupancy of those plots. Sprague’s Pipit Anthus spragueii, Savannah Sparrow Passerculus sandwichensis, and meadowlarks Sturnella sp. showed no evidence for displacement. In contrast, the Le Conte’s Sparrow Ammodramus leconteii, was significantly more likely to occupy suitable plots as distance from the nearest turbine increased. Mean plot occupancy for the Le Conte’s Sparrow was more than four times lower in plots <200 m from the nearest wind turbine compared to those that were >400 m away. Our study highlights the need to investigate displacement at the level of individual species. Our data also suggest that species dependent on cryptic predator evasion strategies may be displaced from wind turbines and this idea warrants further investigation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Grassland birds are the most rapidly declining group of birds in North America (Knopf 1994; Peterjohn and Sauer 1999; Brennan and Kuvlesky 2005). Although most research and conservation efforts for this threatened group have focused on the breeding season (Igl and Ballard 1999; Vickery and Herkert 2001), some migratory populations are thought to be limited during the non-breeding season (Macias-Duarte and Panjabi 2013). For example, the quality of wintering habitat can influence the timing and condition of birds arriving at their breeding grounds, two major determinates of reproductive success (Marra et al. 1998). Furthermore, wintering grounds for North American grassland birds have undergone major land use changes that threaten their viability as suitable habitat (Igl and Ballard 1999; Butler et al. 2009; Macias-Duarte et al. 2009). For example, the coastal prairie ecosystem in Texas and Louisiana has one of the highest diversities of wintering grassland birds in North America (Rich et al. 2004), but less than 1 % remains in its original condition (Diamond and Smeins 1984). Habitat for wintering grassland birds has been destroyed by urbanization and intensely produced agriculture, and degraded by modification of grazing patterns, fire suppression, and interference with other ecological drivers that maintain open grasslands (reviewed in Askins et al. 2007).
The development of wind facilities represents another source of habitat loss for grassland birds as habitat is destroyed and fragmented during the construction of wind turbines and related infrastructure (Pearce-Higgins et al. 2012). Furthermore, several studies have shown that some grassland birds exhibit avoidance behavior near tall human-made structures after the construction period (e.g. wind turbines and power lines; Leddy et al. 1999; Pearce-Higgins et al. 2009, 2012; Pruett et al. 2009), suggesting that merely the presence of a tall structure like a utility-scale wind turbine could degrade habitat to such an extent that it would be avoided or abandoned by sensitive species. This type of habitat loss, i.e. displacement, is often considered to be a greater threat to bird populations than collision fatalities (reviewed in Drewitt and Langston 2006; Kuvlesky et al. 2007).
Overall, wind turbine displacement studies in grassland bird communities are rare, especially given the extent of wind energy development in this habitat type. Furthermore, no study to date has explored the mechanisms behind displacement. Leddy et al. (1999), the only displacement study in North America published thus far, found that the density of grassland passerines increased at distances >180 m from the nearest turbine string in conservation reserve program grasslands in southwest Minnesota; however, this study did not control for potential habitat differences near and far from turbines or explore species-specific effects. Pearce-Higgins et al. (2009) found displacement extending 100–800 m from turbines in seven of 12 species studied within a United Kingdom upland breeding bird community. Revisiting these results, Pearce-Higgins et al. (2012) found that some species recovered after the initial construction period, whereas other did not. In contrast, Devereux et al. (2008), the only study of a non-breeding bird community, found no displacement in four functional groups of wintering farmland birds in the United Kingdom. The results from these three studies suggest that (1) displacement is species-specific, i.e. the sensitivity of species to turbines varies (supported by Pearce-Higgins et al. 2009); (2) winter bird communities may respond differently to wind turbines than breeding bird communities (supported by Leddy et al. 1999; Devereux et al. 2008; Pearce-Higgins et al. 2009); and (3) displacement may depend in part on the extent of habitat modification prior to construction of the wind farm; e.g. bird communities in virgin grasslands may be more susceptible to displacement than bird communities in grassland landscapes modified for agricultural production (supported by Pearce-Higgins et al. 2009).
To successfully mitigate for habitat loss and degradation due to wind energy development, there is a need to understand species-specific displacement and the mechanisms behind this displacement. To address this need, we investigated habitat use and displacement from wind turbines in a wintering grassland bird community in north-central Texas. The results of this study should be especially insightful as Texas is a biodiversity hotspot for wintering grassland birds in North America (Rich et al. 2004), providing wintering habitat for species that breed in a variety of grassland types, including tall grass prairie (e.g. Le Conte’s Sparrow Ammodramus leconteii), mixed grass prairie (e.g. Sprague’s Pipit Anthus spragueii), and short grass prairie (e.g. McCown’s Longspur Rhynchophanes mccownii). Furthermore, Texas has more installed wind capacity than any other state in the U.S. (12,212 MW as of December 2012, www.windpoweringamerica.gov) and the second highest installed wind capacity per km2 of any state (0.018 MW/km2; National Renewable Energy Laboratory).
Methods
Study area
We conducted field work at Wolf Ridge Wind, LLC (33°43′53.538″N, 97°24′18.186″W) in the cross timbers and prairies ecoregion of north-central Texas (Griffith et al. 2007). The wind resource area consists of seventy five 1.5 MW General Electric wind turbines (hub height = 80 m, blade length = 42 m) extended over 48 km2. This area has a diversity of land uses including cattle grazing (pastures), native hay harvesting, and winter wheat Triticum aestivum cultivation (agricultural fields). These land use practices varied both spatially and temporally, as landowners often divided their property into a number of small fields with different uses, and changed uses within fields from year to year. This diversity, as well as differences in harvest rates and grazing pressure, created a mosaic of parcels with contrasting vegetation communities. Pastures were moderately to heavily disturbed based on the grazing pressure within each field. Heavily grazed fields were more likely to contain monocultures of invasive plant species (Bermuda grass Cynodon dactylon and King Ranch bluestem Bothriochloa ischaemum) and large forbs (Amphiachyris dracunculoides and Ambrosia psilostachya), whereas moderately grazed fields contained more native grasses. Fields that were harvested for hay or left fallow experienced the lowest levels of disturbance of any field type, and were dominated by native grasses (e.g. Andropogon gerardii, Panicum virgatum, and Sorghastrum nutans). Winter wheat fields represented the most disturbed field type at Wolf Ridge, with poor ground cover and low plant species diversity.
Survey methods
We used an area-search method (Roberts and Schnell 2006) to survey grassland birds at Wolf Ridge from 18 December 2009 to 14 February 2010 and from 29 December 2010 to 23 February 2011. This method relies on flushing birds within 1 ha search plots and bypasses the creation of a distance based detection function required for other common survey methods (e.g. transects and point counts) which can be problematic for cryptic wintering grassland birds or species that occur at low densities (Roberts and Schnell 2006). One to two searchers navigated the 100 by 100 m square plots with a handheld sub-meter Trimble GeoXH® GPS unit. Moving at a 20 min/ha pace, the searchers first walked the perimeter of each plot and then walked four 60 m lines inside the plot, bringing them within 10 m of every point in the plot (Roberts and Schnell 2006). We used ESRI’s ArcGIS (version 9.3; Redlands, California, USA) to randomly locate plots at least 100 m from the forest edge and each other within each field season. As plots were generated independently each year, some plots overlapped between years. Plots were located 17–1,423 m from the nearest wind turbine (measured from the center of the plot). Following a standard protocol for surveying non-breeding grassland birds, we searched all plots between 1 h after dawn and 1 h before dusk (0800–1600 CST) to avoid increases in activity and detectability as birds moved to and from roost sites (Grzybowski 1982; Roberts and Schnell 2006; Devereux et al. 2008). Within the daylight hours, the order in which plots were surveyed was randomized with respect to distance to wind turbine to avoid potential biases of time of day on detection.
During searches, we recorded the number of birds, species present, and the time at which the birds were observed. In each area-search plot, we gathered microhabitat data from a 1 m2 quadrat placed at one random location. We used a random number generator to obtain a direction (0–360°) and a distance (0–50 m) from the center of the plot to generate the random location. We gathered the following microhabitat data in each quadrat: vegetation microhabitat characteristics including floristic composition (% grass, % forb, % tree/shrub, % litter, % bare ground), plant species richness, maximum vegetation height, and the three most common plant species based on the percentage of total ground cover (Knight 1994). We also recorded the presence or absence of fence lines, trees, and bushes as well as the most common plant species (estimated by % ground cover) and land use (grazing, hay field, or agricultural field) within the entire 1 ha plot.
Statistical analysis
To account for habitat associations by birds, and how used and unused habitat was distributed in relation to turbines, we first looked for species-specific habitat associations using χ2 goodness-of-fit tests. Habitat associations were indicated by a non-random occupancy of area-search plots based on field use (pasture, hay, or agricultural field) and the most common plant species within each plot (α = 0.05). We chose these two variables for the χ2 goodness-of-fit tests because they were clearly definable and varied across the wind resource area. For those species with no significant habitat association, we simply used binary logistic regression to explore whether distance to nearest wind turbine influenced the pattern of plot occupancy. In logistic regression, the odds of an event happening (e.g. observing a bird within a plot) increase by eβ for every one unit change in x (Compton et al. 2002); where β is the estimated coefficient and, in our data, x is distance from the nearest wind turbine. We used odds ratios to provide an additional interpretation of logistic regression models (Keating and Cherry 2004). An odds ratio greater than one indicated that the odds of observing a species or species group increased with distance from the nearest wind turbine, whereas an odds ratio less than one indicated that the odds decreased. We used presence data as our response variable because it was more reliable than counts of individual birds for cryptic species, agile species and species that flock in large numbers.
For species with a significant habitat association, we used a two-step process to examine displacement. First, we identified plots representing suitable and unsuitable habitat using a niche modeling approach. Second, we used binary logistic regression to explore whether distance to the nearest turbine influenced the pattern of occupancy in suitable plots. We chose this two-step approach to test for displacement while simultaneously controlling for multiple confounding variables (e.g. vegetation height and field use) that would also influence the distribution of wintering birds. For the first step, we identified Maxent software (v.3.3.3e; Phillips et al. 2006), as the most appropriate ecological niche modeling approach for our data. We selected this program because our search methods did not yield 100 % detection efficiency for all species and therefore our data set included false absences (Roberts and Schnell 2006). These false absences could lead to otherwise suitable plots being classified as ‘unsuitable’ simply because the focal species was cryptic (Pearson et al. 2007; Hirzel and Le Lay 2008). When false absences are likely, it is recommended that presence-only models should be used (Pearson et al. 2006; Hirzel and Le Lay 2008). In addition, some of our study species had small sample sizes (<20 occupied plots). Similar studies show that Maxent (v.3.3.3e; Phillips et al. 2006) has consistently outperformed other ecological niche modeling methods that rely on presence-only data, particularly with small sample sizes (Elith et al. 2006; Hernandez et al. 2006; Pearson et al. 2007; Wise et al. 2008). Using this program, plot characteristics and microhabitat data (see survey methods) from occupied plots were used to create a probability of presence (POP) between 0 and 1 for each plot. To date, Maxent has commonly been applied to studies across continuous landscapes on a broader scale. In our study, we used this approach to identify habitat suitability in randomly located, non-continuous plots on a more local scale, as the different land uses and land use intensities by the various landowners created sharp contrasts in vegetation structure and composition between plots.
We used all occupied plots to generate POP values for species with fewer than 25 presence locations using the default parameters for Maxent (Phillips et al. 2006). To create POP values for species with more than 25 occupied plots, we had Maxent run five replicates, each cross-validated with a different randomly selected 20 % of occupied plots for model testing. We had Maxent generate response curves for the effect of each variable on POP and then performed a jackknife analysis to measure the importance of variables in each model.
In addition, for species with more than 25 occupied plots, we used area under the ROC curve (AUC) validation, a threshold independent statistic that represents the probability that a random presence site will be more suitable than a random background point (Fielding and Bell 1997). This requires separating presence locations into training and testing samples. Separating presence locations is not recommended when sample sizes are limited (n < 25), as training and testing samples become very small (Pearson et al. 2007). Thus, we used a jackknife validation method developed specifically for validating models with small sample sizes (see Pearson et al. 2007 for details). The model’s predictive performance was assessed at two suitability thresholds: minimum training presence (stringent) and fixed cumulative value of 1 (relaxed). For the stringent threshold, only plots with a POP greater than or equal to the lowest POP of an occupied plot were considered suitable. In contrast, the relaxed threshold allowed plots marginally below the minimum POP of occupied plots to be considered suitable. Then, as part of the aforementioned step two, we performed logistic regression using only suitable plots identified using each threshold.
For each species in which logistic regression showed displacement by turbines, we used distance bins and Fisher’s exact tests to look for natural breaks in the rate of plot occupancy across distance to turbine. This was necessary to quantify displacement and make appropriate mitigation recommendations. Finally, for species that showed both habitat associations and displacement by turbines, we also performed a Mann–Whitney U test to compare the distribution of POP values in unoccupied suitable plots (stringent threshold) with POP values in occupied plots. For the latter, we generated a series of POP values by excluding each plot once in turn from model training. We preformed this test to account for the continuous nature of POP, ensuring that plots occupied by such species were not simply plots with the highest POP (i.e. the most suitable) and by chance far away from turbines. Statistical tests were performed in JMP (v. 7.0.2) and Minitab (v.15.1.30).
Results
We surveyed 93 plots in the winter of 2009/2010 and 117 plots in the winter of 2010/2011. Of the 210 plots, 135 were in pasture, 46 were in hay fields, and 29 were in winter wheat fields. The three most common plant species (estimated by total ground cover in a plot) were meadow dropseed (n = 51 plots), winter wheat (n = 27 plots), and Bermuda grass (n = 26 plots). We recorded 25 species of birds in our searches. At least one bird was recorded in 75 % of plots. In total, we made 484 independent observations accounting for 2,115 individuals (Table 1). Solitary birds accounted for 310 observations, whereas the remaining observations were of flocks as large as ca. 250 individuals.
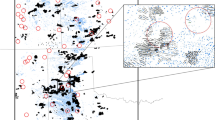
Three species (Sprague’s Pipit, Le Conte’s Sparrow, and Savannah Sparrow Passerculus sandwichensis) and one species group (meadowlarks) had sufficient sample sizes (>15 occupied plots) to test for habitat associations and displacement (Table 1). Eastern Meadowlarks Sturnella magna and Western Meadowlarks Sturnella neglecta were grouped due to difficulties differentiating these species in the field during the non-breeding season. Sprague’s Pipits and meadowlarks did not show habitat associations, whereas Savannah Sparrows and Le Conte’s Sparrows did (Table 2). Thus, we used Maxent to identify suitable plots for these selective species. Average AUC for the five Savannah Sparrow replicates was 0.44 (SE 0.026). We could not use Maxent to differentiate suitable plots from unsuitable plots in this species because the models could not be distinguished from random (AUC = 0.5) at predicting Savannah Sparrow occurrences. In contrast, we found that Maxent had high predictive ability for excluded plots occupied by Le Conte’s Sparrows: 72 % at the stringent threshold and 100 % at the relaxed threshold (P = 0.005, 0.0002 respectively). Vegetation height, field use, and most common plant species were the most important variables in the Le Conte’s Sparrow model. POP values were positively associated with hay fields, increasing vegetation height, meadow dropseed, little bluestem, and tall warm season grasses. POP values were negatively associated with agricultural fields and all other plant species. Mean distance to nearest turbine for plots in which we observed Le Conte’s Sparrows was 497 m (SE 80.2), with the closest occupied plot 182 m from the nearest turbine (Fig. 1a). At the relaxed threshold, Maxent classified 127 plots (60 %) as suitable habitat for Le Conte’s Sparrow, with a mean distance to turbine of 321 m (SE 19.5; Fig. 1b). At the stringent threshold, Maxent classified 86 plots (41 %) as suitable habitat for Le Conte’s Sparrows, with a mean distance to turbine of 338 m (SE 25.7; Fig. 1b). We then performed one logistic regression using the 127 suitable plots at the relaxed threshold and another using the 86 suitable plots at the stringent threshold.

a Le Conte’s Sparrows were observed in 19 of 210 1 ha search plots during the winters of 2009/10 and 2010/11 at Wolf Ridge. There were 21 overlapping plots between the two winters. b Le Conte’s Sparrow probability of presence (POP) as determined by Maxent for each search plot. The warmer colours indicate a higher POP. White crosses indicate wind turbines in both panels. (Color figure online)
Logistic regression showed no evidence that distance to turbine influenced plot occupancy for Sprague’s Pipits, Savannah Sparrows, or meadowlarks (Table 3). In contrast, Le Conte’s Sparrow occupancy increased significantly with distance to turbine, thus the odds of observing a Le Conte’s Sparrow increased by 32 % (relaxed) or 35 % (stringent) for each 100 m increase in distance from turbine (Table 3). For example, under a relaxed threshold the odds of observing a Le Conte’s Sparrow 100–200 m from the nearest turbine were 0.0548. In comparison, at 200–300 m the odds increased to 0.0723. This represents a 32 % increase above 0.0548. The proportion of suitable plots (stringent threshold) occupied by Le Conte’s Sparrows was significantly higher >400 m from the nearest turbine compared to 0–200 m or 200–400 m from the nearest wind turbine (Fisher’s exact tests: P = 0.001, 0.04, respectively; Fig. 2). POP did not differ between occupied and unoccupied suitable plots (stringent threshold; Mann–Whitney U test = 816, n 1 = 19, n 2 = 67, P = 0.92).

Mean (SE) occupancy by Le Conte’s Sparrows in plots that were classified as suitable habitat (stringent threshold) at varying distances from the nearest wind turbine at Wolf Ridge. Range of plots distances >400 m: 408–1,423 m
Discussion
Our study showed that among a community of wintering grassland birds, displacement by wind turbines was species-specific. Sprague’s Pipit, Savannah Sparrow, and meadowlarks showed no evidence for displacement, whereas the Le Conte’s Sparrow exhibited significant evidence for displacement at distances up to 400 m. This study is one of the first to demonstrate species-specific displacement from wind turbines in a North American bird community. Our results pose the question, “why are some wintering grassland birds displaced by wind turbines while others are not?”
When comparing their results with those of Devereux et al. (2008), Pearce-Higgins et al. (2009) hypothesized that species occupying natural or semi-natural habitats would be more sensitive to wind energy development than species occupying intensively produced landscapes. Wolf Ridge provided a microcosm for this hypothesis, as both intensively produced landscapes (e.g. agricultural fields) and semi-natural landscapes (e.g. hay fields dominated by native grasses) were present. Sprague’s Pipits, Savannah Sparrows, and meadowlarks all used a variety of altered habitats, including agricultural fields and heavily grazed fields of non-native monocultures. None of these species showed evidence of displacement (similar to findings by Douglas et al. 2011). In contrast, Le Conte’s Sparrow was limited to hay fields and lightly grazed fields dominated by native vegetation, and showed evidence for displacement. Our results are consistent with the hypothesis proposed by Pearce-Higgins et al. (2009).
Our data also suggest that predator avoidance strategies may be useful for predicting displacement in grassland birds. Birds that forage far from cover (i.e. trees and shrubs), as is common among wintering grassland birds, can avoid predators using one of two strategies (Pulliam and Mills 1977). The first is a social evasion strategy adopted in areas with little cover where increased numbers of individuals assist with predator detection. The second is a cryptic evasion strategy adopted in areas of thick vegetation where a single bird can stay hidden from predators. As social species move in groups and are often strong fliers, they are more likely to use areas in the landscape in which they are exposed to predators. In contrast, cryptic species are often weak fliers that travel alone, and are therefore unlikely to use areas in the landscape where they are exposed to predators (Pulliam and Mills 1977; Grzybowski 1983). If cryptic species perceive wind turbines as a predation risk (i.e. raptor perches), as has been suggested for Greater Sage-Grouse Centrocercus urophasianus and their response to the presence of power lines (Lammers and Collopy 2007), then birds adopting a cryptic evasion strategy may be more likely to avoid areas with wind turbines. The Le Conte’s Sparrow was the only species in our study that exclusively used a cryptic evasion strategy. Furthermore, this species is solitary, has a low flight initiation distance, and is very reluctant to flush from cover (unpublished data). For example, on many occasions we observed individuals running away from searchers rather than taking flight.
To our knowledge, this is the first study to successfully use Maxent to identify habitat suitability in non-continuous plots at a local scale. Because of the substantial differences in vegetation community structure and composition created by different land uses and land use intensities, Maxent was able to differentiate suitable and unsuitable habitat for species that selected habitat based on physical characteristics such as cover availability (e.g. Le Conte’s Sparrow).
Although Savannah Sparrows did show significant habitat associations, Maxent could not model habitat suitability for this species using the variables we measured in our study (i.e. we did not measure all the variables that were important to this species). In support of this, other studies have shown sparrow density to be correlated with seed density in Texas and Oklahoma grasslands (Grzybowski 1983). Although we observed Savannah Sparrows congregating at stands of large forbs, an important source of food for granivorous wintering grassland birds (Smith et al. 2005), we did not quantify the distribution of this resource at our study site. In comparison, we measured habitat characteristics that have been demonstrated to be important to the Le Conte’s Sparrows (e.g. cover availability; Grzybowski 1983), and thus, we were successfully able to model habitat suitability for this species using Maxent.
The U.S. Department of Energy’s goal for wind energy development (20 % of electricity generated by 2030) could be reached with all new construction taking place on already disturbed lands (e.g. cultivated crops and developed land; Kiesecker et al. 2011). Our data, as well as that of Devereux et al. (2008), suggest that siting wind energy facilities in this manner could help reduce indirect effects of wind energy on birds, as species that used cultivated crops as habitat did not show evidence for displacement. Mitigation may be necessary when wind farms are sited in habitat used by species of conservation concern that are susceptible to displacement. Using our data on the Le Conte’s sparrow as an example for a mitigation strategy, 50 ha of grassland should be restored off site for each turbine constructed within suitable habitat (400 m buffer around each turbine). Patterns of plot occupancy and microhabitat use indicated that successfully restored habitat should be dominated by meadow dropseed, little bluestem, and tall warm season grasses, and experience low levels of grazing pressure or haying.
We recommend that studies identify specific species within grassland bird communities that are sensitive to displacement by wind turbines. Our study may indicate that cryptic evasion strategies and reliance on natural or semi-natural habitat may be indicators of sensitivity. For example, other species from the genus Ammodramus use cryptic strategies to evade predators (Dunning 2001), and more than half of the species from this genus are of conservation concern (Rich et al. 2004). We recommend that future studies initially target these species. In addition, such studies should use rope dragging or active flushing strip transects that increase detection rates for species that do not readily flush (Fletcher et al. 2000; Twedt et al. 2008). Furthermore, recent large scale wind energy development on Texas’s Gulf Coast overlaps with wintering and breeding ranges for multiple Ammodramus species (Kuvlesky et al. 2007), and could therefore represent significant habitat loss or degradation for these birds.
Our results show different responses to wind turbines within one bird family and should caution others to not group species for analysis. Although practical, had we grouped all species together (e.g. Leddy et al. 1999) or used a foraging guild like granivores (e.g. Devereux et al. 2008), we would not have detected displacement in Le Conte’s Sparrows at our study site because of the much larger sample size of Savannah Sparrows. Nevertheless, if grouping species is necessary in a displacement study, we recommend grouping based on social behavior and habitat use.
References
Askins RA, Chavez-Ramirez F, Dal BC, Haas CA, Herkert JR, Knope FL, Vickery PD (2007) Conservation of grassland birds in North America: understanding ecological processes in different regions. Ornithol Monogr 64:1–46
Brennan LA, Kuvlesky WP (2005) North American grassland birds: an unfolding conservation crisis? J Wildl Manag 69:1–13
Butler AB, Martin JA, Palmer WE, Carroll JP (2009) Winter use of south Florida dry prairie by two declining grassland passerines. Condor 111:511–522
Compton BW, Rhymer JM, McCollough M (2002) Habitat selection by wood turtles (Clemmys insculpta): an application of paired logistic regression. Ecology 83:833–843
Devereux CL, Denny MJH, Whittingham MJ (2008) Minimal effects of wind turbines on the distribution of wintering farmland birds. J Appl Ecol 45:1689–1694
Diamond DD, Smeins FE (1984) Remnant grassland vegetation and ecological affinities of the upper coastal prairie of Texas. Southwest Nat 29:321–334
Douglas DJT, Bellamy PE, Pearce-Higgins JW (2011) Changes in the abundance and distribution of upland breeding birds at an operational wind farm. Bird Study 58:37–43
Drewitt AL, Langston RHW (2006) Assessing the impacts of wind farms on birds. Ibis 148:29–42
Dunning J (2001) New world Sparrows. In: Elphick C, Dunning J, Sibley D (eds) The Sibley guide to bird life and behavior. Alfred A. Knopf, New York, pp 516–535
Elith J, Graham CH, Anderson RP, Dudík M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton JM, Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberón J, Williams S, Wisz MS, Zimmermann NE (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
Fielding AH, Bell JF (1997) A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ Conserv 24:38–49
Fletcher RJ, Dhundale JA, Dean TF (2000) Estimating non-breeding season bird abundance in prairies: a comparison of two survey techniques. J Field Ornithol 71:321–329
Griffith GE, Bryce SB, Omernik JM, Rogers A (2007) Ecoregions of Texas. Texas Commission on Environmental Quality, Austin
Grzybowski JA (1982) Population structure in grassland bird communities during winter. Condor 84:137–152
Grzybowski JA (1983) Sociality of grassland birds during winter. Behav Ecol Sociobiol 13:211–219
Hernandez PA, Graham CH, Master LL, Albert DL (2006) The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 29:773–785
Hirzel AH, Le Lay GI (2008) Habitat suitability modeling and niche theory. J Appl Ecol 45:1372–1381
Igl LD, Ballard BM (1999) Habitat associations of migrating and overwintering grassland birds in southern Texas. Condor 101:771–782
Keating KA, Cherry S (2004) Use and interpretation of logistic regression in habitat selection studies. J Wildl Manag 68:774–789
Kiesecker JM, Evans JS, Fargione J, Doherty K, Foresman KR, Kunz TH, Naugle D, Nibbelink NP, Niemuth ND (2011) Win-win for wind and wildlife: a vision to facilitate sustainable development. PLoS One 6:e17566
Knight DH (1994) Methods for sampling vegetation: an instruction manual for botany 4700. University of Wyoming, Laramie
Knopf FL (1994) Avian assemblages on altered grasslands. Stud Avian Biol 15:247–257
Kuvlesky WP, Brennan LA, Morrison ML, Boydston KK, Ballard BM, Bryant FC (2007) Wind energy development and wildlife conservation: challenges and opportunities. J Wildl Manag 71:2487–2498
Lammers WM, Collopy MW (2007) Effectiveness of avian predator perch deterrents on electrical transmission lines. J Wildl Manag 71:2752–2758
Leddy KL, Higgins KF, Naugle DE (1999) Effects of wind turbines on upland nesting birds in conservation reserve program grasslands. Wilson Bull 111:100–104
Macias-Duarte A, Panjabi AO (2013) Association of habitat characteristics with winter survival of a declining grassland bird in Chihuahuan Desert grasslands of Mexico. Auk 130:141–149
Macias-Duarte A, Montoya AB, Mendez-Gonzalez CE, Rodriguez-Salazar JR, Hunt WG, Krannitz PG (2009) Factors influencing habitat use by migratory grassland birds in the state of Chihuahua, Mexico. Auk 126:896–905
Marra PP, Hobson KA, Holmes RT (1998) Linking winter and summer events in a migratory bird using stable carbon isotopes. Science 282:1884–1886
Pearce-Higgins JW, Stephen L, Langston RHW, Bainbridge IP, Bullman R (2009) The distribution of breeding birds around upland wind farms. J Appl Ecol 46:1323–1331
Pearce-Higgins JW, Stephen L, Douse A, Langston RHW (2012) Greater impacts of wind farms on bird populations during construction than subsequent operation: results of a multi-site and multi-species analysis. J Appl Ecol 49:386–394
Pearson RG, Thuiller W, Araujo MB, Martinez-Meyer E, Brotons L, McClean C, Miles L, Segurado P, Dawson TP, Lees DC (2006) Model-based uncertainty in species range prediction. J Biogeogr 33:1704–1711
Pearson RG, Raxworthy CJ, Nakamura M, Peterson AT (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117
Peterjohn BG, Sauer JR (1999) Population status of North American grassland birds from the North American breeding bird survey, 1966–1996. Stud Avian Biol 19:27–44
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Pruett CL, Patten MA, Wolfe DH (2009) Avoidance behavior by prairie grouse: implications for development of wind energy. Conserv Biol 23:1253–1259
Pulliam HR, Mills GS (1977) The use of space by wintering sparrows. Ecology 58:1393–1399
Rich TD, Beardmore CJ, Berlanga H, Blancher PJ, Bradstreet MSW, Butcher GS, Demarest DW, Dunn EH, Hunter WC, Iñigo-Elias EE, Kennedy JA, Martell AM, Panjabi AO, Pashley DN, Rosenberg KV, Rustay CM, Wendt JS, Will TC (2004) Partners in flight North American landbird conservation plan. Cornell Lab of Ornithology, Ithaca
Roberts JP, Schnell GD (2006) Comparison of survey methods for wintering grassland birds. J Field Ornithol 77:46–60
Smith MD, Barbour PJ, Burger LW Jr, Dinsmore SJ (2005) Density and diversity of overwintering birds in managed field borders in Mississippi. Wilson Bull 117:258–269
Twedt DJ, Hamel PB, Wodrey MS (2008) Winter bird population studies and project prairie birds for surveying grassland birds. Southeast Nat 7:11–18
Vickery PD, Herkert JR (2001) Recent advances in grassland bird research: where do we go from here? Auk 118:11–15
Wise MS, Hijmans RJ, Li J, Peterson AT, Graham CH, Guisan A, NCEAS Predicting Species Distributions Working Group (2008) Effects of sample size on the performance of species distribution models. Divers Distrib 14:763–773
Acknowledgments
We thank J. Ellis for her assistance in the field and the employees of Wolf Ridge for logistical support. Funding was provided by NextEra Energy Resources; TCU researchers had unrestricted access to company data, and complete independence in all aspects of the analysis, conclusions, and decision to publish the research. T.K.S. received a TCU Biology Department Adkins Fellowship. The manuscript was improved by comments from D. Williams, A. Lehikoinen, T. Burgess, and three anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Stevens, T.K., Hale, A.M., Karsten, K.B. et al. An analysis of displacement from wind turbines in a wintering grassland bird community. Biodivers Conserv 22, 1755–1767 (2013). https://doi.org/10.1007/s10531-013-0510-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-013-0510-8