
Abstract
As part of the expanding cultivation of renewable primary products, oilseed rape has become a prominent part of the agricultural landscape of many European countries. However, the ecological impacts of increasing cultivation of oilseed rape have received little attention so far. In this study, we provide the first systematic investigation of the suitability of oilseed rape as a habitat for the Bluethroat (Luscinia svecica cyanecula), an endangered and highly specialized bird species. We conducted territorial mapping and a habitat analysis, revealing the distinct habitat requirements of L. s. cyanecula in oilseed rape fields over three breeding seasons. The soil type was identified as the crucial factor for habitat occupancy. It acts as a surrogate for soil moisture and this in turn decisively influences food availability. Ditches can further improve habitat quality, whereas reed is not a necessary structure, although it is a regular feature in the primary habitat of the Bluethroat. Occupied oilseed rape stands were structurally different from the semi-natural reed habitat, but provided similar essential habitat requirements, such as shelter from predation and moist, bare soil. Our results clearly demonstrate the utilization of oilseed rape by a rare and threatened wetland bird species and suggest that this crop may act as a secondary habitat.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Renewable primary products and bioenergy production have become increasingly important on the world market during the last decades. In particular, the cultivation of winter oilseed rape (Brassica napus ssp. napus, OSR or canola) has rapidly expanded in many European agricultural landscapes. OSR is now the second most important oilseed crop in the world (USDA Foreign Agriculture Service 2010) and the third most important field crop in Germany. Currently, Germany is the fourth most important producer of OSR in the world (http://faostat.fao.org 2007), and produces over 50% of the EU’s biodiesel, which is 80% derived from OSR (FAO 2008). OSR has therefore become a major element in the agricultural landscape, and further expansion is expected (OECD–FAO 2010).
Oilseed rape is used in the food-, non-food-, and chemical sectors as well as for bioenergy production; cultivation of OSR will continue to be of agricultural and economic importance. For this reason, oilseed rape has been the subject of many studies. These contribute to our knowledge about genetically modified cultivars (e.g. Ceddia et al. 2009; Crawley et al. 1993; Reuter et al. 2009), oil content (e.g. Omidi et al. 2010), improved crop yield (e.g. Sieling and Christen 1997), insect pests (e.g. Veromann 2008) and pollination (e.g. Jauker and Wolters 2008).
However, the ecological impacts of OSR on landscape structure and biodiversity have received relatively little attention so far. Due to the dense and homogeneous structure that excludes certain bird species, and high biocide inputs which potentially have negative effects on birds, oilseed rape is often considered critical by ecologists (e.g. Hoffmann 2007; Bommer 2000a). As with the expansion of maize-growing during recent decades, increasing OSR cultivation will cause a simplification of landscape structure and reduced availability of (semi-) natural habitats, imposing further threats to biodiversity in agricultural landscapes (Stoate et al. 2001). Declining populations of farmland bird species, linked to agricultural intensification, are the subjects of ongoing debate. Although this general trend is recognized (Chamberlain et al. 2000; Donald et al. 2001), some recent studies question this assumption. Bird populations in different regions seem to be affected by different factors (cf. Fox 2004; Wretenberg et al. 2006). For example, species that were increasing in the UK declined in Denmark, despite both countries experiencing the same agricultural intensification (Fox 2004). In contrast, in Sweden, bird species-richness declined although agricultural land use was less intensive than in the UK (Wretenberg et al. 2006). These apparent contradictions emphasize the need for more specific studies to elucidate regional and national patterns.
Despite the general downward trend in farmland bird populations, there is also an increasing number of findings suggesting partly positive effects from current agricultural developments (e.g. Newton 2004). This includes OSR fields, which may provide appropriate habitats for nutrition and reproduction in certain species (Bommer 2000a; Burton et al. 1999; Inglis et al. 1990; Newton 2004), perhaps surprisingly including the white-spotted Bluethroat (Luscina svecica cyanecula). This wetland species, with highly specific habitat requirements, has been considered rare and highly threatened in many European countries until recently (Berndt 1995; De Cornulier et al. 1997; Theiß 1991).
Colonization of arable land by the Bluethroat was first recorded along reed-covered draining ditches in near-coastal areas of the Netherlands and north-western Germany. In these cases, the reed-covered ditches served as the essential habitat structure resembling the primary wetland habitat in eutrophic fens and floodplains, whereas the neighboring arable fields were at best used as supplementary feeding ground (Blaszyk 1963; Reiter 1994). The colonization of oilseed rape fields is a very recent development that started in several Central European regions only in the 1990s (e.g. De Cornulier et al. 1997; Hustings et al. 1995; Kreuziger and Stübing 2005; Theiß 1991). Observations of Bluethroats in oilseed rape in the northern Upper Rhine Valley were made in the late 1990s (Wolf 2000). This rapid colonization of oilseed rape fields was unexpected and there is an ongoing debate over whether this is a temporary phenomenon caused by shifts of populations in typical floodplain habitats during years with severe summer floods (Kreuziger 2001). Similarly, the reasons for the structural suitability of oilseed rape stands as Bluethroat habitats, and the necessity of additional marginal structures such as overgrown ditches, are still under discussion (Krüger 2002).
Here we present data from three different breeding seasons and analysis of the relationship between Bluethroat population and habitat structure of oilseed rape fields. With this study we provide the first systematic investigation of the suitability of oilseed rape fields as habitat for Bluethroats.
Specifically we address the following questions:
-
(i)
Do population densities in oilseed rape fields resemble those in primary wetland habitats?
-
(ii)
What are the main factors affecting the suitability of oilseed rape fields as a Bluethroat habitat?
-
(iii)
Is there structural similarity between oilseed rape stands and primary wetland habitat?
Materials and methods
Study area and study species
The study area is located in an arable region of the northern Upper Rhine Valley in Hesse, Germany (49°51′N, 8°25′E). It comprises 2,610 ha, situated in the Holocene floodplain of the Rhine and the adjacent late Pleistocene lower terrace. As a condition of arable land use, the fossil section of the floodplain is protected against direct flooding by dikes, whereas the functional floodplain is regularly flooded. Nevertheless, the fossil section is still influenced by the riverine water regime and characterized by a high, albeit strongly fluctuating, water table and semi terrestrial soil types. In contrast, the lower terrace has a low water table and drier, purely terrestrial, soil types. The climate of the region is characterized by high mean annual temperatures and low precipitation compared to average Central European conditions (Böger 1991). More than 90% of the study area is under arable agriculture. Typical linear structures comprise field tracks, tree lines, hedgerows and ditches. The latter are confined to the floodplain.
Agriculture is dominated by the intense cultivation of cereals such as wheat, barley and maize. Following the general European trend, oilseed rape growing has rapidly expanded over the past 20 years. Nevertheless in most years it comprises less than 5% of the arable area (Table 1). Winter oilseed rape is sown in autumn and hibernates as a rosette. Flowering starts from mid April onwards and is mainly over by the end of May. The main harvest period in the study area is from about the middle to the end of July (authors’ observations).
Due to early stem elongation in spring, OSR is already tall by the time the first Bluethroats arrive from their northwest African wintering areas in the last ten days of March.
Generally, the arrival of Bluethroats in German breeding areas peaks in the first half of April (Glutz von Blotzheim and Bauer 1988) and finishes at the end of April. Egg-laying starts mid-April (Bonnet 1984; Franz 1998; Krüger 1997; Schmidt 1995). The majority of first broods are fledged by the end of May (Theiß and Franz 1986; Cramp 1988). If habitat conditions are suitable, a second brood can be initiated (Theiß and Franz 1986) which is reared between June and July. Nests are small cups placed on the ground in dense vegetation (Cramp 1988). Preferred breeding habitats are characterized by wet or at least humid conditions with a mosaic of dense vegetation and bare soil, e.g. natural reed beds in riverine systems, bogs or fens (Cramp 1988; Glutz von Blotzheim and Bauer 1988). Food comprises terrestrial invertebrates, mostly insects and their larvae, which is taken from the ground or low vegetation (Cramp 1988). Important predators are ground dwelling mammals, e.g. brown rat (Rattus norvegicus), water vole (Arvicola amphibius), least weasel (Mustela nivalis), stoat (Mustela erminea), European polecat (Mustela putorius) and Red fox (Vulpes vulpes) (Schmidt 1995).
Methods
Mapping of territories
In three years between 2001 and 2007, surveys were carried out. In each survey, breeding male Bluethroats’ territories in the focal oilseed rape fields were mapped with additional use of tape playback (Bibby et al. 1995; Sorjonen and Merila 2000). The surveys were conducted in May and June during the periods of highest Bluethroat singing activity, between 5 a.m. and 11 a.m. and between 5 p.m. and 10 p.m. Territories were only counted as such if males with territorial behavior (song and song-flights) were observed at least twice in the same place. Observations were made every day except when the weather was very windy or rainy. Each field was examined at least three times during each survey year. As far as possible, all sides of each field were accessed during a survey. On average, surveying a field took 15 min, but this varied depending on field size and accessibility. The order, in which the OSR fields in the study area were visited, and the direction of the survey route were chosen at random. In 2002 territory mapping was confined to the floodplain, hence no data for the lower terrace is available.
Habitat measurements
Given Bluethroats’ small breeding ranges (Schlemmer 1988), we mapped all structures adjacent to the focal OSR fields and treated them as linear structures for subsequent analyses. The structures present were mainly ditches, roads, groves and other crops. The crop height (cm), size (ha) and soil type of each oilseed rape field were recorded. Soil type was determined by a soil map (Weidner 1990) and categorized into four classes ranked from dry to wet comprising luvisols (1), fluvisols and vertisols (2), glyosols (3) and fens (4). A detailed list of the examined parameters is given in Table 2.
Habitat measurements were conducted in 2001 and 2007.
Vegetation structure
In 2007 a comparison between the primary habitat (reed, Phragmites australis dominated community) and the secondary habitat (OSR) was carried out. The structure of both habitat types was assessed in terms of vertical density (surface area covered by stalks per square meter) and horizontal density (horizontal transparency at different heights). For both habitat types, four stands with five replicates each (n = 20) were assessed. Stands were chosen randomly in the area of the floodplain. The OSR stands assessed held at least one Bluethroat territory. The reed stands assessed were known to hold breeding Bluethroats consistently (e.g. Handke 1982; Kreuziger 1997), so occupancy was assumed.
To examine vertical density the number of stalks on a defined area was counted and the diameters of each stalk taken. To assess the horizontal structure of the stand, the transparency of the vegetation was assessed as percentage density at ten different heights above ground (10, 20, 30, 40, 50, 70, 90, 110, 130, 150 cm). Heights were marked on a board (160 × 60 cm), which was placed in the stand and percentage cover was assessed to the nearest 5% from a distance of 50 cm. If coverage was less than 5%, scoring was to the nearest 1%; this occurred only in the lowest or highest levels (10, 20 and 150 cm height).
Statistics
The analysis of habitat selection in Bluethroats, based on the environmental structure of each mapped OSR field of the survey years 2001 and 2007, was performed in R 2.13.0 (R Development Core Team 2008), complemented by the packages Gstat (Pebesma 2004) and design (Harrell 2009), using generalized linear models (GLMs). The logistic regression model includes parameters of the environmental structure as predictor variables, and presence/absence of Bluethroats as the response variable (dichotomous).
We used the XY-coordinates of the centroids of the OSR fields as a proxy for their spatial distribution (n = 104) and constructed a bubble plot with ‘Gstat’ (Pebesma 2004). Since spatial autocorrelation was detected when both the lower terrace and the floodplain were considered in the GLM, and taking results of design plot exploration into account, we decided to restrict further analysis to the oilseed rape fields situated in the floodplain (n = 72). Rechecking of the new model revealed no spatial autocorrelation.
We checked for collinearity among the predictor variables using pairplots (Zuur et al. 2007), and a threshold of Spearman’s rank correlation coefficient higher than 0.8 (Menard 2002). As no collinearity was detected, we retained all the variables for analysis. Possible interactions between variables were detected by looking at conditional scatterplots (coplots) and included in the GLM (Zuur et al. 2007). Data exploration by design plot, which allows a direct comparison of the means of nominal variables, indicated that the soil types varied between the floodplain and the lower terrace (Fig. 1) additionally suggesting a possible interaction between the soil type and occupancy.

Number of fields according to their soil types depending on their location in the study area (floodplain vs. lower terrace). For soil type categories see Table 2
In total, 14 predictor variables and 13 interactions entered the regression model (Table 2). To allow for non-linear effects, we considered a model with the linear terms and included squared terms. But as this did not reveal any significant variables we removed the higher order term and proceeded with just the linear terms. We applied a backward stepwise approach for model simplification. Variables were sequentially removed from the model at a P-value of <0.5. A logit link function was used. All models were also assessed using Akaike’s Information Criterion (AIC).
Models were validated by visual examination of the residuals (Zuur et al. 2007).
Vegetation structure in terms of stand density of OSR versus reed stands were tested for differences using Mann–Whitney U tests. With a sample size >30, the Z-approximation test is used and the obtained asymptotic significance P-value (2-tailed) is given. Vegetation structure was analyzed using SPSS 14.0.1.
Results
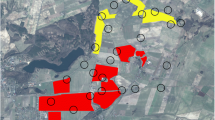
In all surveys we observed a consistently high number of around 49 Bluethroat territories in the oilseed rape fields (Table 1). Maximum numbers of territories per field was 5, with a median of 2 over all years. Field size ranged from 0.5 to 12.8 ha. The smallest field occupied by Bluethroats was 0.5 ha and the biggest 10.5 ha. Mean population density for occupied fields over all years was 6 territories per 10 ha (Table 1). In individual oilseed rape fields, densities ranged from 1.6 to 26.5 territories per 10 ha. Furthermore, total number of territories increased with increasing overall field size during the survey period of 7 years, resulting in a constant population density during all survey years. Notably, in every year, territories were confined to the floodplain of the investigation area, whereas the lower terrace remained completely unoccupied despite containing a similar extent of rape fields (Fig. 2).

Location of Bluethroat territories in the study area in 2001, 2002 and 2007. Oilseed rape fields are shaded
Examination of this pattern revealed distinct differences in the availability of soil types in the study area (Fig. 1). The lower terrace is wholly on soil type 1, whereas the floodplain is dominated by soil type 2. Exploration of relationships between Bluethroat occupancy and the occurance of different soil types by design plot indicated that there were fewer Bluethroat territories on soil type 1 than on the other three soil types.
In the floodplain, 50% of fields with soil type 1, 65% of soil type 2, 83% of soil type 3 and 100% of soil type 4 were occupied by Bluethroats.
The GLM revealed a positive influence of the length of ditches alongside OSR fields on Bluethroat occupancy (Table 3). Thus, the chance of recording a Bluethroat in the OSR field increases with increasing ditch length; a one-unit change in the parameter ‘ditch’ corresponds to a change in the odds of 1.015. This means that the probability of recording a Bluethroat in the considered OSR field changes by 1.5%, if one meter of ditch were added to the habitat. All other variables entered into the regression model were not significant. Figure 3 illustrates the relationship between Bluethroat occurence in an OSR field and the length or availability of ditches.

Bluethroat occupancy of OSR fields (0 absence, 1 presence) in the floodplain depending on the total length of adjacent ditches (m)
The analysis of the vegetation structure revealed significant differences between oilseed rape and reed stands. The two habitat types show opposite trends in vertical and horizontal structure (Fig. 4). The vegetation densities decreased with increasing height in reed stands, whereas they increased in OSR stands. Reed stands were characterized by a thick layer of litter at ground level, giving 100% coverage in the first 5 cm of height above ground. In contrast, OSR stands showed an average cover of only 6% at ground level. Instead, they were densest at the canopy level, 130 cm above ground.

Horizontal densities in oilseed rape (n = 20) and reed stands (n = 20) of the northern Upper Rhine Valley. Mean value ± SD
Similar findings were obtained for stand densities. The Mann–Whitney-U-test revealed significant differences in the amount of surface covered by stalks (Z = −2.518, P < 0.05). One square meter of reed held on the average 400 stems with a mean diameter of 0.5 cm. In contrast, one square meter of oilseed rape comprised a mean of 45 stems each 1.2 cm in diameter. Thus, a mean of about 0.7% of the substrate was occupied by stems in reed stands, and about 0.5% in OSR.
Discussion
Bluethroats are often considered to be habitat specialists confined to natural (reed) and semi-natural (overgrown ditches) wetland habitats. Occurrences of the Bluethroat in agricultural landscapes are mainly associated with reed (Blaszyk 1963; Petersen 2001). Krüger (2002) found that only rapeseed fields with adjacent reed-covered ditches were occupied by Bluethroats in Lower Saxony, Germany. Similarly, reed was significant for permanent occupancy of territories in the Austrian Hanság (Reiter 1994). However, our results indicate that oilseed rape, in itself, is an appropriate habitat for this species. Regression analysis revealed no predictive power of reed and other taller or more natural marginal structures.
Grüll (2001) mentioned reed as a key factor for the settlement of Bluethroats in the poorly-covered agricultural landscape due to its function as good shelter. In our study area, however, it can be assumed that this function has been taken over by oilseed rape stands themselves, in that they provide dense vegetation cover even in early spring due to their dense felted canopy, composed of twigs and pods.
Instead, the soil type turned out to be the most important factor. It accounts for the main difference between lower terrace and floodplain (Fig. 1), of which only the latter harbors Bluethroat territories. However, it is questionable whether it is the soil type itself, that is the important factor, or some other parameter that is closely linked to and represented by the soil type.
Due to the morphological and hydrological situation of our study area, it can be assumed that soil type is closely linked to soil moisture. According to their structural and compositional characterization, soil types markedly differ in their water balance, resulting in a ranking of our soil type categories from 1, the driest to 4, the wettest (Sponagel et al. 2005; Hessisches Landesamt für Umwelt und Geologie 2002). This can be proved during periods of high groundwater levels. During a particularly strong and long-lasting flooding event in spring 2001 soil types in the floodplain were clearly reflected by the height and duration of inundation, whereas the terrace remained completely dry, with groundwater levels 1 m below the soil surface (information derived from maps online available at: http://www.grundwasser-online.de/gwo_portal/index.php). In line with this findings regression analysis revealed drainage ditch length as the only significant variable for occupancy (Table 3; Fig. 3), which may be seen as a surrogate of soil moisture and the average height of the ground water level in a particular area.
Soil moisture most likely has a direct effect on the quantity and quality of food supply. Gilroy et al. (2008) found occurance of Yellow Wagtails (Motacilla flava flavissima) to be related to soil penetrability, and this in turn to influence insect availability. In case of the Bluethroat, findings that soil moisture is linked to food availability have been made by several other studies (e.g. Bommer 2000a; Geslin et al. 2002; Lübcke 1990; Schmidt 1995). Bluethroats mainly forage on insects and their larvae, which may be more abundant in moist conditions. This is indicated by several studies linking insect preferences with soil moisture (e.g. Holland et al. 2007; Langenstück 1997; Traugott 2006).
The dense canopy in OSR fields is responsible for a cool microclimate in the stand due to shading (Alpmann 2006). OSR therefore promotes retention of soil moisture and its associated good foraging conditions (Kruckenberg 1999). This corresponds to the findings of Gruar et al. (2006) for Reed Buntings in Britain, which indicates that OSR fields supply food and nesting resources also related to wetter soil conditions. Reed buntings spent 76% of foraging events in OSR fields, although this crop only accounted for 39% of the available habitats (Burton et al. 1999). Generally, various studies suggest OSR to be attractive to bird species because of a good invertebrate (and seed) supply (e.g. Bommer 2000a; Gruar et al. 2006; Stoate et al. 1998; Surmacki 2005; Watson and Rae 1998; Wolf 2000). In case of the Bluethroat, De Cornulier et al. (1997) showed rape to be rich in insects and assumed this to be the main reason for its settlement. Reiter (1994) found non-permanent occupied territories to be significantly more strongly correlated with OSR than with any other field crop; this is explained by good food availability and sheltered foraging.
During field work, Bluethroats were rarely observed foraging outside OSR fields. This observation is supported by the fact that the shady canopy of OSR prevents understory growth, and thus provides bare soil sparsely covered with vegetation on large scale. At the same time, we could show (Fig. 4) that birds’ mobility on the ground is not hampered, due to the early die-back of leaves low on the stalk. This is necessary for the Bluethroat, which searches for its food on the ground and on lower vegetation (Franz 1998). Thus, it is possible that the Bluethroats studied mainly foraged in the oilseed rape fields since other broad-leaved crops [mentioned as foraging places by Blaszyk (1963) and Petersen (2001)], were largely absent from the study site; it was dominated by short-spired wheat, which is unsuitable for Bluethroats (Blaszyk 1963; Reiter 1994).
Regression analysis indicated an additional influence of nearby ditches on site occupancy by Bluethroats. The importance of wet ditches for several bird species is often emphasized (e.g. Peach et al. 2004), but an actual proof is still lacking. Preliminary work mostly associates the link between the presence of ditches and the occurrence of Bluethroats with foraging opportunities, availability of nesting sites and presence of reed (Blaszyk 1963; Krüger 2002; Petersen 2001; Schlemmer 1988). However, this does not apply to our study site, because most ditches did not contain reed. Usually they were densely covered with grassland vegetation, and even the ditch banks regularly mown. Furthermore, 30% of the occupied rape fields did not have adjacent ditches. Ditches might act as a kind of “hot spot” for insect emergence, as they represent the wettest place in the area—and a defined and stable level of moisture has been shown to be important for invertebrate biomass and emergence densities (e.g. Gilroy et al. 2010; Holland et al. 2007), in accordance with the theorized relationship between soil type and moisture that we have proposed. Morris and Webb (1987) stated that ditches enhance insect diversity by increasing the variety of microhabitats available to arthropod species, and Leon-Cortes et al. (2000) found the highest densities of butterflies along ditches despite their rarity in their study area. Although the ditches of our study site themselves do not appear to be suitable foraging sites for Bluethroats, insects may spread from there into the surrounding habitat, and thus become available for foraging Bluethroats.
Gruar et al. (2006) suggest a potential advantage of nesting in OSR due to the relative inaccessibility of nests to predators, leading to greater breeding success. But, at least for second broods that are reared between mid-June and July (Theiß and Franz 1986; Glutz von Blotzheim and Bauer 1988), the early harvest of OSR by the mid to end of July might lead to a complete loss of this brood, if executed before fledging of young. Therefore, future investigations on breeding success are clearly needed to confirm our findings.
On average OSR is treated once with an herbicide, fungicide and insecticide each from April to flowering (Wolf 2000). After flowering around 10th May until harvest hardly any mechanical stress or chemical applications take place (Bommer 2000b; Cramer 1990; Wolf 2000). The varied impact of biocide applications is indicated by recent research. Whereas Burton et al. (1999) found spraying with a desiccating herbicide prior to harvest not to be harmful for eggs and nestlings of Reed Buntings, Gruar et al. (2006) emphasize the negative effect of molluscicides on wildlife. Further research on the impact of biocides is needed.
Structural similarity to reed is often considered to be the reason for the occupancy of oilseed rape by Bluethroats (e.g. Berndt 1995; Bommer 2000b; Kreuziger 2001). Contrary to such expectations, our analysis of the vegetation structure revealed strong differences between rape and reed. But both habitats provide nesting as well as foraging sites, and protection from predators; only their spatial configuration is different. Whereas reed stands are characterized by densely covered patches with adjoining patches sparsely covered with vegetation, in oilseed rape, these characteristics are present simultaneously and in the same place.
Although our study is not directly comparable with theirs in terms of the study area, we observed similar population densities to those seen by Reiter (1994) for an agricultural area of the Austrian Hanság and by Orlowski and Sek (2005) for semi-natural reed-beds of south-western Poland. In our study, population densities across the whole study area amounted to 0.2 territories per 10 ha. The mean density for occupied OSR fields was 6 territories per 10 ha, corresponding with the maximum densities observed in Central Europe (Bauer et al. 2005). Within individual occupied fields, the maximum densities observed were 26.5 territories per 10 ha. An increasing number of territories and higher probability of occurrence with increasing field size (Baldi and Kisbenedek 1999; Mason and MacDonald 2000; Orlowski and Sek 2005) could not be confirmed in our study. In fact, even very small fields of 0.5 ha were occupied by Bluethroats. These findings support the conclusion that oilseed rape fields are a suitable habitat for the Bluethroat.
Conclusions
Taking account of the microclimate, soil moisture, sparsely covered soil surface and good protection, oilseed rape alone apparently may provide appropriate habitat conditions for Bluethroats. Similar findings have been made for the Reed Bunting in Britain (Gruar et al. 2006), which has also been traditionally considered as a typical wetland species. However, as indicated by their complete absence from the late lower terrace, high ground water levels seem to be necessary for successful long-term colonization of OSR by the Bluethroat.
Some authors (e.g. Kreuziger 2001) assume that Bluethroats only temporarily occupy OSR fields located next to (large) reed beds in times of strong population pressure and if primary habitats are simultaneously destroyed by severe summer floods. Our results, confirming constant colonization of OSR fields over seven years, plus recent findings from other regions (e.g. Krüger 2002), do not support this assumption. For the explanation of the recent population increase of the Bluethroat in Central Europe (e.g. Bauer et al. 2005; Franz 1998), the rapid and unexpected use of OSR fields must be considered a crucial factor. The predicted increasing cultivation of oilseed rape in the EU can therefore be of continuing support for Bluethroat populations. Although an OSR monoculture is not favorable, at least oilseed rape fields on groundwater-proximate alluvial soils can be of conservation importance in view of the continuing loss of natural primary habitats. Therefore further research on this topic, with particular focus on the evaluation of actual breeding success, is strongly needed.
This study shows that against the recent trend of declining farmland bird species, oilseed rape fields have the potential to improve the habitat situation and stabilize population size of certain species.
References
Alpmann L (2006) Raps–Anbau und Verwertung einer Kultur mit Perspektive. Landwirtschaftsverlag, Münster
Baldi A, Kisbenedek T (1999) Species-specific distribution of reed-nesting passerine birds across reed-bed edges: effects of spatial scale and edge type. Acta Zool Hung 45:97–114
Bauer H-G, Bezzel E, Fiedler W (2005) Das Kompendium der Vögel Mitteleuropas: Alles über Biologie. Gefährdung und Schutz, Aula
Berndt RK (1995) Aktuelle Veränderung der Habitatwahl schleswig-holsteinischer Brutvögel–Verstädterung, Wechsel von Nadel- in Laubholz, Besiedlung von Wintersaaten und Ackerbrachen. Corax 16:109–124
Bibby CJ, Burgess ND, Hill DA, Bauer H-G (1995) Methoden der Feldornithologie: Bestandserfassung in der Praxis. Neumann, Radebeul
Blaszyk P (1963) Das Weißsternige Blaukehlchen, Luscinia svecica cyanecula, als Kulturfolger in der gebüschlosen Ackermarsch. J Ornithol 104:168–181
Böger K (1991) Grünlandvegetation im Hessischen Ried: Pflanzensoziologische Verhältnisse und Naturschutzkonzeption. Botanische Vereinigung für Naturschutz in Hessen, Frankfurt am Main
Bommer K (2000a) Ölsaaten als zunehmend bevorzugte Nist- und Nahrungsstätten für wildlebende Vögel und Säugetiere in Oberschwaben/Baden-Württemberg mit Ausblicken auf Mitteleuropa, Biozidanwendungen sowie Brutvögel in Hanf und Leguminosen. Orn Jh Bad-Württ 16:85–176
Bommer K (2000b) Machen Ölsaaten Vögel fett? Falke 47:100–107
Bonnet P (1984) Les Passereaux marqueurs d’anthropisation dans un marais salant de l’ouest de la France (Guérande). Thèse Univ, Rennes I
Burton NHK, Watts PN, Crick HQP et al (1999) The effects of preharvesting operations on reed buntings Emberiza schoeniclus nesting in oilseed rape Brassica napus. Bird Study 46:369–372
Ceddia MG, Bartlett M, Perrings C (2009) Quantifying the effect of buffer zones, crop areas and spatial aggregation on the externalities of genetically modified crops at landscape level. Agric Ecosyst Environ 129:65–72
Chamberlain DE, Fuller RJ, Bunce RGH et al (2000) Changes in the abundance of farmland birds in relation to the timing of agricultural intensification in England and Wales. J Appl Ecol 37:771–788
Cramer N (1990) Raps : Anbau und Verwertung ; [Züchtung - Anbau und Vermarktung von Körnerraps]. Ulmer, Stuttgart
Cramp S (1988) The birds of the western Palearctic, vol V. Oxford University Press, Oxford
Crawley MJ, Hails RS, Rees M et al (1993) Ecology of transgenic oilseed rape in natural habitats. Nature 363:620–623
De Cornulier T, Bernard R, Arroyo B et al (1997) Geographic extension and change in the ecology of the Bluethroat Luscinia svecica in central-western France. Alauda 65:1–6
Donald PF, Green RE, Heath MV (2001) Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc R Soc B 268:25–29
FAO (2008) The state of food and agriculture biofuels: prospects, risks and opportunities. FAO, Rome
Fox A (2004) Has Danish agriculture maintained farmland bird populations? J Appl Ecol 41:427–439
Franz D (1998) Das Blaukehlchen: Von der Rarität zum Allerweltsvogel?. Aula-Verlag, Wiesbaden
Geslin T, Lefeuvre J-C, Le Pajolec Y et al (2002) Salt exploitation and landscape structure in a breeding population of the threatened bluethroat (Luscinia svecica) in salt-pans in western France. Biol Conserv 107:283–289
Gilroy JJ, Anderson GQA, Grice PV et al (2008) Could soil degradation contribute to farmland bird declines? Links between soil penetrability and the abundance of yellow wagtails Motacilla flava in arable fields. Biol Conserv 141(12):3116–3126
Gilroy JJ, Anderson GQA, Grice PV et al (2010) Mid-season shifts in the habitat associations of yellow Wagtails Motacilla flava breeding in arable farmland. Ibis 152(1):90–104
Glutz von Blotzheim UN, Bauer KM (1988) Handbuch der Vögel Mitteleuropas. Aula, Wiesbaden
Gruar D, Barritt D, Peach WJ (2006) Summer utilization of oilseed rape by reed buntings Emberiza schoeniclus and other farmland birds. Bird Study 53:47–54
Grüll A (2001) Populationsuntersuchungen am Weißsternigen Blaukehlchen (Luscinia svecica cyanecula) im Neusiedler See-Gebiet. Egretta 44:1–44
Handke K (1982) Ergebnisse einjähriger Burtvogel-Untersuchungen in Hessens größtem NSG “Kühkopf-Knoblochsaue” (Kreis Groß-Gerau). Luscinia 44:269–302
Harrell FE (2009) Design: Design Package. R package version 2.3-0. http://CRAN.R-project.org/package=Design
Hessisches Landesamt für Umwelt und Geologie (2002) Erläuterung zur Bodenkarte von Hessen 1:50000. http://www.hlug.de/static/medien/boden/fisbo/bk/bfd50/extdoc/pdf/erl_bk_l9999.pdf. Cited 1 July 2011
Hoffmann J (2007) Vogelindikator für die Agrarlandschaft auf der Grundlage der Abundanzen der Brutvogelarten im Kontext zur räumlichen Landschaftsstruktur. Landbauforschung Völkenrode 57(4):333–347
Holland JM, Thomas CFG, Birkett T et al (2007) Spatio-temporal distribution and emergence of beetles in arable fields in relation to soil moisture. Bull Entomol Res 97(01):89–100
Hustings F, Froppen R, Beemster N et al (1995) Spectaculaire opleving van Blauwborst Luscinia svecica cyanecula als broedvogel in Nederland. Limosa 68:147–158
Inglis IR, Isaacson AJ, Thearle RJP et al (1990) The effects of changing agricultural practice upon Woodpigeon Columba palumbus numbers. Ibis 132:262–272
IUSS Working Group WRB (2006) World reference base for soil resources 2006. World Soil Resources Report No. 103. FAO, Rome
Jauker F, Wolters V (2008) Hover flies are efficient pollinators of oilseed rape. Oecologia 156:819–823
Kreuziger J (1997) Die Bedeutung von Sukzessions- und Renaturierungsprozessen für die Vogelgemeinschaft einer Flußaue (NSG Kühlkopf-Knoblochsaue, Nördliche Oberrheinniederung, Hessen). Dissertation, Techn. Hochsch., Darmstadt
Kreuziger J (2001) “Heute hier–morgen dort”: Bestandsentwicklung des Blaukehlchens (Luscinia svecica cyanecula) im NSG “Kühkopf–Knoblochsaue” (Kreis Groß–Gerau). Vogel u Umwelt 12:33–45
Kreuziger J, Stübing S (2005) Die aktuelle Bestandssituation des Blaukehlchens (Luscinia svecica) in Hessen. Vogel u Umwelt 16:31–42
Kruckenberg H (1999) Erfolg der ostfriesischen Nachtigall: Blaukehlchen in Ostfriesland. Falke 46:36–40
Krüger T (1997) Das Blaukehlchen Luscinia svecica im Oldenburger Land. Jahresber Ornithol Arb gem Oldenbg 14:46–69
Krüger T (2002) Verbreitung, Bestand und Habitatwahl des Blaukehlchens (Luscinia svecica cyanecula) in Niedersachsen 2001: Ergebnisse einer landesweiten Erfassung. Vogelkdl Ber Niedersachs 34:1–21
Langenstück C (1997) Beitrag zur Biologie der Canthariden-Larven auf Ackerflächen in SO-Niedersachsen. Diplomarbeit, Technische Universität Braunschweig
Leon-Cortes JL, Cowley MJR, Thomas C (2000) The distribution and decline of a widespread butterfly Lycaena phlaeas in a pastoral landscape. Ecol Entomol 25(3):285–294
Lübcke W (1990) Wie wirkt sich die Zunahme von Mais- und Rapsanbau auf die Vogelwelt aus? Vogelkdl Ber Edertal 16:55–64
Mason CF, MacDonald SM (2000) Corn Bunting Miliaria calandra populations, landscape and land-use in an arable district of eastern England. Bird Conserv Int 10:169–186
Menard SW (2002) Applied logistic regression analysis. SAGE, Thousand Oaks
Morris MG, Webb NR (1987) The importance of field margins for the conservation of insects. Br Crop Prof Counc Monogr 35:53–65
Newton I (2004) The recent declines of farmland bird populations in Britain: an appraisal of causal factors and conservation actions. Ibis 146:579–600
OECD–FAO (2010) Agricultural outlook 2010–2019. Organisation for Economic Co-operation and Development, Paris
Omidi H, Tahmasebi Z, Naghdi Badi HA et al (2010) Fatty acid composition of canola (Brassica napus L.), as affected by agronomical, genotypic and environmental parameters. C R Biol 333:248–254
Orlowski G, Sek M (2005) Semi-natural reedbeds as breeding habitat of bluethroat (Luscinia svecica L.) on sewage farm in Wroclaw city (south-western Poland). Pol J Ecol 53:135–142
Peach WJ, Robinson RA, Murray KA (2004) Demographic and environmental causes of the decline of rural song thrushes Turdus philomelos in lowland Britain. Ibis 146:50–59
Pebesma EJ (2004) Multivariable geostatistics in S: the gstat package. Comput Geosci 30:683–691
Petersen B (2001) Zur Verbreitung, Bestandsentwicklung und Habitatwahl des Weißsternigen Blaukehlchens (Luscinia svecica cyanecula) auf der ostfriesischen Halbinsel. Beitr Vogel-u Insektenwelt Ostfriesl 160:3–52
R Development Core Team (2008) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0. http://www.R-project.org
Reiter A (1994) Breeding population and selection of habitat of the Bluethroat (Luscinia svecica cyanecula) in the Austrian part of Hanság in the years 1988–1990 and suggestions for the protection of this species. Egretta 37:45–59
Reuter H, Schmidt G, Schröder W et al (2009) Regional distribution of genetically modified organisms (GMOs)—Up-scaling the dispersal and persistence potential of herbicide resistant oilseed rape (Brassisca napus). Ecol Ind. doi:10.1016/j.ecolind.2009.03.007
Schlemmer R (1988) Untersuchungen zur Habitatstruktur des Weißsternigen Blaukehlchens Luscinia svecica cyanecula, Wolf 1810, im unteren Isartal. Verh Orn Ges Bayern 24:607–650
Schmidt E (1995) Das Blaukehlchen. Neue Brehm-Bücherei. 426. Westarp Wissenschaften, Magdeburg
Sieling K, Christen O (1997) Effect of preceding crop combination and N fertilization on yield of six oil-seed rape cultivars (Brassica napus L.). Eur J Agron 7:301–306
Sorjonen J, Merila J (2000) Response of male Bluethroats Luscinia svecica to song playback: evidence of territorial function of song and song flights. Ornis Fenn 77(1):43–47
Sponagel H, Grottenthaler W, Hartmann KJ (2005) Bodenkundliche Kartieranleitung. Schweizerbart, Stuttgart
Stoate C, Moreby S, Szczur J (1998) Breeding ecology of farmland Yellowhammers Emberiza citrinella. Bird Study 45:109–121
Stoate C, Boatman ND, Borralho RJ et al (2001) Ecological impacts of arable intensification in Europe. J Environ Manage 63:337–365
Surmacki A (2005) Habitat use by three Acrocephalus warblers in an intensively used farmland area: the influence of breeding patch and its surroundings. J Ornithol 146:160–166
Theiß N (1991) Weißsterniges Blaukehlchen Luscinia svecica cyanecula brütet erstmals in einem Rapsfeld. Orn Anz 30:80–82
Theiß N, Franz D (1986) Nachweis von Zweitbruten beim Weißsternigen Blaukehlchen (Luscinia svecica cyanecula). J Ornithol 127:511–513
Traugott M (2006) Habitat use and activity patterns of larval and adult Cantharis beetles in arable land. Eur J Soil Biol 42(2):82–88
USDA Foreign Agriculture Service (2010) Oilseeds: worldmarkets and trade. Circular Services June 2010, Washington DC
Veromann E (2008) Do cropping system and insecticide use in spring oilseed rape affect the abundance of pollen beetles (Meligethes aeneus Fab.) on the crop? Int J Pest Manage 54:1–4
Watson A, Rae R (1998) Use by birds of rape fields in east Scotland. Brit Birds 91(4):144–145
Weidner E (1990) Bodenkarte der Nördlichen Oberrheinebene 1:50000. Hessisches Landesamt für Bodenforschung, Wiesbaden
Wolf H (2000) Vögel im Rapsfeld–Müssen Naturschützer umdenken? Collurio 18:53–62
Wretenberg J, Lindstrom A, Svensson S et al (2006) Population trends of farmland birds in Sweden and England: similar trends but different patterns of agricultural intensification. J Appl Ecol 43:1110–1120
Zuur AF, Ieono EN, Smith GM (2007) Analysing ecological data: statistics for biology and health. Springer Science Business Media, LLC, New York
Acknowledgments
We are grateful to Valentin Klaus, Till Kleinebecker and Benedikt Gießing for valuable comments on the manuscript. We thank Dorothea Lemke for advice with statistical analysis. Matthias Harnisch provided information on the study area. We thank Phil Prosser for considerable improvement of the English writing.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Berndt, A.M., Hölzel, N. Energy crops as a new bird habitat: utilization of oilseed rape fields by the rare Bluethroat (Luscinia svecica). Biodivers Conserv 21, 527–541 (2012). https://doi.org/10.1007/s10531-011-0199-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-011-0199-5