
Abstract
The assessment of species rarity considers local abundance (scarce or abundant population), habitat affinity (stenoecious or euryecious species), and geographic distribution (stenotopic or eurytopic species). When analyzed together these variables classify species into eight categories, from common species to those having small populations, unique habitats, and restricted geographic distribution (form 7), as proposed by Rabinowitz in 1981. Based on these categories, it is possible to calculate the frequency of the different forms of rarity of the species present in a given site. The Brazilian Atlantic rainforest is considered a hotspot of the world biodiversity harboring many endemic species, which have restricted geographic distribution. Our objective was to identify the forms of rarity of tree species and their proportions in the southern portion of the Brazilian Atlantic rainforest using Rabinowitz’s forms of rarity. All the seven forms of rarity are present in the 846 tree species we analyzed: 46% eurytopic and 54% stenotopic, 73% euryecious and 27% stenoecious, 76% locally abundant and 24% locally scarce species. Eurytopic, euryecious locally abundant species accounted for 41.1%, whereas 58.9% were somehow rare: 4.5% eurytopic, euryecious locally scarce, 0.2% eurytopic, stenoecious locally abundant, 0.1% eurytopic, stenoecious locally scarce, 19.5% stenotopic, euryecious locally abundant, 8.0% stenotopic, euryecious locally scarce, 15.6% stenotopic, stenoecious locally abundant, and 11.0% stenotopic, stenoecious locally scarce. Considering that the most restrictive forms of rarity precedes extinction, the application of Rabinowitz’s system demonstrated that most tree species of the southern Brazilian Atlantic rainforest are threatened due to their restricted geographic distribution, restriction to a single habitat, reduced local abundance, or even to a combination of these variables.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Local abundance, habitat preference, and geographic range vary greatly among species, even among related species (Ricklefs 2002), being the main variables used in assessing rarity. Habitat type, taxonomic precision, and persistence over evolutive and ecological time can be considered additional variables in identifying rare species and the restrictions influencing them (Gaston 1994). In 1981, Deborah Rabinowitz suggested that rarity could take on different forms depending on a combination of three main variables (Ricklefs 2002). Considering local abundance, habitat affinity, and geographic range to be independent variables, Rabinowitz et al. (1986) established rarity categories based on their different combinations. When analyzed together, these three variables allow species to be classified into eight categories, in which the first category comprises common species and the eighth category encompasses the most restrictive form of rarity, namely the local endemics. It is possible therefore to calculate the frequency of the different forms of rarity among the species present in a given study area. In order to perform this type of analysis a databank containing sufficient information about the three independent variables is necessary (Rabinowitz et al. 1986). This way of assessing rarity was originally applied to the plant species-poor system of the British Islands (Rabinowitz et al. 1986), but it has also been used in rich systems both for plants in the Amazon forest (Pitman et al. 1999) and for animals all over the world (Yu and Dobson 2000).
A report published by the “Conservation International” indicated that Brazil has the greatest biodiversity among the 17 nations that detain 70% of the plant and animal species on the planet (Mittermeier et al. 1997). Additionally, not only is the absolute number of species high in Brazil, but also the number of endemic plants, which puts this country in first place in terms of global diversity (Dias 1998). Myers (1990) indicated 18 global “hotspots”, and the Brazilian Atlantic rainforest stands out among them. Myers et al. (2000) expanded the criteria for selecting “hotspots”, but the Brazilian Atlantic rainforest continued deserving a high ranking, in the fourth place.
The Brazilian Atlantic rainforest is also known by other names, e.g. Atlantic Forest sensu stricto (s.s.) and Dense Ombrophilous Atlantic Forest Phyto-Ecological Region (Veloso 1992). It originally extended along the Atlantic Coast of Brazil from near the Cape of São Roque in Rio Grande do Norte State until the municipality of Osório in Rio Grande do Sul State (Andrade-Lima 1966). Presently the forest is devastated over most of its range (Silva and Leitão-Filho 1982; Leitão-Filho 1994). The Dense Ombrophilous Atlantic Forest Phyto-Ecological Region is heterogeneous and composed of many different forest formations (Leitão-Filho 1994) and various associated ecosystems (Scarano 2002) whose species diversity can often be greater than that observed in the Amazon Forest (Silva and Leitão-Filho 1982; Martins 1989; Leitão-Filho 1994).
Martins (1991) observed that 9–39% of the species in any survey in the Atlantic rainforest were represented by just a single individual. Scudeller et al. (2001) reported that the Atlantic rainforest in São Paulo State is characterized by a predominance of species with low constancy and restricted distribution. Endemism is very high in the Atlantic rainforest (Gentry 1992; Thomas et al. 1998). In Rabinowitz’s rarity system, these endemisms are included under forms 6 and 7, in which species have restricted geographic distributions and occur in scarse populations confined to unique habitats (Kruckeberg and Rabinowitz 1985).
When environments are disturbed by human activities the populations of many species are reduced or become extinct. Therefore certain categories of species must be closely monitored and conservation efforts focused on them. These species are especially vulnerable if their area of occurrence is limited, with one or just a few small populations, or if they have special habitat requirements (Primack and Rodrigues 2001)—that is, if the species are rare according to the classification of Rabinowitz et al. (1986). The most restrictive forms of rarity can be seen as precursors to extinction. It is important to determine which forms of rarity are present in a given phyto-ecological region in order to be able to intensify conservation measures and reserve planning (that can help preserve a group of species), as well as to plan actions that will help conserve individual species—mainly those having known economic value (landscaping, chemical, forestry, food source, etc.), or representing key-species in the ecological community.
Our general objective was to identify the forms of rarity and their proportions among arboreal species in the southern portion of the Brazilian Atlantic rainforest using the rarity classification system developed by Rabinowitz. Considering the high indices of regional endemism already noted for this rainforest (Gentry 1992; Thomas et al. 1998) and the known patterns of species with low constancy and restricted distribution (Scudeller et al. 2001), we expected to find most species with forms of rarity related to restricted geographic range (stenotopy). Given the large proportion of species represented by single individuals (Martins 1991), we also expected that most stenotopic species also presented small population wherever they occurred.
Materials and methods
Databank of the tree taxa of the Atlantic rainforest
We used the FITOGEO databank system (in a Microsoft Access 97 format) that was compiled to manage floristic and phytosociological information (Scudeller and Martins 2003). In order to delimit the Dense Ombrophilous Atlantic Forest Phyto-Ecological Region, we used the area defined by the Brazilian Institute of Geography and Statistics (Veloso 1992).
The creation of a databank to analyze the abundance categories of Atlantic rainforest species involved three steps. The first step was to filter and copy all of the phytosociological tables referring to tree communities in the Atlantic rainforest from the databank used by Scudeller et al. (2001). We defined a phytosociological table as a list of species with their respective phytosociological descriptors presented in any publication found during our bibliographic search (Caiafa and Martins 2007). A bibliographic reference could have more than one phytosociological table, as these could refer to different localities, altitudes, or soil types.
The second step of establishing the databank consisted of adding new phytosociological tables concerning the Atlantic rainforest derived from dissertations, thesis, books, and publications in specialized periodicals. We considered only periodicals with an editorial board and ISSN. Only references published until the beginning of 2005 that indicated the numbers of individuals of each species sampled, the total size of the sample, and the size of the smallest individual sampled were included. We eliminated references that dealt with the initial stages of forest regeneration, as well as those that only furnished information about the most abundant species, or did not include trees, or that stipulated a maximum height for inclusion in the sample. Applying these criteria, we compiled an up-to-date databank with a total of 225 phytosociological tables from 113 bibliographic references.
The third step consisted of verifying valid binomials to assure uniform taxonomic identities. This was done by using specialized literature (such as theses, monographs, and revisions) as well as consulting sites such as W3TROPICOS www.mobot.mobot.org/w3t/search and The International Plant Names Index www.ipni.org/index.html.
Criteria used for including a phytosociological table into the refined databank
Phytosociological surveys have been performed using many different designs and methods (plots or point-quarters, different sizes of the smallest individual sampled, and different sample sizes, plot sizes and sampling techniques). Also, the taxonomic treatments may have been performed with differing degrees of rigor. These differences can influence the results of the metanalysis, as shown by Caiafa and Martins (2007). As such, we adopted certain selection criteria to attain a minimum of homogeneity among the surveys: (1) the survey site had to be located within the Dense Ombrophilous Atlantic Forest Phyto-Ecological Region as delimited by Veloso (1992), independently of the phytogeographic classification adopted by the author of the publication; (2) the publication had to specify the growth habit of the sampled plants (in order to allow us to pick up the tree species from the phytosociological table); (3) the publication had to specify the abundance of each species and the total size of the sample; (4) more than 80% of the taxa sampled had to be identified to the species level; (5) the smallest individual sampled had to have a trunk diameter at breast height (DBH) ≥ 4.8 or 5 cm; and (6) the minimum sample size had to be 300 individuals. These criteria are similar to those that have been used by other authors working with metadata in Brazilian phyto-ecological regions (Castro et al. 1999, Scudeller et al. 2001).
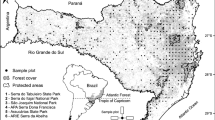
After applying these criteria, we were left with only 41 of the 225 surveys available. The criterion that most contributed to the reduction in the number of acceptable surveys was the size of the smallest individual included in the sample (DBH ≥ 4.8 or 5 cm). This criterion is very important and must not be ignored, as was demonstrated by Caiafa and Martins (2007). Of these 41 surveys, two were from Bahia State and five from Pernambuco, and hence we decided to analyze only the southern region of the Dense Ombrophilous Atlantic Forest—represented by a total of 34 surveys (Fig. 1, Table 1). The southern portion of the Atlantic rainforest includes the states of the Southern Region (Rio Grande do Sul, Santa Catarina, and Paraná) and those of the Southeastern Region (São Paulo, Rio de Janeiro, Minas Gerais, and Espírito Santo).

Position of Brazil in South America (A) and southern Brazil (B).The gray area represents the largest remnants of the Atlantic rainforest.The dots indicate the surveys used in the analysis, and when they are out of the gray area they represent areas bellow the resolution of the map scale. States: MG Minas Gerais, ES Espírito Santo, RJ Rio de Janeiro, SP São Paulo, PR Paraná, SC Santa Catarina e RS Rio Grande do Sul
Classification of the species into forms of rarity
We used the following criteria to classify the species considering the three variables that define the forms of rarity proposed by Rabinowitz et al. (1986):
Geographic distribution
In order to evaluate whether a given species has a wide (eurytopic) or restricted (stenotopic) geographic distribution, the samples were grouped into 2º latitudinal strips (approximately 220 km N–S) starting from the most northern survey (4º S) until the most southern (30º S), for a total of 13 latitudinal belts defining an extension of approximately 2.860 km. We considered species as eurytopic if they were distributed on three or more latitudinal belts (occurring on an extension greater than 660 km). We considered a species as stenotopic if it was found on at most two latitudinal belts (occurring on an extension of at most 440 km). For this classification of geographic ranges we used the entire dataset for the full extension of the Atlantic rainforest (i.e. all 225 phytosociological tables).
Habitat preference
In order to evaluate the habitat preference of the species we considered as distinct habitats the different forest formations that occur in the Dense Ombrophilous Atlantic Forest Phyto-Ecological Region. Veloso (1992) distinguished the vegetation formations in terms of altitudinal belts: (1) Lowlands (LL), with altitudes up to 100 m a.s.l. between 4º N and 16º S, up to 50 m between 16º S and 24º S, and up to 30 m between 24º S and 32º S; (2) Submontane (SB), situated on slopes at altitudes of 100–600 m a.s.l. between 4º N and 16º S, of 50–500 m between 16º S and 24º S, and of 30–400 m between 24º S and 32º S; (3) Montane (M), situated on the top of high plateaus or mountains from 600 to 2000 m a.s.l. between 4º N and 16º S, from 500 to 1500 m between 16º S and 24º S, and from 400 to 1000 m between 24º S and 32º S; and (4) High-montane (HM), situated above the limits established for the Montane formations. The division of the tree flora into altitudinal belts was confirmed by other authors such as Oliveira-Filho and Fontes (2000) and Scudeller et al. (2001). We considered that species would have only a single habitat (stenoecious species) when they occurred in only one of the four forest formations cited above; and we considered species as having various habitats (euryecious species) when they were found to occur in more than one forest formation of the Dense Ombrophilous Atlantic Forest Phyto-Ecological Region. To determine each species habitat preferences we used the complete set of 225 surveys.
Population size
In order to classify the species into abundance categories, we followed Pitman et al. (1999) with modifications according to Martins (1991), who encountered large numbers of species sampled as single individuals in surveys in the Brazilian Atlantic rainforest, and embraced the theory of “singletons” (species represented by just a single individual) and “doubletons” (species represented by only two individuals) proposed by Preston (1962) and developed by Gaston (1994) and other authors. We considered as scarce those species with up to two individuals in any survey analyzed; and as abundant those species sampled with three or more individuals in at least one survey. In this analysis we only considered the refined 34 surveys from the southern portion of the Brazilian Atlantic rainforest that fitted the criteria we established.
After classifying each species according to these three variables, we proceeded to classify the species into the forms of rarity proposed by Rabinowitz et al. (1986). This way we could calculate the proportion of each rarity form in the southern portion of the Dense Ombrophilous Atlantic Forest.
Results
We found all the seven forms of rarity proposed by Rabinowitz et al. (1986) in the southern portion of the Brazilian Atlantic rainforest (Table 2). Of the 846 arboreal species analyzed 46% had wide geographic distribution (eurytopic species) and 54% showed restricted geographic distribution (stenotopic species); 73% occurred in various types of habitats (euryoecious species) and 27% occurred only in unique habitats (stenoecious species); 76% were found in locally abundant populations and 24% were locally scarce. Of the total number of species, 380 (45%) were present only in a single sample.
Of the total 846 species found in the 34 surveys we analyzed, 41.1% were common species (Table 3). These can be considered eurytopic, euryoecious locally abundant species. The other 58.9% species were distributed among rarity forms in the following way. A proportion of 4.5% was rare (form 1) because of their overall small population sizes (Table 4). These species are classified as locally scarce, eurytopic euryoecious species. When a species has a wide geographic distribution with abundant populations, but only occurs in a single habitat, it is rare (form 2) due to its unique habitat, that is, it represents a locally abundant, eurytopic stenoecious species. Only two species (0.2%) had this form of rarity: Andira nitida Mart. ex Benth. and Miconia pusilliflora (DC.) Naud. Species with wide geographic distributions in a single habitat and having scarse populations are rare (form 3) because of the last two characteristics—being locally scarse, eurytopic stenoecious species. Only one species (0.1%) had this form of rarity: Guettarda viburnoides Cham. & Schltdl. A proportion of 19.5% was represented by form 4, that is, locally abundant, stenotopic euryoecious species (Table 5). Locally scarse stenotopic, euryecious species (form 5) represented 8% of the rare species (Table 6). Locally abundant stenotopic, stenoecious species (form 6) accounted for 15.6% of the total (Table 7), whereas 11% were represented by the most restrictive form of rarity (form 7): stenotopic, stenoecious species with locally scarse populations (Table 8).
Discussion
Variables of rarity
Species with restricted geographic distributions and occurring in only a single or at most a few surveys (low constancy) predominate in the Atlantic rainforest of São Paulo State (Scudeller et al. 2001). Accordingly, our results showed that stenotopic species predominate in the southern portion of the Atlantic rainforest. As such, estenotopy is the most important variable in determining the rarity of most of the tree species in the southern region of the Atlantic rainforest. The rarity of tree species due to stenotopy may be the principal variable related to the observed decay in floristic similarity with increasing geographic distance that was reported by Scudeller et al. (2001) in the Atlantic rainforest in São Paulo State. We believe that distribution at distances not greater than 440 km would be a conservative belt width to use even in an extensive phyto-ecological region such as the Dense Ombrophilous Atlantic Forest, especially considering the low latitudinal range found by Scudeller et al. (2001) for tree species in São Paulo State.
Habitat affinity was the second most important variable affecting the forms of rarity of tree species in the southern portion of the Brazilian Atlantic rainforest. In order to classify habitat requirements as either unique (stenoecy) or varied (euryoecy) we considered the occurrence of each species in a single or in more than one altitudinal formations in the Dense Ombrophilous Atlantic Forest Phyto-Ecological Region according to Veloso (1992). In their analysis of the Atlantic rainforest of São Paulo State, Scudeller et al. (2001) detected a strong altitudinal influence on species distribution and abundance, supporting the results of other authors such as Salis et al. (1995) and Torres et al. (1997). As such, we believe that the criteria we adopted to classify the habitat affinity of tree species include important elements of their ecological niche.
Local population size is also an important variable influencing the forms of rarity of tree species in the southern Atlantic rainforest. Our results corroborate other authors, as for example, Scudeller et al. (2001), who concluded that most tree species in the Atlantic Forest s.s. of São Paulo State have restricted ecological niches. Pires and Prance (1977) observed that tree communities of the upland (terra firme) Amazon Forest are composed of few locally abundant species and a plethora of locally scarce species. Our results indicated that the same pattern occurs in the southern portion of the Atlantic rainforest.
Forms of rarity
Our results differed from those reported by Pitman et al. (1999) and Rabinowitz et al. (1986). Pitman et al. (1999) investigated the forms of rarity in little more than 1,500 ha of the lowland rainforest in Peru and analyzed two different matrices: one considering the species occurring in two or more surveys, and another considering the species occurring in three or more surveys. In the study of Rabinowitz et al. (1986) concerning the flora of the British Isles, 15 ecological and taxonomic “judges” classified the species according to rarity variables in order to minimize possible bias in the analyses—which were made by adopting conservative and a liberal criteria. In both studies common species occurred in greater proportions, and some forms of rarity were not found: Pitman et al. (1999) did not find stenotopic species (forms 4–7) and Rabinowitz et al. (1986) did not find some forms of rarity, which varied according to the conservative or liberal criterion used. Differently from Pitman et al. (1999), who examined only a single forest formation, we considered the forms of rarity in the entire southern sector of the Dense Ombrophilous Forest Phyto-Ecological Region, which included different forest formations. Differently from Rabinowitz et al. (1986), who considered all the different plant formations in England, whether forest or not, we considered only rainforest formations. As such, the different criteria used as well as the different study focuses would explain the different results found. One of the most important obstacles to our analysis of rarity was related to problems of taxonomic identification, mainly the large proportion of unknown taxa in the Dense Ombrophilous Atlantic Forest (Caiafa and Martins 2007).
In order to classify the species according to their affinity for the habitat, we adopted the division of the Atlantic rainforest into formations according to Veloso (1992). The heuristics of this procedure was based on various authors, such as Salis et al. (1995), Torres et al. (1997) and Scudeller et al. (2001), who found great consistency in the classification system adopted by the Brazilian Institute of Geography and Statistics IBGE. More studies will be necessary in the Dense Ombrophilous Atlantic Forest Phyto-Ecological Region to investigate variations in species abundance, as the existing surveys do not provide adequade sampling density (Caiafa and Martins 2007). Additionally, the criteria we used to classify the geographic distribution of the species as wide or restricted was subjective. Scudeller et al. (2001) considered species constancy as well as latitudinal range in investigating geographic distribution. In our study, we opted for a subjective division into two-degree latitudinal belts, as it was not possible to use relative constancy. Some species have low relative constancy but a discontinuous distribution, implying that low constancy does not indicate restricted geographic distribution. Oliveira-Filho and Fontes (2000) reported a north–south differentiation in the Atlantic rainforest that was probably caused by variations in temperature and rainfall regimes. We do not know, however, how much of this variation is included within the two-degree latitudinal belts we used to classify the geographic distribution of the species.
Rabinowitz’s approach to classify the forms of rarity involves a certain degree of subjectivity in attributing dichotomic states to each variable of geographic range, habitat preference, and local abundance, whereas these variables are known to vary continuously (Gaston 1994; Pitman et al. 1999). Attributing dichotomic states to continuous variables may imply in classifying some species as common when they are otherwise rare and vice versa. It could be convenient to modify Rabinowitz’s rarity classification system in future studies to consider the effects of continuous variations in those variables. However, in spite of some criticism (Gaston 1994; Pitman et al. 1999), Rabinowitz’s system represents the first solid approach to rarity that includes and systematizes many concepts that otherwise are sparse throughout the literature. We conclude that the application of Rabinowitz’s rarity classification system to the Brazilian Atlantic rainforest confirms stenotopy as the main form of rarity, and also indicates that many tree species are currently threatened due to their restricted geographic distribution, unique habitats, reduced local abundance, or because of a synergism among these variables.
References
Andrade-Lima D (1966) Vegetação. In: Instituto Brasileiro de Geografia e Estatística (ed) Atlas Nacional do Brasil. IBGE, Rio de Janeiro
Caiafa NA, Martins FR (2007) Taxonomic identification, sampling methods, and minimum size of the tree sampled: implications and perspectives for studies in the Brazilian Atlantic Rainforest. FEC 1:95–104
Castro, AAJF, Martins, FR, Tamashiro, JY, Shepherd, J (1999) How rich is the flora of Brazilian Cerrados? Ann Mo Bot Gard 86:192–224
Dias BFS (ed) (1998) Primeiro relatório nacional para a convenção sobre diversidade biológica Brasil. Ministério do Meio Ambiente dos Recursos Hídricos e da Amazônia Legal, Brasília
Gaston K (1994) Rarity. Chapman and Hall, London
Gentry AH (1992) Tropical forest biodiversity: distributional patterns and their conservational significance. Oikos 63:19–82
Kruckeberg AR, Rabinowitz D (1985) Biological aspects of endemism in higher plants. Annu Rev Ecol Syst 16:447–479
Leitão-Filho HF (1994) Diversity of arboreal species in Atlantic rain forest. An Acad Bras Ciênc 66:91–96
Martins FR (1989) Fitossociologia de florestas do Brasil: um histórico bibliográfico. Pesq Série Bot 40:105–161
Martins FR (1991) Estrutura de uma floresta mesófila. Editora da Unicamp, Campinas
Mittermeier, RA, Gil, PR, Mittermeier, CG (1997) Megadiversity: Earth’s biologically wealthiest nations. CEMEX Agrupación Sierra Madre
Myers N (1990) The biodiversity challenge: expanded hot-spots analysis. The Environmentalist 10:243–255
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GAB, Kent J (2000) Biodiversity hot-spots for conservation priorities. Nature 403:853–858
Oliveira-Filho AT, Fontes MAL (2000) Patterns of floristics differentiation among Atlantic forests in south-eastern Brazil, and the influence of climate. Biotropica 32:793–810
Pires JM, Prance GT (1977) The amazon forest: a natural heritage to be preserved. In: Prance GT, Elias TS (eds) Extinction is forever. New York Botanical Garden, New York
Pitman NCA, Terborgh J, Silman MR, Percy-Nuñez V (1999) Tree especies distributions in upper Amazonin forest. Ecology 80:2651–2661
Preston FW (1962) The canonical distribution of commonness and rarity: part I. Ecology 43:185–215
Primack RB, Rodrigues E (2001) Biologia da Conservação. Midiografy, Londrina
Rabinowitz D, Cairns S, Dillon T (1986) Seven forms of rarity and their frequency in the flora of the British Isles. In: Soulé ME (ed) Conservation biology: the science of scarcity and diversity. Sinauer Associates, Massachusetts
Ricklefs R (2002) Rarity and diversity in Amazonian forest trees. Trends Ecol Evol 15:83–84
Salis SM, Tamashiro JY, Shepherd GJ (1995) Floristic comparison of mesophytic semideciduous forests of the interior the State of São Paulo, Southeast of Brazil. Vegetatio 119:155–164
Scarano FR (2002) Structure, function and floristic relationships of plant communities in stressful habitats marginal to the Brazilian Atlantic rain forest. Ann Bot 90:517–524
Scudeller VV, Martins FR (2003) Fitogeo um banco de dados aplicado a fitogeografia. Acta Amaz 33:9–21
Scudeller V, Martins FR, Shepherd GJ (2001) Distribution and abundance of arboreal species in the Atlantic ombrophilous dense forest in Southeastern Brazil. J Plant Ecol 152:185–199
Silva AF, Leitão Filho HF (1982) Composição florística e estrutura de um trecho de mata atlântica de encosta no município de Ubatuba, SP. Rev Bras Bot 9:43–52
Thomas WW, Carvalho AMA, Garrison J, Arbelaez AL (1998) Plant endemism in two forests in southern Bahia, Brazil. Biodivers Conserv 7:311–322
Torres RB, Martins FR, Kinoshita LS (1997) Climate, Soil and tree flora relationships in forests in the state of São Paulo, southeastern Brasil. Rev Bras Bot 20:41–49
Veloso HP (1992) Sistema Fitogeográfico. In: Instituto Brasileiro de Geografia e Estatística (ed) Manual Técnico da Vegetação Brasileira. IBGE, Rio de Janeiro
Yu J, Dobson FS (2000) Seven forms of rarity in mammals. J Biogeography 27:131–139
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Caiafa, A.N., Martins, F.R. Forms of rarity of tree species in the southern Brazilian Atlantic rainforest. Biodivers Conserv 19, 2597–2618 (2010). https://doi.org/10.1007/s10531-010-9861-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-010-9861-6