
Abstract
Oceanic islands have been the grand stage of documented extinctions. In view of limited resources, efficient prioritization is crucial to avoid the extinction of taxa. This work lists the top 100 management priority species for the European archipelagos of the Macaronesian region (Azores, Madeira and the Canary Islands), taking into account both their protection priority and their management feasibility. Bryophytes, vascular plants, molluscs, arthropods and vertebrates were scored by species experts following two sets of criteria: (i) protection priority, including ecological value, singularity, public institutions’ management responsibilities and social value; (ii) management feasibility, including threats knowledge and control feasibility, external socio-economical support for management and biological recovery potential. Environmental managers weighted the same criteria according to their management importance. Final species scores were determined by the combination of both species valuation and criteria weighting. Vascular plants dominate the Top 100 list, followed by arthropods and vertebrates. The majority of listed taxa are endemic to one archipelago or even to a single island. The management feasibility criteria did not dictate that all taxa must be eminently endangered, as for most of the species it should be relatively easy to control threats. The main advantages of this process are the independent participation of scientists and conservation managers, the inclusion of criteria on both protection priority and management feasibility and the taxonomically unbiased nature of the process. This study provides a potentially useful biodiversity conservation tool for the Macaronesian archipelagos that could be readily implemented by the respective regional governments in future legislation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Oceanic islands are unique geological and biological units. In many cases, they had remained isolated from human influence until recently, and they have been the stage for many recent reported extinctions (Lawton and May 1995; Sadler 1999; Brook et al. 2003; Steadman and Martin 2003; Cardoso et al. in press). This has stimulated the debate as to whether endemic island species are inherently less resilient to threats than their continental counterparts (Manne et al. 1999; Sax et al. 2002; Frankham 2005). Though this higher intrinsic vulnerability is often questioned (Manne and Pimm 2001; Biber 2002), islands are relatively small areas with large percentages of endemic species that have little chance of escape from any sudden threat such as habitat destruction or invasive species. Such circumstances may lead to a higher extinction risk (Duncan and Blackburn 2007). In fact, external threats to restricted territories such as oceanic islands have led to more frequent extinctions than in continental territories (Paulay 1994; Quammen 1997).
The emerging conservation crisis in natural ecosystems and processes is part of political agendas in many countries and organizations. Similar to most European countries, Portugal and Spain signed the Convention for Biological Diversity, wherein they agreed to promote species conservation and use the necessary resources to stop the extinction of species by 2010 (UNEP 2002). However, no pragmatic goals were agreed upon, and how each country planned to reach these objectives was not defined (see Baillie et al. 2008). Many different approaches have been devised and proposed for systematic conservation planning (Margules and Pressey 2000; Margules and Sarkar 2007), including a simple ranking of sites according to their species richness or rarity measures (Borges et al. 2005a), measures that take into account the naturalness of habitats (Machado 2004; Cardoso et al. 2007), and the use of algorithms that consider the complementarity of sites, such as irreplaceability value (Pressey et al. 1994; Ferrier et al. 2000). Regardless of the approach used, these measures are ineffective if not accompanied by political actions that legitimise and enforce them (Johnson et al. 2007; Wood et al. 2008).
Legal mechanisms are available in many countries that provide managers with tools and opportunities to counteract extinction processes, including the declaration of a species as threatened (Klemm and Shine, 1993; Martín et al. 2005). However, the concept of a threatened or endangered species is not universal, as the definition of these terms varies. Non-Governmental Organizations (NGOs) have a tendency to apply looser criteria, while governments’ criteria are usually stricter, as they are forced to act if a species is legally declared as threatened (Bouchet et al. 1999; Jeffrey 2001). The concept of threat is itself open to discussion as, for example, a species with a very restricted area of occupancy can be naturally rare but not in decline (Martín 2009).
On the other hand, investment in species with a low probability of avoiding extinction or which require an extreme amount of resources for their recovery might not be the optimal conservation strategy. Because resources are limited, to focus all efforts on a single or a few species will necessarily condemn other species that could be saved with much fewer resources. Regardless of the conservation approach adopted, a combination of scientific and political agendas is needed to best achieve a species priority ranking that is both objective and practical (Bottrill et al. 2008; Schmeller et al. 2008). If such a classification is based upon criteria that have been previously agreed between scientists and managers, then these three requirements for effective conservation can be met: (1) ranking species according to their priority for management in view of limited resources; (2) avoiding ad hoc conservation of species that have priority due only to their charisma; and (3) conversion of a threatened species list into a legally protected species list.
Our work focused on the oceanic archipelagos of the Azores, Madeira (including the Selvagens Islands) and the Canary Islands, which are all part of the Macaronesian biogeographic region. The Macaronesian archipelagos are included in the Mediterranean biodiversity hotspot (Myers et al. 2000). Indeed, the number of unique endemic species and subspecies of terrestrial organisms in these islands is estimated to be around 420 for the Azores (Borges et al. 2005b), 1,419 for Madeira (Borges et al. 2008) and 3,672 for the Canary Islands (Izquierdo et al. 2004).
The aim of this study is to list the 100 taxa with the highest management priority in the Macaronesian region, the so-called Top 100, following an approach adapted from Marsh et al. (2007) based on both ecological and management criteria in oceanic islands. This arbitrary limitation in the number of taxa is a direct path to action, as it identifies species or subspecies that are both threatened and have a high probability of recovery (see Marsh et al. 2007; Bottrill et al. 2008). This list meets the three requirements previously mentioned, as both species experts and environmental managers were involved in its formulation. It includes taxa (species and subspecies) from most major groups of organisms, including arthropods, molluscs, vertebrates, bryophytes and vascular plants. The inclusion of almost all terrestrial (excluding the poorly known fungi and lichens) and some marine taxa is a step forward, one that enables a better understanding of the Macaronesian biodiversity status. The detailed results were published in a book (Martín et al. 2008), which includes a file for each one of the Top 100 Macaronesian species as well as the Top 100 species and results for each archipelago (Arechavaleta and Martín 2008; Cardoso et al. 2008; Faria et al. 2008). These results can readily be adopted by each regional government as important guidelines for future biodiversity conservation legislation.
Materials and methods
The methodology used to list and rank management priority taxa was based on Marsh et al. (2007). Threatened taxa were evaluated for both protection priority and management feasibility, and we obtained the 100 management priority taxa from the combination of both evaluations.
Target taxa selection
It would be impossible to evaluate all the indigenous (native and endemic) species and subspecies present in the study region during this project time-frame. The process of target taxa selection was different for each archipelago. In the Canary Islands, the target taxa were those considered threatened according to the most recent regional evaluations (Martín et al. 2005), as well a few other taxa whose status had recently worsened. In the Azores, due to the lack of such listings, the target taxa were identified by the specialists of each group as under threat and in need of urgent conservation measures, or were included because they were already listed in European directives or lists (Cardoso et al. 2008). In Madeira, the target plant taxa were selected by applying existing threatened species lists (Fontinha et al. 2001; Jardim et al. 2006), while all other taxa were chosen by experts (Faria et al. 2008). The number of target taxa was 310 for the Azores, 190 for Madeira and 156 for the Canary Islands.
Criteria definition
The criteria used to evaluate the taxa were modified from Marsh et al. (2007), in order to better reflect the situation of oceanic islands in general, and the Macaronesian reality in particular. To evaluate a taxon’s protection priority, we considered four factors: (1) ecological species value; (2) species singularity; (3) public institutions’ management responsibilities; and (4) species social value (Table 1). Some of these criteria were further divided into several sub-criteria:
-
PP.1.1. Ecological role—a measure of taxon input into ecological interactions of the ecosystem. Higher scoring was given to species with a unique and irreplaceable role in ecosystem structure or function.
-
PP.2.1. Rarity according to distribution—measured at a 2 × 2 km scale (as in IUCN 2008), this measure was intended to safeguard the taxa with more restricted distributions. Higher scores were given to taxa with smaller areas of occupancy. For the Azores, Canary Islands and molluscs of Madeira, data were obtained from the ATLANTIS database that includes the spatial distribution of all terrestrial species (see details in Izquierdo et al. 2004). For all other taxa in Madeira, data were obtained from specialists’ databases.
-
PP.2.2. Rarity according to abundance—this measure was intended to safeguard taxa that had lower abundances. Higher scoring was given to taxa with lower estimated number of individuals.
-
PP.2.3. Taxonomic singularity—this measure acknowledged that species with a larger phylogenetic distance to their closest relatives have a more unique genetic heritage. Higher scoring was given to species with no close relatives worldwide.
-
PP.3.1. Occurrence—endemic species are probably the most fundamental part of the natural heritage in a small region, and losing them would imply the extinction of a taxon. Taxa that only occur in Macaronesia, or that have a high percentage of individuals in the region, received higher scores.
-
PP.3.2. Population or area of occupancy decline—taxa that are experiencing a significant decline have higher probability of extinction. Higher scores were given to larger losses or any losses in smaller areas.
-
PP.4.1. Social value—this measure indicated the importance society gives to the taxon, either because of direct or indirect benefits. If a species is well regarded by local human populations it had higher scores.
Compared with criteria Marsh et al. (2007) used to measure consequences of extinction, the main difference in this study was the inclusion of species conservation status indicators (criteria PP.2.1, 2.2 and 3.2). These status indicators were added because threatened species lists were not available for many groups in the Azores and Madeira, and hence these criteria were incorporated to increase the scoring of the most threatened taxa. Due to the lack of population genetic structure information for most taxa, this criterion was not used in our work (see Marsh et al. 2007).
To evaluate management feasibility, we considered three factors: (1) knowledge of threats and threat control feasibility; (2) external socio-economical support to species recovery; and (3) species biological recovery potential (Table 2). Some of these criteria were once again further divided into several sub-criteria:
-
MF.1.1. Threat knowledge—to reverse a declining tendency in a given species, it is necessary to know what is causing such a decline. The better the threats were understood, the higher was the score for this criterion.
-
MF.1.2. Threat control feasibility—if it is impossible to control or eliminate a threat, any resources that are applied in this way are simply wasted. Higher threat control success probabilities yielded higher scores.
-
MF.2.1. Financial means and costs—avoiding the extinction of species may be extremely expensive, often requiring specific programs and projects. Taxa that require fewer resources for their conservation were given higher scores.
-
MF.2.2. Community support—if a species is so well regarded that the local human population itself might contribute to its conservation, the management feasibility is much augmented. Higher human population support implied higher scores.
-
MF.2.3. Protected areas—species populations inside protected areas should in theory have higher chances of survival. The higher the percentage of a taxon’s populations that exist inside reserves, the higher the score.
-
MF.3.1. Biological potential—taxa with short biological cycles and high fertility rates are easier to recover, and thus have a higher biological recovery potential. Higher scores were given to taxa with shorter population doubling times.
Compared to the criteria used in Marsh et al. (2007) regarding a taxon’s potential for successful recovery, the main difference here was the inclusion of the protected areas criterion (MF.2.3). This was deemed important since protected areas are probably the most effective conservation tools used in the Macaronesian Islands, and their existence greatly facilitates any recovery action.
Scoring workflow
The (sub-)criteria were first defined by the project coordinators (Martín, Arechavaleta, Borges and Faria), who acted as the liaisons for the three regional public administrations (Fig. 1).

Workflow of the approach followed for this work
The tables defining the criteria (Tables 1 and 2) were then sent to environmental managers from several governmental institutions in the three archipelagos (Fig. 1). Each manager had to distribute a total of 100 points per table according to the different sub-criteria and weight them according to the relative importance of the different species conservation factors. Their scores were then sent back to the project coordinators who averaged all values of all managers. A sample of 24 managers revealed no large dispersion of values, so no further consultations were required (as in the DELPHI methodology, see Linstone and Turoff 1975).
The same tables (Tables 1 and 2) were simultaneously sent to species experts (Fig. 1), who scored each taxon 1–4 for each sub-criterion, according to the parameters previously established, and without previous knowledge of the weighting that environmental managers had attributed to each sub-criterion. Several workshops with the project coordinators and species experts were then conducted in each archipelago to standardise, as much as possible, the interpretation of criteria (Fig. 1). Thirty-six species experts participated in this stage.
Final species scores were calculated by multiplying the raw scores by their respective relative weights, and summing the weighted scores in each criterion. A rank of the 656 target taxa was reached, and the Top 100 management priority taxa were identified.
Results
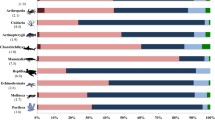
The 100 taxa selected as priorities (listed in Appendix—Table 5) are predominantly from the Canary Islands (Fig. 2). This is in accord with the number of endemic taxa in each archipelago, as the Canary Islands have more than 60% of the Macaronesian endemic species (see above).

Number of species at the Top 100 priority for management at each archipelago and divided by higher taxa
The Top 100 list is largely dominated by flowering plants (Fig. 2). Arthropods and vertebrates follow in the list and all other groups are less represented. All 100 taxa selected are terrestrial and endemic to one of the archipelagos, with the exception of the Mediterranean monk seal (Monachus monachus) and the barnacle (Megabalanus azoricus). Of the 98 endemic taxa, 86 are restricted to a single island and 15 belong to genera endemic to a single archipelago.
The average weight given by environmental managers to each criterion was highly variable (Table 3). Higher importance was given to a species’ ecological role in the ecosystem, threats knowledge and control feasibility. In contrast, species’ social value, community support for species protection and species taxonomic singularity were considered less important (Table 3).
The scoring given by species experts revealed the most common characteristics of the species in the Top 100 priority list (Table 4). Most species share their ecological role with more than two others, and less than one-fifth were considered keystone taxa. These keystone taxa are mainly troglobiont arthropods that often are the only representatives of a particular trophic guild in a cave (such as Trechus oromii and Maiorerus randoi).
The vast majority of taxa have a very restricted distribution, only living on a single island and occupying less than 5% of the island’s area. Some, like the snails Idiomela subplicata and Geomitra turricula or the spurge Euphorbia anachoreta are even restricted to minute islets. Most of these taxa are either plants or arthropods, although three species of Gallotia Canarian lizards and the Azorean bullfinch (Pyrrhula murina) also have extremely restricted distributions. The majority of taxa listed have less than 1,000 mature individuals. Some of the taxa with the most restricted distribution also have very few individuals, with an obvious relationship between distribution and abundance. The Canary plant Solanum vespertilium doramae is among the least abundant, with about ten individuals in the wild, as was Jasminum azoricum in Madeira, with less than six individuals.
The majority of taxa belong to genera with five or more species, and only seven represent monotypic genera. These latter species belong to various groups of organisms, from arthropods (Macarorchiestia martini) and molluscs (I. subplicata) to vertebrates (Neophron percnopterus) and plants (Azorina vidalii, Monizia edulis). Almost all taxa are endemic to a single archipelago, with the exception of the monk seal and the barnacle.
Three-quarters of the taxa suffered some sort of population or distribution decline during the last 10 years or three generations. Many species experienced severe reductions, including 16 plant taxa. The majority of the listed taxa have a low social value, and the exceptions are mostly vertebrates (Fringilla teydea polatzeki) or plants with ornamental or medicinal use (such as Geranium maderense and A. vidalii).
Threats to the survival of all the selected taxa have been identified. Nevertheless, for some species the relative importance of these threats remains unknown, even for vertebrates such as a Canarian bird (F. teydea polatzeki).
The ability to control known threats is usually high. The only species for which known threats seem to be unmanageable is the spurge E. anachoreta, which is endemic to the minute islet of Ilhéu de Fora (Selvagens). In many cases, the costs of threat control are not excessive, and some species require very small investments. For example, the cave spider Turinyphia cavernicola would be better preserved with simple measures directed towards visitors to Algar do Carvão (Terceira, Azores), the cave where the species is endemic. However, threat control for almost half of the taxa would require a long-term financial commitment, likely involving specific projects.
The local human populations are expected to assume a neutral position regarding the management of most taxa. Nevertheless, are there cases where local communities may oppose species management measures. For instance, the pigeon Columba trocaz is hunted for meat in Madeira, and the barnacle M. azoricus is much appreciated as a delicacy in the Azores. On the other hand, three vertebrates of the Canary Islands (N. percnopterus, F. teydea polatzeki and Gallotia simonyi) have received enough community support that recovery actions can be implemented by locals on their own.
Most populations of threatened taxa are found inside protected areas, often created precisely to protect them. However, many cave arthropods are exclusive to caves that are under no protective measures (Cixius cavazoricus, Macharorchiestia martini, M. randoi, Pseudoblothrus oromii, Thalassophilus azoricus, Trechus jorgensis and T. oromii).
It is estimated that most taxa would take between 5 and 10 years to double their populations if all threats were removed. Two taxa, however, have the potential to double their abundance in less than 1 year: the slug Parmacella tenerifensis and the small flowering plant Crambe sventenii.
Discussion
The adaptations made to the criteria in the original proposal of Marsh et al. (2007) allowed us to circumvent two problems in its application: the lack of information for many groups and the idiosyncrasies of oceanic island ecosystems.
We had no information on the population genetic structure for most groups, as equally abundant data are extremely hard to obtain for clearly distinct taxa such as insects and birds. We also did not have threat categorization information for all taxa. Even if this data existed for all groups, it would probably be applied differently for each archipelago, making direct comparisons impossible or at least problematic. Given these two problems, we had to add or delete some criteria from the original proposal (Marsh et al. 2007). The new criteria classified taxa not only according to the consequences of extinction, but also expanded the concept to reflect their protection priority, which includes the species status.
Applying the criteria to oceanic islands also required a number of adaptations. Due to isolation, islands and archipelagos are biogeographic units where the proportion of endemic taxa is much higher than in similar mainland areas. Also, many species have extremely restricted distributions even if they are not endangered (Martín 2009). These characteristics of oceanic islands had to be reflected in the ranking method, and thus new criteria were added. Therefore, our protocol could be considered an adaptation of the Marsh et al. (2007) protocol, but specifically adapted both to a much wider range of taxonomic groups and to island archipelagos.
Among the innovations and advantages of the proposed methodology, the independent participation of both scientists and managers in the definition of species conservation priorities is essential (Fig. 1). While managers are best at recognizing species recovery priorities, they are usually not aware of which species are in greater danger as they are not familiar with the biology and ecology of all the species living in the areas under their jurisdiction. On the contrary, species experts usually know very well the taxonomic status, ecology and biology of the species they study, but are either not aware of what is more important in a recovery program or are too biased towards their taxa of preference to realise the feasibility of their recovery.
Another major advantage of this approach is the taxonomically unbiased nature of the process. It is far too common that species are selected for conservation programs exclusively due to their charisma (Roberge and Angelstam 2004). This leads to disproportionate and unjustified resource allocation towards some taxa. For example, the Birds (79/409/EEC) and Habitats (92/43/EEC) Directives of the European Union are the main drivers of conservation efforts in many European countries, but the protected species lists in these directives are utterly biased towards vertebrates and vascular plants. This implies that only projects concerning species within those groups have access to the European Union LIFE program funding, the largest European contributor to conservation projects. Only with an unbiased approach that covers all taxa for which sufficient information is available will it be possible to better direct resources to species that could benefit the most.
The unbiased taxon selection approach used in this study resulted in group proportions (Fig. 2) that are probably a combination of two factors: the number of species in each group (mainly endemic) and the knowledge available for each species. Vertebrates and vascular plants are arguably the best known, and the Macaronesian islands are rich in endemic vascular plants, hence their dominance on the list. Arthropods, molluscs and bryophytes are the least known groups. Nevertheless, arthropods have many endemic species and represent a relatively large proportion of the 100 priority taxa. An analysis by archipelago reinforces this view, as most of the arthropods listed (12 of the 17 species; Appendix—Table 5) are from the Azores, from where a much more complete knowledge of the spatial distribution and abundance of its endemic arthropod fauna is available (Borges et al. 2005a) than for the other archipelagos.
Some species that ranked very high on the protection priority list did not make the Top 100 list because it would be very hard if not impossible to protect/recover them. This is the case of the beetles Deucalion oceanicum and Paradeucalion desertarum from the Selvagens and Desertas Islands, respectively. Both species were found to be of high conservation interest, but the poor available knowledge would not allow for threat control (Fig. 3). D. oceanicum, for example, is completely dependent upon its host E. anachoreta, another Selvagens endemic species, and the only species in the Top 100 for which the threats are thought to be unmanageable. Despite this difficulty, this Euphorbia species did reach the final list because it was the highest protection priority species of all the initial 656 taxa (Fig. 3, and Appendix—Table 5).

Weighted scores for protection priority and management feasibility for each evaluated taxon. The 100 taxa with the highest combined scores are in black, the taxa that did not reach the Top 100 are in white
An opposite situation to E. anachoreta is that of the frostweed Helianthemum tholiforme from Gran Canaria. This shrub scored average in protection priority; however, its populations are thought to be so easy to manage and recover that it scored the highest of all species in management feasibility, thus reaching the Top 100 list (Fig. 3, Appendix—Table 5). Other plants such as Cheirolophus arboreus and Cheirolophus sventenii gracilis also had very high feasibility scores, but they did not have a high enough protection priority score to be listed.
The taxa with the highest overall scores (see Appendix—Table 5) had to score very high in both fields. This was the case for the water-clover Marsilea azorica from Terceira (Azores) and the frostweed Helianthemum inaguae from Gran Canaria (Fig. 3). Both species live in a single known locality and have very few individuals. However, at least for the latter, artificial propagation has already been attempted with success, causing it to score high in terms of management feasibility (Santana et al. 2006).
Troglobiont organisms, mainly arthropods, were found to be of striking importance. These had the highest ecological role scores, due to the fact that they are usually the single representatives of their trophic guilds in the very simple food chain found in caves. In addition, the distribution of these species is very restricted, with many species endemic to a single cave. However, troglobiont organisms scored the lowest for social value and protected areas, as local human populations are usually unaware of these species, and most caves do not house any flagship vertebrate species and therefore were not included in protected areas. Although some recent work claims to support the flagship species concept (Sergio et al. 2006), this is usually regarded as inefficient or even inappropriate (Cabeza et al. 2008; Roth and Weber 2008). Our study suggests that this concept, which is based on the reasoning that flagship species may serve as umbrella species, clearly fails in many situations.
The relationship between taxon distribution and abundance was apparent in this work. Taxa with a very limited area of occupancy are obviously limited in the number of individuals they can maintain, so they usually scored high in both criteria. This relation has been long known (Gaston 1994; Lawton 2000) and further emphasises the need to protect these “double rare” species. Species that are only able to survive in a single habitat type, which is a common situation for rare species, present all forms of rarity: distribution, abundance and habitat (Rabinowitz 1981).
The taxonomic singularity criterion revealed a pattern common to many oceanic islands, which is that they are isolated from even their closest neighbours, either other archipelagos or the mainland. Oceanic islands harbour many endemic species and genera, and this unique heritage is impossible to safeguard in natural conditions anywhere else.
The Top 100 species are obvious candidates for public awareness initiatives. The total number of species in the priority list, 100, was chosen not according to the species overall scoring, but based on the public appeal of that number. Public outreach should be a main priority of any conservation program and could help raise the social value scores and community support for many of these species in the future (Holmes 2003; Dalton 2005). This can have the negative effect of diverting attention from taxa that were scored immediately below the Top 100 tending to stabilize the list. However, as species are recovered or further endangered, this stabilizing effect can be counteracted.
According to environmental managers, knowledge of the threats and the ability to control them are the two main management feasibility factors, which are even more important than the financial means available. The recovery potential was also highly valued, in recognition that if a species is beyond any reasonable recovery effort it is probably not worthwhile to invest disproportionate resources trying to do so (Restani and Marzluff 2002).
The taxa scores obtained during this evaluation are good candidates for defining future land conservation priorities in Macaronesia. As an example, the “irreplaceability” value of areas to be preserved (Pressey et al. 1994; Ferrier et al. 2000) demands that each species has to reach a specific target, for example, in the proportion of its distribution area that should be protected. These targets can be set according to our overall scores, with high-scoring species demanding higher targets than low-scoring species. Such an approach makes it relatively easy to map irreplaceability for entire regions, and therefore facilitates future delimitation of new protected areas, which in turn helps raise the protected areas criterion scores.
The taxa with high conservation priority scores but for which threats are unknown are obvious candidates for further research. The proposed approach was able to perceive these knowledge gaps, which once filled, will raise the overall scores of those taxa. Likewise, species with high conservation priority scores and low social value are good candidates for awareness programs to improve the knowledge general public has about them. Therefore, the Top 100 species priority list is also a guide for future efforts in ecological research and environmental education.
These results demonstrate that it is possible to include a wide range of taxa when adequate ranking methodologies are applied. The conservation of species requires the joint effort of both the scientific community and conservation managers based in Parks or in the Government. In addition, our results show that for many taxa, conservation action has to go beyond current protected areas, such as with cave species. The Top 100 species priority list should be adopted by the different regional governments as guidelines for future biodiversity conservation legislation. It must be acknowledged, however, that this is a dynamic process and that re-evaluations are desirable every few years. Taxa conservation status, available knowledge, and political and social conditions all change, so the validity of the list presented here will decrease over time. However, this time frame should be long enough so that adequate measures can be implemented without the ghost of the next assessment. Moreover, this work, if continued, represents the starting point of a long-term assessment of Macaronesian biodiversity.
References
Arechavaleta M, Martín JL (2008) La perspectiva archipelágica: Canarias. In: Martín JL, Arechavaleta M, Borges PAV, Faria B (eds) Top 100—Las cien especies amenazadas prioritarias de gestión en la región europea biogeográfica de la Macaronesia. Consejería de Medio Ambiente y Ordenación Territorial, Gobierno de Canarias, Santa Cruz de Tenerife, Spain, pp 129–145
Baillie JEM, Collen B, Amin R et al (2008) Toward monitoring global biodiversity. Conserv Lett 1:18–26
Biber E (2002) Patterns of endemic extinctions among island bird species. Ecography 25:661–676
Borges PAV, Aguiar C, Amaral J et al (2005a) Ranking protected areas in the Azores using standardized sampling of soil epigean arthropods. Biodivers Conserv 14:2029–2060
Borges PAV, Cunha R, Gabriel R et al (2005b) Description of the terrestrial Azorean biodiversity. In: Borges PAV, Cunha R, Gabriel R (eds) A list of the terrestrial fauna (Mollusca and Arthropoda) and flora (Bryophyta, Pteridophyta and Spermatophyta) from the Azores. Direcção Regional de Ambiente and Universidade dos Açores. Horta, Angra do Heroísmo and Ponta Delgada, Portugal, pp 21–68
Borges PAV, Abreu C, Aguiar AMF et al (2008) Description of the Madeira and Selvagens archipelagos terrestrial and freshwater biodiversity. In: Borges PAV, Abreu C, Aguiar AMF (eds) A list of the terrestrial fungi, flora and fauna of Madeira and Selvagens archipelagos. Direcção Regional do Ambiente da Madeira and Universidade dos Açores, Funchal and Angra do Heroísmo, Portugal, pp 13–26
Bottrill MC, Joseph LN, Carwardine J et al (2008) Is conservation triage just smart decision making? Trends Ecol Evol 23:649–654
Bouchet P, Falkner G, Seddon MB (1999) Lists of protected land and freshwater molluscs in the Bern Convention and European Habitats Directive: are they relevant to conservation? Biol Conserv 90:21–31
Brook BW, Sodhi NS, Ng PKL (2003) Catastrophic extinctions follow deforestation in Singapore. Nature 424:420–423
Cabeza M, Arponen A, Van Teeffelen A (2008) Top predators: hot or not? A call for systematic assessment of biodiversity surrogates. J Appl Ecol 45:976–980
Cardoso P, Borges PAV, Gaspar C (2007) Biotic integrity of the arthropod communities in the natural forests of Azores. Biodivers Conserv 16:2883–2901
Cardoso P, Borges PAV, Costa AC et al (2008) La perspectiva archipelágica: Azores. In: Martín JL, Arechavaleta M, Borges PAV, Faria B (eds) Top 100—Las cien especies amenazadas prioritarias de gestión en la región europea biogeográfica de la Macaronesia. Consejería de Medio Ambiente y Ordenación Territorial, Gobierno de Canarias, Santa Cruz de Tenerife, Spain, pp 79–107
Cardoso P, Arnedo MA, Triantis KA, Borges PAV (in press) Drivers of diversity in Macaronesian spiders and the role of species extinctions. J Biogeogr. doi:10.1111/j.1365-2699.2009.02264.x
Dalton TM (2005) Beyond biogeography: a framework for involving the public in planning of U.S. marine protected areas. Conserv Biol 19:1392–1401
Duncan RP, Blackburn TM (2007) Causes of extinction in island birds. Anim Conserv 10:149–150
Faria BF, Abreu C, Aguiar AF (2008) La perspectiva archipelágica: Madeira. In: Martín JL, Arechavaleta M, Borges PAV, Faria B et al (eds) Top 100—Las cien especies amenazadas prioritarias de gestión en la región europea biogeográfica de la Macaronesia. Consejería de Medio Ambiente y Ordenación Territorial, Gobierno de Canarias, Santa Cruz de Tenerife, Spain, pp 109–128
Ferrier S, Pressey RL, Barrett TW (2000) A new predictor of the irreplaceability of areas for achieving a conservation goal, its application to real-world planning, and a research agenda for further refinement. Biol Conserv 93:303–325
Fontinha S, Sim-Sim M, Sérgio C, Hedenäs L (2001) Briófitos Endémicos da Madeira. Biodiversidade Madeirense: Avaliação e Conservação, vol 1. Direcção Regional do Ambiente and Secretaria Regional do Ambiente e dos Recursos Naturais, Funchal, Portugal
Frankham R (2005) Genetics and extinction. Biol Conserv 126:131–140
Gaston KJ (1994) Rarity. Chapman and Hall, London
Holmes CM (2003) The influence of protected area outreach on conservation attitudes and resource use patterns: a case study from western Tanzania. Oryx 37:305–315
IUCN (2008) Guidelines for using the IUCN red list categories and criteria, version 7.0. http://intranet.iucn.org/webfiles/doc/SSC/RedList/RedListGuidelines.pdf. Accessed 13 May 2009
Izquierdo I, Martín JL, Zurita N, Arechavaleta M (eds) (2004) Lista de especies silvestres de Canarias (hongos, plantas y animales terrestres). Consejería de Política Territorial y Medio Ambiente del Gobierno de Canarias, Spain
Jardim R, Fernandes F, Carvalho JA (2006) Flora Vascular. Fauna e Flora da Madeira, Espécies Endémicas Ameaçadas. Vertebrados e Flora Vascular. Governo Regional da Madeira, Funchal, Portugal
Jeffrey DW (2001) The roles of environmental non-governmental organisations in the next century. Biol Environ Proc R Irish Acad 101:151–156
Johnson KN, Duncan S, Spies TA (2007) Regional policy models for forest biodiversity analysis: lessons from coastal Oregon. Ecol Appl 17:81–90
Klemm C, Shine C (1993) Biological diversity conservation and the law: legal mechanisms for conserving species and ecosystems. Environmental Policy and Law Paper 29. IUCN, Gland, Switzerland
Lawton JH (2000) Community ecology in a changing world. International Ecology Institute, Oldendorf/Luhe, Germany
Lawton JH, May RM (1995) Extinction rates. Oxford University Press, Oxford
Linstone HA, Turoff M (eds) (1975) The Delphi method: techniques and applications. Addison-Wesley Publishing Company, Reading, MA
Machado A (2004) An index of naturalness. J Nat Conserv 12:95–110
Manne LL, Pimm SL (2001) Beyond eight forms of rarity: which species are threatened and which will be next? Anim Conserv 4:221–230
Manne LL, Brooks TM, Pimm SL (1999) The relative risk of extinction of passerine birds on continents and islands. Nature 399:258–261
Margules CR, Pressey RL (2000) Systematic conservation planning. Nature 405:243–253
Margules CR, Sarkar S (2007) Systematic conservation planning. Ecology, biodiversity and conservation series. Cambridge University Press, Cambridge
Marsh H, Denis A, Hines H et al (2007) Optimizing allocation of management resources for wildlife. Conserv Biol 21:387–399
Martín JL (2009) Are the IUCN standard home-range thresholds for species a good indicator to prioritize conservation urgency in small islands? A case study in the Canary Islands (Spain). J Nat Conserv 17:87–98
Martín JL, Fajardo S, Cabrera MA et al (2005) Evaluación 2004 de especies amenazadas de Canarias. Especies en peligro de extinción, sensibles a la alteración de su hábitat y vulnerables. Consejería de Medio Ambiente y Ordenación Territorial. Gobierno de Canarias, Santa Cruz de Tenerife, Spain
Martín JL, Arechavaleta M, Borges PAV, Faria BF (eds) (2008) Top 100—Las cien especies amenazadas prioritarias de gestión en la región europea biogeográfica de la Macaronesia. Consejería de Medio Ambiente y Ordenación Territorial. Gobierno de Canarias, Santa Cruz de Tenerife, Spain
Myers N, Mittermeier RA, Mittermeier G et al (2000) Biodiversity hotspots for conservations priorities. Nature 403:853–858
Paulay G (1994) Biodiversity on oceanic islands: its origin and extinction. Am Zool 34:134–144
Pressey RL, Johnson IR, Wilson PD (1994) Shades of irreplaceability: towards a measure of the contribution of sites to a reservation goal. Biodivers Conserv 3:242–262
Quammen D (1997) The song of dodo: island biogeography in an age of extinction. Simon and Schuster, New York
Rabinowitz D (1981) Seven forms of rarity. In: Synge H (ed) The biological aspects of rare plant conservation. Wiley, Chichester, UK, pp 205–217
Restani M, Marzluff JM (2002) Funding extinction? Biological needs in the allocation of resources to endangered species recovery. Bioscience 52:169–177
Roberge JM, Angelstam P (2004) Usefulness of the umbrella species concept as a conservation tool. Conserv Biol 18:76–85
Roth T, Weber D (2008) Top predators as indicators for species richness? Prey species are just as useful. J Appl Ecol 45:987–991
Sadler JP (1999) Biodiversity on oceanic islands: a paleoecological assessment. J Biogeogr 26:75–87
Santana I, Valdés F, Luis JC (2006) Micropropagation of Helianthemum inaguae, a rare and endangered species from the Canary Islands. Bot Macaronesica 26:55–64
Sax DF, Gaines SD, Brown JH (2002) Species invasions exceed extinctions on islands worldwide: a comparative study of plants and birds. Am Nat 160:766–783
Schmeller DS, Gruber B, Budrys E et al (2008) National responsibilities in European species conservation: a methodological review. Conserv Biol 22:593–601
Sergio F, Newton I, Marchesi L, Pedrini P (2006) Ecologically justified charisma: preservation of top predators delivers biodiversity conservation. J Appl Ecol 43:1049–1055
Steadman DW, Martin PS (2003) The late quaternary extinction and future resurrection of birds on Pacific Islands. Earth Sci Rev 61:133–147
UNEP (2002) Report on the sixth meeting of the conference of the parties to the Convention on Biological Diversity (UNEP/CBD/COP/6/20/Part2) Strategic Plan Decision VI/26
Wood LJ, Fish L, Laughren J, Pauly D (2008) Assessing progress towards global marine protection targets shortfalls in information and action. Oryx 42:340–351
Acknowledgements
The generation of the Top 100 list was one of the objectives of the BIONATURA Interreg IIIB project, with the cooperation of ARENA (Azores), Direcção Regional do Ambiente (Madeira) and Dirección General del Medio Natural del Gobierno de Canarias. We are grateful to the following people: Gustavo Vieira, from MBA-consultores, who was the administrative coordinator of the three archipelagos’ associates and responsible for the Interreg secretariat. Sara Santos, who acted as liaison between the researchers of the University of the Azores and Direcção Regional do Ambiente e do Mar (Azores). Elizabeth Ojeda, María Nieves Zurita, Silvia Fajardo and Sofía Rodríguez, who helped in the organization of expert workshops and in finding information for Canary taxa. José Ramón Docoito and Beatriz Herrera, who were in charge of the travel logistics. Salvador de la Cruz, who coordinated the work of experts in the evaluation of target taxa in the Canary Islands. Julián Arechavaleta, who helped with the analyses and software tools. Environmental managers from several public or private institutions from the three archipelagos weighted the priority criteria: António Domingos Abreu, Ángel Bañares, Maria José Bettencourt, Maria Botelho, Miguel Ángel Cabrera, Ana Calero, Bárbara Chaves, José Alberto Delgado, Silvia Fajardo, Ángel Fernández, Manuel Filipe, Paulo Freitas, Mercedes González, María Ángeles Llaría, Nuno Loura, Manuel Martín, Elena Mateo, Félix Medina, João Melo, Dília Menezes, Duarte Nunes, Nuno Pacheco, Paulo Pimentel, Juan Carlos Rando, Pedro Raposo, Miguel Ángel Rodríguez and Rui Sequeira. The evaluators of target taxa, apart from this work’s authors, were: Aurelio Acevedo, Ana Cabrera, Eduardo Carqué, Salvador de la Cruz, Juan Domingo Delgado, José Ramón Docoito, Silvia Fajardo, Eduardo García del Rey, Heriberto López, Manuel Marrero, Katia Martín, José Antonio Mateo, Ricardo Mesa, Helena Morales, José Naranjo, Manuel Naranjo, Elizabeth Ojeda, Alfredo Reyes, María Leticia Rodríguez, Sofía Rodríguez, Rogelio Herrera, Arnoldo Santos and Stephan Scholz. Additionally, Guillermo Delgado and Juan Luis Rodríguez Luengo have participated in workshops for evaluating threatened species.
Author information
Authors and Affiliations
Corresponding author
Appendix
Rights and permissions
About this article
Cite this article
Martín, J.L., Cardoso, P., Arechavaleta, M. et al. Using taxonomically unbiased criteria to prioritize resource allocation for oceanic island species conservation. Biodivers Conserv 19, 1659–1682 (2010). https://doi.org/10.1007/s10531-010-9795-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-010-9795-z