
Abstract
Climate change affects all levels of biology and is a major threat for biodiversity. Hence, it is fundamental to run biodiversity monitoring programs to understand the effects of climate change on the biota and to be able to adjust management and conservation accordingly. So far, however, very few existing monitoring programs allow for the detection of climate change effects, as shown by a survey undertaken by the European project EuMon. Despite this shortcoming, several methods exist which allow to make inferences from existing data by integrating data across different monitoring programs: correlative analyses, meta-analyses and models. In addition, experiments are thought to be useful tools to understand the effects of climate change on plants and animals. Here, we evaluate the utility of these four main approaches. All these methods allow to evaluate long term effects of climate change and make predictions of species’ future development, but they are arguable. We list and compare their benefits and inconveniences, which can lead to uncertainties in the extrapolation of species responses to global climate change. Individual characteristics and population parameters have to be more frequently monitored. The potential evolution of a species should be also modelled, to extrapolate results across spatial and temporal scales as well as to analyse the combined effects of different climatic and biotic factors, including intra but also interspecific relationships. We conclude that a combination of methodologies would be the most promising tool for the assessment of biological responses to climate change, and we provide some thoughts about how to do so. Particularly, we encourage long-term studies along natural gradients (altitudinal or latitudinal) on the same species/habitats to be able to extrapolate to large geographic scales, and to have more complete data sets, necessary to understand the mechanisms of responses. Such data may provide a more accurate base for simulations across spatial and temporal scales, especially if they are publicly available in a common database. These recommendations could allow the adaptation of species management and the development of conservation tools to climate change which threatens species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Current and predicted patterns of global climate change are a major concern in many areas of socio-economy, ecology, farming and politics. In many regions, climate change has resulted in warming over the past century (IPCC 2007). Over the last 30 years, the effects have become particularly obvious in the natural environment (Kerr 2007a, b). Along with disturbance, habitat loss and fragmentation, climate change is one of the main drivers of terrestrial biotic change and has multifarious effects (Fig. 1; Pounds et al. 1999; Walther et al. 2002; Musolin 2007; Skelly et al. 2007), e.g. by increasing the prevalence of pathogens (Bosch et al. 2007), changing behavior, and altering seasonal activities (Saether et al. 2000; Wuethrich 2000). Responses include temperature-dependent physiological responses (direct effects with consequences on biometric, behavioral and phenological traits), responses depending on environmental parameters related to temperature (indirect effects through humidity, precipitation, snow-cover, growth season, extent of sea ice), or from alterations of feedbacks in interspecific exchanges and relationships, such as competition, predation, symbiosis, parasitism and mutualistic association (Davis et al. 1998; Koh et al. 2004; Ahola et al. 2007; Bosch et al. 2007). Climate change affects all levels of life, from the individual, population, species, community and ecosystem to the eco-region level (Fig. 1). Phenological changes are the most frequently demonstrated responses through precocity of migration arrival and/or egg laying/parturition (Winkler et al. 2002) and breeding time shift (Beebee 2002). Another general pattern observed in many species is a poleward or an altitudinal expansion of species’ distributions (Parmesan 2007). If species profit from climate change by expanding their distribution (taking account of problems from invasive species), the ultimate threat to all species will be extinction (Thomas et al. 2004). Estimates of changes in biodiversity that are accurate enough to detect climate change effects are imperative to diagnose the state and trend of biodiversity, and to make adjustments to population management and conservation.

A schematic view of the overall framework on the chain of climate change effects considered for a given species. Direct effects are reported with continuous arrows. Indirect and feedback effects are reported with dotted arrows
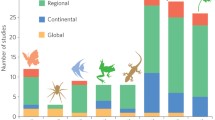
However, it is generally difficult to predict long-term biological responses, as we have little knowledge concerning lag-times between a given effect and its related responses. To show and understand climate change impacts on biodiversity, it is essential to monitor individuals/populations/species over a long time period, usually spanning several decades as effects are detectable only after many years (e.g. Yoccoz et al. 2001; Walther et al. 2002). Usually, the collected data are used in large-scale correlative analyses (e.g. at community or meta-population scales) and the results are combined in meta-analyses. The latter analysis type integrates a large collection of analyses from individual studies (Glass 1976). Further, models are used to simulate the evolution of complex processes. The rarest assessments of biological responses to climate change are experimental approaches. These approaches may be classed in two groups. On the one hand are large scale studies, which give general responses without considering ecological complexity (correlative analysis, meta-analysis and theoretical models). On the other hand are small scale studies that have more explanatory power than large scale studies but show too short time specific responses (population level). The quality of current studies (Fig. 2) shows the need to have multispecific and multiscale responses to improve our current knowledge of climate change effects on biodiversity.

Classification of the studies according to key characteristics such as the number of studied parameters, the spatial/time scale considered or their explanatory power. Explanatory power is the quality, the accuracy and the ability to explain biological phenomenon. The signs + and − mean the value of the quantity of numbers of studied parameters, the quality of the explonatory power and the importance of the scale (time, spatial). Studies at population and individual levels are considered as small scales (we need less time and space to understand functioning of individuals and populations than ecosystems). Studies at metapopulation, species, community, ecosystem and also population levels are considered as large scales because their functioning is complex. For example, in studies at species level, few parameters are measured, explanatory power is quite few, and spatiotemporal scale is often high
Here, we give an overview of biodiversity monitoring related to climate change based on a literature survey. We describe the strengths and weaknesses of different approaches in regard to the power to make predictions of a species’ future fate, and to determinate appropriate time/spatial scales of studies. Finally, we provide recommendations to find the optimal methods for accurately assessing biological responses to climate change.
Materials and methods
We made a literature survey by using the database of ISI web of knowledge and the search string (“climate change” AND response AND “type of analysis”). The string for “type of analysis” corresponds to correlative analyses, meta-analyses, experimental analyses (experimental design, microcosms, open-garden), and models (bioclimatic envelope models, ecological niche models, mechanistic models like population viability analyses). We analyzed a minimum of 20 articles per approach to have a representative sample of each method, and included only those articles which showed effects of global change on biodiversity. To document the best tools for the assessment of recent and future biological responses to climate change, considering current approaches, we only used articles published between 2000 and 2008. We analyzed several parameters important for the analysis of global change impact on biodiversity across studies classified according to their approach (e.g. environmental parameters, time and spatial scales; see Table 2). To measure importance of each approach, we calculated a proportion as the amount of articles studying responses to climate change at a particular ecological level, spatial or temporal scale relative to the total number of articles per approach (given as N for each column in Table 2).
Correlative approaches
Numerous volunteer and professional naturalists in different governmental and non-governmental organizations monitor changes in biodiversity across the globe (Danielsen et al. 2005; Schmeller et al. 2009). Such data are essential for analyses of biological responses to climate change.
Correlative analyses are the most frequently employed approaches to analyze effects of climate change. They consider relationships between an environmental factor(s) and evolution of a character monitored over relatively long time-scales, with a minimum of 10 years in most cases (Chamaillé-Jammes et al. 2006). Correlations are evaluated with respect to an environmental temperature increase [e.g. local annual/monthly/daily mean temperatures, North Atlantic Oscillation (NAO), El Niño Southern Oscillation (ENSO)] or environmental parameters associated with global change (e.g. sea ice cover, CO2 production rate, moisture, Barbraud and Weimerskirch 2001; Walther et al. 2002). The database from EuMon is representative to monitored species and habitats which were used to analyze climate change effects (Table 1). Correlative analyses are applied in all species groups although impact of climate change is less-well studied in mammals (Table 1), and at large organizational scales, such as community, metapopulation or species scales (Hassall et al. 2007). They have been undertaken in various habitats although studies in freshwater habitats were the most represented (Table 1). These show global responses at a regional scale and allow a global and widespread vista of climate change effects on biodiversity. Correlative analyses are based on monitoring of simple parameters like individual counts, presence/absence data, date of egg-laying or estimates of density (Whitfield et al. 2007) at any given scale, but mainly focus on phenological phenomena (Both 2007) and range distribution data (Lovejoy and Hannah 2005).
Studying phenological changes using correlative approaches is a good way to understand the decoupling of phenological relationships, which may alter food-web structure in ecosystems (Both 2007). These are also useful ways to estimate the evolution of interactions among trophic levels. Similarly, correlative analyses show that distribution change responses in individual species may disrupt their interactions with other species at the same or adjacent trophic levels. More recently, correlative studies have investigated other important changes in life history (growth, survival, fecundity, size, Figuerola 2007; Halpern and Cottenie 2007) or genetic traits (Balanyá et al. 2006). Hence, correlative studies are most comprehensive and can estimate species’ adaptive responses that may be related to climate change and underscore important consequences for biodiversity and ecosystem functioning (Wilson et al. 2007). Thus, if correlative analyses are a useful descriptive instrument and have proven to be a relatively efficient method to estimate recent climate change effects, they also have some weaknesses and pose specific statistical problems especially with respect to model selection (Grosbois et al. 2008).
Correlative analyses depend on the quality of monitoring which faces two practical difficulties in regard to a robust assessment of climate change effects on biodiversity. Firstly, there is a need to sustain monitoring across the years to ensure representativeness of time series. Secondly, sufficiently precise monitoring data have to be obtained to allow for the detection of significant changes. Not all monitoring objectives, targeted biological parameters and protocols produce data relevant for the study of global change effects. The need for specific monitoring programs may conflict with the usually limited amount of available financial and human resources (Schmeller et al. 2009) and may lead to inaccuracies in detecting climate change affects (i.e. Yoccoz et al. 2001).
Monitoring schemes which analyze climate change effects are rare. The EuMon project (Schmeller et al. 2006) surveyed biodiversity monitoring efforts across Europe and collected meta-data on 449 species biodiversity monitoring schemes and 69 habitat monitoring schemes (see e.g. Henry et al. 2008; Lengyel et al. 2008). So far, the EuMon database contains 10 habitat monitoring schemes (1.6%) and 48 species schemes (7.7%) begun before 1975. Schemes set up to analyze climate change averaged a duration of 17 ± 2.2 years (median = 11 years). Hence, the EuMon survey (although not being representative of all European countries, see Schmeller et al. 2006) strongly suggests that at the European scale, we have insufficient knowledge on populations at the beginning of the rapid warming period from the mid-1970s, which substantially weakens the power of correlative analyses. Further, monitoring data per se are problematic for adequate analysis of species responses to climate change due to data biases and a lack of statistical power (e.g. Strayer 1999; Parmesan and Yohe 2003; Harker and Shreeve 2008), which need to be considered and corrected (e.g. Van Swaay et al. 2002). Logistic constraints of long-term data collection also result in a low quantity of used parameters in correlations (mean = 4, range = 2–8; Table 2). Thus, species interactions (predation, symbioses, competition) and environmental parameters (except temperatures) are hardly considered. Furthermore, habitats may evolve with respect to temperatures leading to a gradual change interacting with detected biological responses, underestimating or altering biological predictions under climate change.
In addition to limitations on the data, results from correlative analyses must be carefully interpreted (Grosbois et al. 2008). Significant correlation of two parameters indicates a relationship between them, but it does not signify a cause-effect linkage. For example, two environmental conditions, temperature and humidity, are often considered to be interdependent. However, an effect of temperature in a given situation could in reality reflect the impact of humidity (Lorenzon et al. 2001). Such indirect effects have to be considered to avoid misinterpretation and false predictions. To disentangle the effects of each predictor, specific statistical tools can test for the robustness of the correlation. For example, hierarchical partitioning allows separation of the contribution of each predictor in relation to the explained variance in a regression model, both independently and in conjunction with other predictors, and this can be calculated for all possible candidate models (Mac Nally 2002; Grosbois et al. 2008). For such an approach, it is crucial to have data from many possible predictors. With this kind of data, bayesian model averaging (BMA) is a useful technique designed to help account for the uncertainty inherent in the model selection process, a possibility that traditional statistical analysis often neglect. By averaging over many different competing models, BMA incorporates model uncertainty into the results on parameters and predictions. BMA has shown its efficiency for many statistical model classes and has proven that in all cases it improved the performance of the prediction.
Correlative analyses also often assume a linear relationship between cause and effect, neglecting the fact that ecological interactions or individual and population responses to the environment usually show non-linearity (Logan and Allen 1992; Saether et al. 2000). Use of simple linear regression analyses can lead to find wrong responses or no relation between climatic and biologic variables. Thus, predictions from correlative studies must be developed with caution and one must minimize potential traps by including a large number of different environmental factors.
Modelling approaches
Modeling approaches are used to make predictions about the future of biodiversity. Most of the models use an important number of parameters (mean = 6, range = 2–19; Table 2), which means that they can simulate complex processes, allowing for comprehensive predictions. For example, in Tews et al. (2007), the models included species movement, annual primary productivity and several scenarios of climate change. However, most models on global change effects do not incorporate the population level (Griebeler and Seitz 2007). This is especially true so when models are dealing with predictions at the level of the community or the ecosystem (Table 2). Two main types of models are used to predict responses to climate change: mechanistic models and ecological niche models. Mechanistic models are mostly employed to predict changes at the local scale. They are usually used to test relationships between a given species’ species trait and some environmental variable(s) (Delbart et al. 2008). The strategy of such models is to compile relevant information about entities at the lower level of the system (i.e. individuals), to formulate theories about their behaviour, to incorporate these hypothetical behaviours into a simulation model, and to observe the emergence of properties at the system level related to some particular questions (Grimm and Railsback 2005). Few of these models are concerned by making predictions on the evolution of life history traits (Jiguet et al. 2007; Ricker et al. 2007) or about genetic variation (Ditto and Frey 2007). Population viability analyses (PVA; e.g. Henle et al. 2004a; Massot et al. 2008) are used to forecast population health, extinction risk, and to create action plans for conservation (e.g. Henle et al. 2004a, b). In general, a PVA is developed for a target population or species (e.g. a Limestone endemic shrub, Maschinski et al. 2006). The accuracy of a PVA depends on the inclusion of spatial variation, true temporal variation and on the assumptions made about parameters such as survival, migration and reproduction parameters. However, relatively few of the PVA produced so far can predict biodiversity changes under different climate change scenarios (e.g. metapopulation viability of a butterfly under global warming scenarios, Schtickzelle and Baguette 2004). Besides PVA, individual-based simulations are also used to predict the evolution of species dynamics under various climatic constraints (Meynecke 2004; Tews et al. 2007).
Effects of global warming at large spatial scales are usually investigated with bioclimatic envelope models (BEM; Araújo et al. 2005, 2006; Svenning and Skov 2006; Levinsky et al. 2007) or ecological niche models (ENM; Nunes et al. 2007). ENMs are used to predict the geographic range of a species from occurrence (presence/absence) records and environmental data layers (sets of conditions which integrated climatic variables, abiotic/biotic parameters and biotic interactions). They need knowledge of many data. BEM is a forecasting method which considers species or groups by habitat and characterizes their spatial distribution on the basis of environmental parameters (humidity, temperature, etc.). Thus, BEMs can predict the development of the spatial distribution of a species in the context of a future climate change (Hill et al. 2002; Burns et al. 2003; Araújo et al. 2006; Jetz et al. 2007; Levinsky et al. 2007). BEMs are mostly used to adjust the management of species, to improve protection plans (Normand et al. 2007) or to examine the conservation status of species. A three-step process is developed: (1) mathematical models are created to link species to the present climate envelope, (2) climate change scenarios are generated to simulate potential range of the distribution of the species in the future, and (3) comparisons between present and predicted distribution are performed (Moen et al. 2004; Virkkala et al. 2008). The result is an interpolation among known sampling locations on the basis of observations of associations between the characteristics of species and of its environment. An advantage of BEMs, like other models, is the integration of other predictors than temperature (e.g. precipitation, physical characteristics, habitats’ quality and species interactions).
However, BEMs have some shortcomings. Spatial and temporal autocorrelations and lack of accuracy in the data used (Schwartz et al. 2006; Levinsky et al. 2007) can generate significant biases. For example, data are often collected from floristic or faunistic atlases, which merge data collected over a long period of time, ignoring local or recent changes across the whole distribution. BEMs also ignore or make important simplifying assumptions about some biological processes. In particular, they do not take into account the capacity of dispersal of the species and more precisely how the species will modify its dispersal probability according to a change of the environment (Massot et al. 2008). This is also true for the species’ habitat which is hypothesized to strictly follow the climate (no delay, no evolution). Finally, very few climatic scenarios are usually considered (e.g. two scenarios in Normand et al. 2007). Examining a high number of climatic scenarios is essential in order to build predictive envelops such as to increase the probability to encapsulate the true species distribution range evolution. Decisions of conservation and management of species should be made by taking into account the whole set of possibilities, hence the importance to examine a large number of climatic scenarios. To overcome some of these pitfalls, ecologists have started to improve the precision of the predictors included in their models (e.g. Malcolm et al. 2005; Svenning and Skov 2006), to use long-term monitoring data (Andalo et al. 2005; Mouritsen et al. 2005) or to employ methods to validate the predictive performances of their simulations (Araújo et al. 2005).
Models are essential to predict changes in biodiversity and to reinforce policy in biodiversity management (Bakkenes et al. 2002). Models allow describing complex processes at any spatial scale. Simulations, however, are generally global and are relying on a set of assumptions, often not verbalized, which increase even more the uncertainty of projected effects of climate change.
Experimental approaches
Mechanisms by which the climate change is affecting population and community dynamics are best described by using experiments (Etterson 2004; Hollister et al. 2005; Table 2). In particular, experiments permit to explore the effect of even higher temperature increases. Experimental tools are therefore imperative to forecast species responses or to adapt species management and conservation policy (Vilchis et al. 2005). Many species-specific parameters like abundance, phenology (Sherry et al. 2007), fitness (Santos 2007), effects on structure (Roy et al. 2004), interspecific interactions and dynamics of plant communities (Vilchis et al. 2005; Aerts et al. 2006) can be analysed with an experimental approach. Describing experimentally the processes operating at local scales might then allow extrapolation at larger spatial scales (Bruelheide 2003). However, experimental studies of the landscape level or at even larger scale are rather rare mainly because of logistical difficulties (Parmesan 2006).
Two types of experimental designs are developed: microcosm (or common garden) and open garden experiments (e.g. OTCs, Klein et al. 2004; Zavaleta 2006). Open garden experiments are using (semi)-natural areas (Stenstrom and Jonsdottir 2004), while experiments in microcosms are conducted in fully artificial environments (Núñez-Olivera et al. 2004; Santos 2007). In the two cases, the logistics of an experimental protocol may be difficult to implement (because of expensive experimental design and biases involved by use of an artificial environment; Table 2). Experiments may monitor many environmental parameters (temperatures, moisture, nutrients, CO2 quantity, habitat quality; mean = 5, range = 3–12; Table 2). Experiments are usually conducted on a short time scale of 2–3 years and are sufficient to understand primary responses of plants (Hollister et al. 2005; Cross and Harte 2007), invertebrates (Vilchis et al. 2005) and some soil processes to experimental warming (Parton et al. 2007). Such fast responses may help to anticipate the susceptibility of species and habitats responses to future climate changes.
The control of particular environmental parameters is one of the main advantages of an experimental approach. Biotic and abiotic parameters and their interactions can be included in the protocol allowing more accurate predictions for a species’ future (Zavaleta 2006; for an example see Vilchis et al. 2005 or Sherry et al. 2007) and detecting non linearities in a species response. Thus, more recent studies have begun to include a larger number of factors in their experimental design.
Whatever the method (microcosm, OTC, transects), researchers can not control every environmental parameter potentially affected by a temperature increase, i.e. it is impossible to create an artificial habitat reflecting in totality a natural environment. Hence, only certain aspects of climate change can be addressed with experiments and most studies concentrate on the effects of a few environmental variables (temperature, moisture) on particular population/community and on short time scales (Table 2). The complexity arising over larger scales (spatial, temporal) or multitrophic levels are ignored. Experiments may then yield different responses than those observed in nature. For example, Sjögersten and Wookey (2004) showed that an experimental temperature increase leads to a slower decomposition of litter in contradiction to findings by Shaw and Harte (2001) who found that higher temperature correlated with a speedier decomposition.
As we already stated above, experiments are often conduced over a short period of time and focus on animal and plant species with rather short generation times (Table 2). Therefore, generalisation to long-lived species or to long-term dynamics of ecosystems is questionable (e.g. Hollister et al. 2005; Wiedermann et al. 2007). For example, Mora et al. (2007) conclude from their studies that all threats need to be simultaneously considered in an analysis, since the risk of extinction through a combined effect of threats is more important than commonly anticipated. Thus, an experimental approach is a necessary step toward the understanding of complex ecological processes, but researchers have to develop multifactorial experiments at higher trophic levels (birds, squamates and mammals) and on appropriate time and spatial scales to avoid misinterpretation of experimental results.
Meta-analyses
Meta-analysis represents a set of statistical methods for quantitative research synthesis developed in medicine and social sciences in the late 1970s and introduced to ecology at the beginning of the 1990s (Gurevitch et al. 2001). Meta-analyses of climate change effects study a set of biological responses across all/many species groups and often across different spatial scales. Databases used in meta-analyses come from studies applying one or several of the above approaches: correlative analyses, models or experiments. The main parameters studied are abundance, phenology, range distribution, and species diversity (Table 2).
One advantage of meta-analyses is that a broad spectrum of findings can be combined to find a fingerprint of climate change (Root et al. 2003), allowing to infer the species status and to make projections about future biologic changes. Multi-species analyses at large scale, one form of meta-analyses, allow evaluating the responsiveness of many species to environmental perturbations and they can thus derive some common denominator. For example, Jiguet et al. (2007) tested the responses of 71 French breeding birds’ species to an increase of temperature and showed that the thermal range is a relevant predictor of the resilience of a species to extreme temperatures.
Meta-analyses are highly sensitive and may detect even small effects. They are therefore appropriate tools to find general prediction on a species’ future. For example, Briones et al. (2007) reviewed data from 44 studies on enchytraeids (a group of annelids) to predict the impact of climate change on their geographical distribution and were able to predict their future role as source of carbon in response to climate change. Hence, meta-analyses can statistically draw more general and quantitative conclusions on some controversial issues compared to single studies or reviews. They provide new insights and research directions. For instance, Parmesan (2007) showed an average spring advancement in the northern hemisphere of 2.8 days decade−1 but she also spotlighted that the advancement was related to species groups, supporting that amphibians had an earlier breeding than any other taxonomic group and that in butterflies emergence and migratory arrival were earlier than the first flowering of herbs. Those results (see also Barbraud and Weimerskirch 2006) highlight the importance of meta-analyses in understanding and anticipating future consequences of climate change.
However, meta-analysis also has several drawbacks, mainly resulting from publication bias, subjectivity in literature selection and non-independence among studies (Table 2). Indeed, studies showing statistically significant effects are more likely published than those that show no difference (e.g. Palmer 1999; Gurevitch et al. 2001), and thus may underestimate species non-responsiveness to climate change (Parmesan 2007). Publication bias affects the main conclusions of 15–21% of meta-analyses (Jennions and Møller 2002). To detect biases, various methods and software packages (Lei et al. 2007) were developed (like Funnel plot or “trim and fill” methods; Jennions and Møller 2002), allowing correction for potential publication bias. In meta-analyses, few parameters are analysed (mean = 3, range = 2–4; Table 2) mainly because of the difficulties to find variables studied in the same way (with a common definition or a common methodology), crucial point to make comparisons between studies. Indeed, the numerous studies entering a meta-analysis have not the same objectives, parameters, and methodology, which preclude to perform an exhaustive analysis of their results. Consequently, it limits the number of common parameters that can be included in a meta-analysis.
Despite the biases, meta-analyses are a rather robust method to estimate global climate change effects on biodiversity. Interpretation of results needs to consider potential biases, to avoid inaccurate conclusions of future changes and threats of biodiversity.
Discussion
Each of the approaches reviewed here faces methodological, spatial and temporal limitations (Table 2) which constrain their predictive power. Here, we discuss the predictive power of each method and develop a methodological framework for future assessments of biological responses to global climate change.
Correlative analyses are an essential tool to estimate recent effects and species adaptive/maladaptative responses to climate change. They are mostly used to shed new light on changes in phenology and area distributions. However, accuracy of species responses may be biased and the predictive power therefore low. First, correlative analyses depend on the quality of monitoring data. Generally, long time series species data are lacking, as monitoring is costly and labor intensive (Table 2) and very few monitoring schemes are maintained over long periods of time (see EuMon database 2008). Hence, we only have an episodic view of historical populations. Thus, to precisely estimate biological responses to climate change, we need to have either long-term series data or several short-term series in different time periods, but in the same region (for data integration see Henry et al. 2008) at the individual, population, and community levels. In addition, the targeted biological parameters, monitoring objectives and methods are not always relevant. To control for confounding effects or correlation between various causal factors, which could lead to inappropriate biological inferences, we must use statistical tools to test for the robustness of correlations. More biotic and abiotic parameters need also to be available for the region of interest. In spite of the limitation of correlative analyses, they remain an essential approach to analyzing information concerning recent long-term in natura effects of climate change on biodiversity.
Models simulate effects of climate change on species to evaluate long-term effects of climate change and to make predictions on the future fate of species. Model predictions can be validated using recent data collections (e.g. in population viability analyses). The models’ main advantage is their ability to simulate varied and complex biological processes because many biotic and abiotic parameters can be included. However, some uncertainties remain with respect to the reliability of model projections of climate change effects. To increase the predictive power of models, we have to develop appropriate models and apply them at the population level. We also need to build more comprehensive and global models such as mechanistic niche models, i.e. integrating ENM and mechanistic parameters, which integrate species interactions and consider biodiversity at a community level. The improvement of predictions can only be achieved by increasing the range of species responses and the numbers of climatic scenarios. Further, it is imperative to consistently validate the predictive performance of models used for simulation.
Experiments measure rapid species responses in a particular environment at both the population and community levels (Table 2). They allow researchers to set up a more detailed scenario of climate change effects. However, experimental methods have to be designed and chosen carefully. Indeed, microcosm experiments have the advantage of controlling all parameters included in the artificial environment, which is impossible in open-garden experiments. Limitations of experiments include their focus on species with short lifespans (invertebrates, plants) to obtain fast species responses, and on the restricted scale of experimental set-ups. Investigation of complex parameter implications over larger scales or multitrophic levels is very difficult and any extrapolation to long-lived species at larger scales may also be limited. A multiplication of experiments in (semi)-natural environments and with stronger emphasis on monitoring during experiments might, however, help to overcome these limitations.
Meta-analyses represent a highly sensitive method that can allow general predictions on a species’ future and have a more detailed overview with respect to species groups when compared with correlative analyses. Their predictive power to determine species responses may be restricted by the small number of common parameters in different studies. That results from the difficulty in standardizing methods and the period of data collection across studies. Further, a large proportion of studies included in meta-analyses is correlative and therefore represent only a few biological and environmental parameters. Hence, the interpretation of results from meta-analyses needs to be treated with caution and may not be generalizable per se.
Recommendations
A combination of the different methodologies is the most promising approach for assessment of biological responses to climate change. As a first step, the number of observation programs at different spatial scales needs to be increased. Specifically, individual and population parameters need to be more frequently monitored, including life history traits, physiology, genetics, and abundance (Macmynowski et al. 2007). Within this framework, long-term studies offer the best chance for understanding the processes whereby temperature increases will affect a species at a particular scale. Other monitoring programs in similar habitats or on similar species might then complement such studies. These data need to be analyzed across species with different life histories to determine the underlying mechanisms which involve changes in phenology and range distributions at different temporal and spatial scales. Further, data collected in the early decades of the twentieth century need to be made publicly available and integrated in novel analyses. The EuMon project showed that while there is a lot of biodiversity monitoring currently underway in Europe, data integration across schemes is poor. Good integration would greatly improve our understanding of climate change effects on biodiversity (Henry et al. 2008).
In a second step, to improve quality of long-term predictions and better anticipate species management and conservation, we suggest that the most practical approach consists in using natural gradients (latitudinal, altitudinal) on short-lived species, as well as long-lived species. These gradients need to be monitored over an extended time frame, to test many environmental and species’ parameters, to analyse non-linearity of climate change effects and to determine the most important factors for the change. To decouple ecological from evolutionary time scales, it is necessary to consider both the generation time and the number of years. Indeed, if natural selection effects can be detected at very small time scales, the overall response to selection will be better understood at the scale of the generation. This is especially true for species with a long generation time where the ecological and evolutionary time scales are most disconnected. However, both time scales are essential since species management decisions can not be postponed until the effect of global warming is perceptible at an evolutionary scale. Experiments need to focus on a subset of representative species, such that a complete set of approaches and accurate predictions about the effect of climate change are available for these species, allowing for some generalization to other species with similar life history traits. However, generalizations have to be considered with extreme caution, as they depend on the level of organisation studied (individual, population, species, community, ecosystem) and on the environmental parameters: each parameter will vary in time and space at its own scale.
Finally, the future potential development of a species should be modelled, to extrapolate results across spatial and temporal scales and to analyse the combined effects of different climatic and biotic factors. These should include both inter- and intra-specific relationships and both ENM and mechanistic parameters. Besides data integration as a basic step, meta-analyses and other data integrative methods (Henry et al. 2008) can serve a similar purpose, to consider which parameters are important and should be included in experimental and model approaches.
A last point to consider is the ecotoxicological research that could be a reference to adequately study species’ responses to climate change. Similar methods [long-term monitoring (Aebischer 1990); experiments (Bradley and Ormerod 2002); models (Ashauer et al. 2007)] and similar levels of integration (individual, population, communities, species) are used to assess chemical and biological effects in ecological context. From the beginning (1970), ecotoxicological research faced same problems and uncertainties quoted herein. Thus, we should improve methods and analyses considering mistakes, misinterpretations and solutions from ecotoxicological research.
References
Aebischer NJ (1990) Assessing pesticide effects on non-target invertebrates using long-term monitoring and time-series modeling. Funct Ecol 4:369–373. doi:10.2307/2389598
Aerts R, Cornelissen JHC, Dorrepaal E (2006) Plant performance in a warmer world: general responses of plants from cold, northern biomes and the importance of winter and spring events. Plant Ecol 182:65–77
Ahola MP, Laaksonen T, Eeva T et al (2007) Climate change can alter competitive relationships between resident and migratory birds. J Anim Ecol 76:1045–1052. doi:10.1111/j.1365-2656.2007.01294.x
Andalo C, Beaulieu J, Bousquet J (2005) The impact of climate change on growth of local white spruce populations in Québec, Canada. For Ecol Manage 205:169–182. doi:10.1016/j.foreco.2004.10.045
Araújo MB, Pearson RG, Thuillers W et al (2005) Validation of species–climate impact models under climate change. Glob Chang Biol 11:1504–1513. doi:10.1111/j.1365-2486.2005.01000.x
Araújo MB, Thuiller W, Pearson RG (2006) Climate warming and the decline of amphibians and reptiles in Europe. J Biogeogr 33:1712–1728. doi:10.1111/j.1365-2699.2006.01482.x
Ashauer R, Boxall ABA, Brown CD (2007) Simulating toxicity of carbaryl to Gammarus pulex after sequential pulsed exposure. Environ Sci Technol 41:5528–5534. doi:10.1021/es062977v
Bakkenes M, Alkemade JRM, Ihle F et al (2002) Assessing effects of forecasted climate change on the diversity and distribution of European higher plants for 2050. Glob Chang Biol 8:390–407. doi:10.1046/j.1354-1013.2001.00467.x
Balanyá J, Oller JM, Huey RB et al (2006) Global genetic change tracks global climate warming in Drosophila subobscura. Science 313:1773–1775. doi:10.1126/science.1131002
Barbraud C, Weimerskirch H (2001) Emperor penguins and climate change. Nature 411:183–186. doi:10.1038/35075554
Barbraud C, Weimerskirch H (2006) Antarctic birds breed later in response to climate change. Proc Natl Acad Sci USA 103:6248–6251. doi:10.1073/pnas.0510397103
Beebee TJC (2002) Amphibian phenology and climate change. Conserv Biol 16:1454–1455. doi:10.1046/j.1523-1739.2002.02102.x
Bosch J, Carrascal LM, Duran L et al (2007) Climate change and outbreaks of amphibian chytridiomycosis in a montane area of central Spain; is there a link? Proc R Soc B Biol Sci 274:253–260
Both C (2007) Comment on “rapid advance of spring arrival dates in long-distance migratory birds”. Science 315:598b. doi:10.1126/science.1136148
Bradley DC, Ormerod SJ (2002) Long-term effects of catchment liming on invertebrates in upland streams. Freshw Biol 46:161–171. doi:10.1046/j.1365-2427.2002.00770.x
Briones MJI, Ineson P, Heinemeyer A (2007) Predicting potential impacts of climate change on the geographical distribution of enchytraeids: a meta-analysis approach. Glob Chang Biol 13:2252–2269. doi:10.1111/j.1365-2486.2007.01434.x
Bruelheide H (2003) Translocation of a montane meadow to simulate the potential impact of climate change. Appl Veg Sci 6:23–34
Burns CE, Johnston KM, Schmitz OJ (2003) Global climate change and mammalian species diversity in US national parks. Proc Natl Acad Sci USA 100:11474–11477. doi:10.1073/pnas.1635115100
Chamaillé-Jammes S, Massot M, Aragon P et al (2006) Global warming and positive fitness response in mountain populations of common lizards Lacerta vivipara. Glob Chang Biol 12:392–402. doi:10.1111/j.1365-2486.2005.01088.x
Cross MS, Harte J (2007) Compensatory responses to loss of warming-sensitive plant species. Ecology 88:740–748. doi:10.1890/06-1029
Danielsen F, Burgess ND, Balmford A (2005) Monitoring matters: examining the potential of locally-based approaches. Biodivers Conserv 14:2507–2542. doi:10.1007/s10531-005-8375-0
Davis AJ, Jenkinson LS, Lawton JH et al (1998) Making mistakes when predicting shifts in species range in response to global warming. Nature 391:783–786. doi:10.1038/35842
Delbart N, Picard G, Le Toans T et al (2008) Spring phenology in boreal Eurasia over a nearly century time scale. Glob Chang Biol 14:603–614. doi:10.1111/j.1365-2486.2007.01505.x
Ditto AM, Frey JK (2007) Effects of ecogeographic variables on genetic variation in montane mammals: implications for conservation in a global warming scenario. J Biogeogr 34:1136–1149. doi:10.1111/j.1365-2699.2007.01700.x
Etterson JR (2004) Evolutionary potential of Chamaecrista fasciculata in relation to climate change. I. Clinal patterns of selection along an environmental gradient in the great plains. Evol Int J Org Evol 58:1446–1458
EuMon (2008) A comprehensive database on monitoring practices in Europe. http://eumon.ckff.si. Cited 12 Nov 2008
Figuerola J (2007) Climate and dispersal: black-winged stilts disperse further in dry springs. PLoS One 2:e539. doi:10.1371/journal.pone.0000539
Forcada J, Trathan PN, Reid K et al (2006) Contrasting population changes in sympatric penguin species in association with climate warming. Glob Chang Biol 12:411–423. doi:10.1111/j.1365-2486.2006.01108.x
Gilman SE, Wethey DS, Helmuth B (2006) Variation in the sensitivity of organismal body temperature to climate change over local and geographic scales. Proc Natl Acad Sci USA 103:9560–9565. doi:10.1073/pnas.0510992103
Glass GV (1976) Primary, secondary, and meta-analysis of research. Educ Res 5:3–8
Griebeler EM, Seitz A (2007) Effects of increasing temperatures on population dynamics of the zebra mussel Dreissena polymorpha: implications from an individual-based model. Oecologia 151:530–543. doi:10.1007/s00442-006-0591-0
Grimm V, Railsback SF (2005) Individual-based modeling and ecology. Princeton University Press, Princeton, p 428
Grosbois V, Gimenez O, Gaillard J-M, Pradel R, Barbraud C, Clobert J, Møller AP, Weimerskirch H (2008) Assessing the impact of climate change variation on survival in populations. Biol Rev Camb Philos Soc 83:357–399. doi:10.1111/j.1469-185X.2008.00047.x
Gurevitch J, Curtis P, Jones M (2001) Meta-analysis in ecology. Adv Ecol Res 32:199–247. doi:10.1016/S0065-2504(01)32013-5
Halpern BS, Cottenie K (2007) Little evidence for climate effects on local-scale structure and dynamics of California kelp forest communities. Glob Chang Biol 13:236–251. doi:10.1111/j.1365-2486.2006.01243.x
Harker R, Shreeve T (2008) How accurate are single site transect data for monitoring butterfly trends? Spatial and temporal issues identified in monitoring Lasiommata megera. J Insect Conserv 12:125–133. doi:10.1007/s10841-007-9068-7
Hassall C, Thompson DJ, French GC et al (2007) Historical changes in the phenology of British Odonata are related to climate. Glob Chang Biol 13:933–941. doi:10.1111/j.1365-2486.2007.01318.x
Henle K, Davies KF, Kleyer M et al (2004a) Predictors of species sensitivity to fragmentation. Biodivers Conserv 13:207–251. doi:10.1023/B:BIOC.0000004319.91643.9e
Henle K, Sarre S, Wiegand K (2004b) The role of density regulation in extinction processes and population viability analysis. Biodivers Conserv 13:9–52. doi:10.1023/B:BIOC.0000004312.41575.83
Henry P-Y, Lengyel S, Nowicki P et al (2008) Integrating ongoing biodiversity monitoring: potential benefits and methods. Biodivers Conserv 17:3357–3382. doi:10.1007/s10531-008-9417-1
Hill JK, Thomas CD, Fox R et al (2002) Responses of butterflies to twentieth century climate warming: implications for future ranges. Proc R Soc Lond B Biol Sci 269:2163–2171
Hollister RD, Webber PJ, Tweedie C (2005) The response of Alaskan arctic tundra to experimental warming: differences between short- and long-term responses. Glob Chang Biol 11:525–536. doi:10.1111/j.1365-2486.2005.00926.x
Holzapfel AM, Vinebrooke RD (2005) Environmental warming increases invasion potential of alpine lake communities by imported species. Glob Chang Biol 11:2009–2015
IPCC (2007) Summary for policymakers. In: Solomon S, Qin D, Manning M, Chen M, Marquis M, Averyt KB, Tignor M, Miller HL (eds) Climate change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Jennions MD, Møller AP (2002) Publication bias in ecology and evolution: an empirical assessment using the ‘trim and fill’ method. Biol Rev Camb Philos Soc 77:211–222. doi:10.1017/S1464793101005875
Jetz W, Wilcove DS, Dobson AP (2007) Projected impacts of climate and land-use change on the global diversity of birds. PLoS Biol 5:e157. doi:10.1371/journal.pbio.0050157
Jiguet F, Gadot AS, Julliard R et al (2007) Climate envelope, life history traits and the resilience of birds facing global change. Glob Chang Biol 13:1672–1684. doi:10.1111/j.1365-2486.2007.01386.x
Kerr RA (2007a) Global warming: how urgent is climate change? Science 318:1230–1231. doi:10.1126/science.318.5854.1230
Kerr RA (2007b) Climate change: global warming is changing the world. Science 316:188–190. doi:10.1126/science.316.5822.188
Klanderud K, Totland O (2007) The relative role of dispersal and local interactions for alpine plant community diversity under simulated climate warming. Oikos 116:1279–1288. doi:10.1111/j.0030-1299.2007.15906.x
Klein JA, Harte J, Zhao X-Q (2004) Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau. Ecol Lett 7:1170–1179. doi:10.1111/j.1461-0248.2004.00677.x
Koh LP, Dunn RR, Sodhi NS et al (2004) Species coextinctions and the biodiversity crisis. Science 305:1632–1634. doi:10.1126/science.1101101
Lei X, Peng C, Tian D et al (2007) Meta-analysis and its application in global change research. Chin Sci Bull 52:289–302. doi:10.1007/s11434-007-0046-y
Lengyel S, Kobler A, Kutnar L et al (2008) A review and a framework for the integration of biodiversity monitoring at the habitat level. Biodivers Conserv 17:3341–3356. doi:10.1007/s10531-008-9359-7
Levinsky I, Skov F, Svenning J-C et al (2007) Potential impacts of climate change on the distributions and diversity patterns of European mammals. Biodivers Conserv 16:3803–3816. doi:10.1007/s10531-007-9181-7
Logan JA, Allen JC (1992) Nonlinear dynamics and chaos in insect populations. Annu Rev Entomol 37:455–477. doi:10.1146/annurev.en.37.010192.002323
Lorenzon P, Clobert J, Massot M (2001) The contribution of phenotypic plasticity to adaptation in Lacerta vivipara. Evol Int J Org Evol 55:392–404
Lovejoy TE, Hannah L (eds) (2005) Climate change and biodiversity. Yale University Press, New Haven, pp 387–396
Mac Nally R (2002) Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables. Biodivers Conserv 11:1397–1401. doi:10.1023/A:1016250716679
Macmynowski DP, Root TL, Ballard G et al (2007) Changes in spring arrival of nearctic-neotropical migrants attributed to multiscalar climate. Glob Chang Biol 13:2239–2251. doi:10.1111/j.1365-2486.2007.01448.x
Malcolm JR, Markham A, Neilson RP et al (2005) Case study: migration of vegetation types in a greenhouse world. In: Lovejoy TE, Hannah L (eds) Climate change and biodiversity. Yale University Press, New Haven, pp 252–255
Maschinski J, Baggs JE, Quintana-Ascencio PF et al (2006) Using population viability analysis to predict the effects of climate change on the extinction risk of an endangered limestone endemic shrub, arizona cliffrose. Conserv Biol 20:218–228. doi:10.1111/j.1523-1739.2006.00272.x
Massol F, David P, Gerdeaux D et al (2007) The influence of trophic status and large-scale climatic change on the structure of fish communities in Perialpine lakes. J Anim Ecol 76:538–551. doi:10.1111/j.1365-2656.2007.01226.x
Massot M, Clobert J, Ferrière R (2008) Climate warming, dispersal inhibition and extinction risk. Glob Chang Biol 14:461–469. doi:10.1111/j.1365-2486.2007.01514.x
Menzel A, Sparks TH, Estrella N et al (2006) European phenological response to climate change matches the warming pattern. Glob Chang Biol 12:1969–1976. doi:10.1111/j.1365-2486.2006.01193.x
Meynecke J-O (2004) Effects of global climate change on geographic distributions of vertebrates in North Queensland. Ecol Modell 174:347–357. doi:10.1016/j.ecolmodel.2003.07.012
Moen J, Aune K, Edenius L et al (2004) Potential effects of climate change on treeline position in the Swedish mountains. Ecol Soc 9:16
Møller AP, Flensted-Jensen E, Mardal W (2006) Dispersal and climate change: a case study of the Arctic tern Sterna paradisaea. Glob Chang Biol 12:2005–2013. doi:10.1111/j.1365-2486.2006.01216.x
Mora C, Metzger R, Rollo A et al (2007) Experimental simulations about the effects of overexploitation and habitat fragmentation on populations facing environmental warming. Proc R Soc B Biol Sci 274:1023–1028. doi:10.1098/rspb.2006.0338
Mouritsen KN, Tompkins DM, Poulin R (2005) Climate warming may cause a parasite-induced collapse in coastal amphipod populations. Oecologia 146:476–483. doi:10.1007/s00442-005-0223-0
Musolin DL (2007) Insects in a warmer world: ecological, physiological and life-history responses of true bugs (Heteroptera) to climate change. Glob Chang Biol 13:1565–1585. doi:10.1111/j.1365-2486.2007.01395.x
Normand S, Svenning J-C, Skov F (2007) National and European perspectives on climate change sensitivity of the habitats directive characteristic plant species. J Nat Conserv 15:41–53. doi:10.1016/j.jnc.2006.09.001
Nunes MFC, Galetti M, Marsden S et al (2007) Are large-scale distributional shifts of the blue-winged macaw (Primolius maracana) related to climate change? J Biogeogr 34:816–827. doi:10.1111/j.1365-2699.2006.01663.x
Núñez-Olivera E, Martínez-Abaigar J, Tomás R et al (2004) Influence of temperature on the effects of artificially enhanced UV-B radiation on aquatic bryophytes under laboratory conditions. Photosynthetica 42:201–212. doi:10.1023/B:PHOT.0000040591.74037.f1
Palmer RA (1999) Detecting publication bias in meta-analyses: a case study of fluctuating asymmetry and sexual selection. Am Nat 154:220–233. doi:10.1086/303223
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annu Rev Ecol Evol Syst 37:637–669. doi:10.1146/annurev.ecolsys.37.091305.110100
Parmesan C (2007) Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Glob Chang Biol 13:1860–1872. doi:10.1111/j.1365-2486.2007.01404.x
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42. doi:10.1038/nature01286
Parton WJ, Morgan JA, Wang G et al (2007) Projected ecosystem impact of the prairie heating and CO2 enrichment experiment. New Phytol 174:823–834. doi:10.1111/j.1469-8137.2007.02052.x
Pounds JA, Fogden MPL, Campbell JH (1999) Biological response to climate change on a tropical mountain. Nature 398:611–615. doi:10.1038/19297
Richardson AJ, Schoeman DS (2004) Climate impact on plankton ecosystems in the Northeast Atlantic. Science 305:1609–1612. doi:10.1126/science.1100958
Ricker M, Gutiérrez-García G, Daly DC (2007) Modeling long-term tree growth curves in response to warming climate: test cases from a subtropical mountain forest and a tropical rainforest in Mexico. Can J For Res 37:977–989. doi:10.1139/X06-304
Root TL, Price JT, Hall KR et al (2003) Fingerprints of global warming on wild animals and plants. Nature 421:57–60. doi:10.1038/nature01333
Roy BA, Gusewell S, Harte J (2004) Response of plant pathogens and herbivores to a warming experiment. Ecology 85:2570–2581
Saether B, Tufto J, Engen S et al (2000) Population dynamical consequences of climate change for a small temperate songbird. Science 287:854–856. doi:10.1126/science.287.5454.854
Santos M (2007) Evolution of total net fitness in thermal lines: Drosophila subobscura likes it ‘warm’. J Evol Biol 20:2361–2370. doi:10.1111/j.1420-9101.2007.01408.x
Schmeller DS, Gruber B, Bauch B et al (2006) EuMon—Arten- und lebensraum-monitoring in Europa. Naturschutz Landschaftsplanung 39:384–385
Schmeller DS, Henry P-Y, Julliard R et al (2009) Advantages of volunteer-based biodiversity monitoring in Europe. Conserv Biol 23:307–316. doi:10.1111/j.1523-1739.2008.01125.x
Schtickzelle N, Baguette M (2004) Metapopulation viability analysis of the bog fritillary bitterfly using RAMAS/GIS. Oikos 104:277–290. doi:10.1111/j.0030-1299.2004.12825.x
Schwartz MD, Ahas R, Aasa A (2006) Onset of spring starting earlier across the Northern hemisphere. Glob Chang Biol 12:343–351. doi:10.1111/j.1365-2486.2005.01097.x
Shaw MR, Harte J (2001) Control of litter decomposition in a subalpine meadow-sagebrush steppe ecotone under climate change. Ecol Appl 11:1206–1223
Sherry RA, Zhou XH, Gu SL et al (2007) Divergence of reproductive phenology under climate warming. Proc Natl Acad Sci USA 104:198–202. doi:10.1073/pnas.0605642104
Sjögersten S, Wookey PA (2004) Decomposition of mountain birch leaf litter at the forest-tundra ecotone in the Fennoscandian mountains in relation to climate and soil conditions. Plant Soil 262:215–227. doi:10.1023/B:PLSO.0000037044.63113.fe
Skelly DK, Joseph LN, Possingham HP et al (2007) Evolutionary responses to climate change. Conserv Biol 21:1353–1355. doi:10.1111/j.1523-1739.2007.00764.x
Stenstrom A, Jonsdottir IS (2004) Effects of simulated climate change on phenology and life history traits in Carex bigelowii. Nord J Bot 24:355–371. doi:10.1111/j.1756-1051.2004.tb00850.x
Strayer DL (1999) Statistical power of presence–absence data to detect population declines. Conserv Biol 13:1034–1038. doi:10.1046/j.1523-1739.1999.98143.x
Svenning J-C, Skov F (2006) Potential impact of climate change on the northern nemoral forest herb flora of Europe. Biodivers Conserv 15:3341–3356. doi:10.1007/s10531-005-1345-8
Tews J, Ferguson MAD, Fahrig L (2007) Potential net effects of climate change on high arctic peary caribou: lessons from a spatially explicit simulation model. Ecol Modell 207:85–98. doi:10.1016/j.ecolmodel.2007.04.011
Thomas CD, Cameron A, Green RE et al (2004) Extinction risk from climate change. Nature 427:145–148. doi:10.1038/nature02121
Umina PA, Weeks AR, Kearney MR et al (2005) A rapid shift in a classic clinal pattern in Drosophila reflecting climate change. Science 308:691–693. doi:10.1126/science.1109523
Van Swaay CAM, Plate CL, Van Strien A (2002) Monitoring butterflies in the Netherlands: how to get unbiased indices. Proceedings of the section experimental and applied entomology of the Netherlands entomological society, vol 13, pp 21–27
Verburg P, Hecky RE, Kling H (2003) Ecological consequences of a century of warming in Lake Tanganyika. Science 301:505–507. doi:10.1126/science.1084846
Vilchis LI, Tegner MJ, Moore JD et al (2005) Ocean warming effects on growth, reproduction, and survivorship of southern california abalone. Ecol Appl 15:469–480. doi:10.1890/03-5326
Virkkala R, Heikkinen RK, Leikola N et al (2008) Projected large-scale range reductions of northern-boreal land bird species due to climate change. Biol Conserv 141:1343–1353. doi:10.1016/j.biocon.2008.03.007
Walker G (2007) A world melting from the top down. Nature 446:718–721. doi:10.1038/446718a
Walther G-R, Post E, Convey P et al (2002) Ecological responses to recent climate change. Nature 416:389–395. doi:10.1038/416389a
Whitfield SM, Bell KE, Philippi T et al (2007) Amphibian and reptile declines over 35 years at La Selva, Costa Rica. Proc Natl Acad Sci USA 104:8352–8356
Wiedermann MM, Nordin A, Gunnarsson U et al (2007) Global change shifts vegetation and plant–parasite interactions in a boreal mire. Ecology 88:454–464. doi:10.1890/05-1823
Wilson RJ, Gutierrez D, Gutierrez J et al (2007) An elevational shift in butterfly species richness and composition accompanying recent climate change. Glob Chang Biol 13:1873–1887. doi:10.1111/j.1365-2486.2007.01418.x
Winkler DW, Dunn P, McCulloch C (2002) Predicting the effects of climate change on avian life-history traits. Proc Natl Acad Sci USA 99:13595–13599. doi:10.1073/pnas.212251999
Wuethrich B (2000) How climate change alters rhythms of the wild. Science 287:793–795. doi:10.1126/science.287.5454.793
Yoccoz NG, Nichols JD, Boulinier T (2001) Monitoring of biological diversity in space and time. Trends Ecol Evol 16:446–453. doi:10.1016/S0169-5347(01)02205-4
Zavaleta ES (2006) Shrub establishment under experimental global changes in a California grassland. Plant Ecol 184:53–63. doi:10.1007/s11258-005-9051-x
Acknowledgments
This research was supported by the CNRS and the EuMon project (EU-wide monitoring methods and systems of surveillance for species and habitats of Community interest). The EuMon-project was funded within the 6th Framework of the European Commission. The authors are grateful to Murielle Richard, Priscilla McAllum as well as two anonymous reviewers who provided valuable comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lepetz, V., Massot, M., Schmeller, D.S. et al. Biodiversity monitoring: some proposals to adequately study species’ responses to climate change. Biodivers Conserv 18, 3185–3203 (2009). https://doi.org/10.1007/s10531-009-9636-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-009-9636-0