
Abstract
Rapid acceleration of industrial development in north-eastern British Columbia is currently occurring without a comprehensive assessment of the effects it will have on ecological or cultural systems. The cumulative effects of past development are already being felt within the Treaty 8 First Nations of the region but these effects have not been quantified from a landscape point of view. Using habitat modeling and GIS analysis the effects of 35 years of agricultural and industrial development on forest biodiversity and ecological integrity were investigated for a 410,000 ha landscape in north-eastern British Columbia. The study identified a significant increase in area impacted by developments which lead to a shift in landscape structure and significant change in forest biodiversity. Forest fragmentation and habitat loss has resulted in an increase in open, early seral and edge habitats at the expense of contiguous mature forests. The change in landscape structure resulted in a significant increase in species richness as generalist and early seral species responded positively to increases in these habitats. A significant increase in brown-headed cowbird parasitism risk was also detected. Changes in landscape structure, reduction in habitat for 22% of modeled species, increase parasitism and predation risk due to fragmentation, and increased access have resulted in a cumulative effect of recent resource development on ecological integrity that is both additive and synergistic. The propose acceleration of future development will increase the risk to maintaining the biodiversity and ecological integrity in the Peace-Moberly region of British Columbia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cumulative effects occur due to the spatial and temporal accumulation of change in an environmental system (Spaling and Smit 1993). They are generated by sequential and interactive activities over time and within the same space (MacDonald 2000). Cumulative effects can be additive, synergistic, or antagonistic (Gergel 2002). Synergistic effects occur when the combined effects are greater than the sum of individual effects (i.e. additive effects) while antagonistic effects occur when the combined effects are less than the sum of individual effects (Gergel 2002). MacDonald (2000) stated that additive effects are the most common in physical systems but that the complexity added by secondary and/or indirect effects can create synergism or antagonism. Cumulative effects are characterized by the interdependence between time, space, and activity which create complexity and impart the need to consider causality (Spaling and Smit 1993). Analyzing cumulative effects can be constrained by causality unless system interactions and indirect effects are identified (Dixon and Montz 1995). An assessment of cumulative effects therefore requires the integration of cause, process and effect into a systematic analysis that seeks to address the sources and pathways of cumulative environmental change and the resultant cumulative effects (Spaling and Smit 1993). The objective of a cumulative effects analysis is to scientifically assess the impacts of past, present and/or proposed human activities in a manner that can be incorporated into planning approaches to inform decision makers about the interactions between human and environmental systems (Spaling and Smit 1993; MacDonald 2000).
In Northeast British Columbia, the cumulative effects of uncoordinated and accelerated resource extraction activities are already causing major concern for the Treaty 8 First Nations within the region. They are concerned about changes in landscape structure, forest biodiversity, and what this means for the sustainability of their cultural resources. For many cultures, rapid and unnatural change to the environment can erode their beliefs and customs that are embodied in the environment they have known for thousands of years (Flannery 1994). After the one-off bonanza of the oil and gas rush, combined with increased forestry and agricultural development, what will be left of the forested landscape and what will be the state of biodiversity? To address this complex question a need exists to quantify the cumulative effects of resource extraction in order to justify the inherent link between cultural and ecological integrity.
The objective of this study is to assess the degree of change to landscape structure due to the cumulative effects of resource development and the response of faunal biodiversity so that decision makers can understand the degree of impact that past development has had on ecological integrity and what future development may contribute to.
The management of forest ecosystems and the conservation of biodiversity require managing ecological integrity (Dale and Beyeler 2001). Ecological integrity refers to the maintenance of structure, species composition, and the rate of ecological processes (Kimmins 2004). If conservation of biodiversity is an objective of achieving sustainable management than the maintenance of ecosystem structure requires management actions that maintain ecological integrity across all scales. The maintenance of biodiversity is regarded as an indicator of ecological integrity (Angermeier and Karr 1994). Management actions that modify ecosystem structures and/or result in land-use change will exacerbate our ability to maintain current biodiversity through its impact on ecological integrity (Sala et al. 2000; Hansen et al. 2001; Poore 2003). Of all management actions, land-use change is the greatest driver for change in biodiversity (Sala et al. 2000).
The Peace River regions of Northeast British Columbia and Northwest Alberta have been undergoing rapid land-use change since the 1990s as a result of the development of their oil and gas resources in addition to the continual development of a large commercial forestry and agricultural industry (Schneider et al. 2003). As a result of recurrent agriculture development, continual forest management and the rapid development of an oil and gas sector, the integrity of forest ecosystems are being threatened as forest landscapes become younger and more fragmented every year due to the cumulative development of these resource extraction activities (Schneider et al. 2003). The rapid encroachment of oil and gas, forestry and agriculture in Northern Alberta has caused a reduction in habitat quality of woodland caribou (Rangifer tarandus caribou) through the fragmentation of the Boreal forest landscapes by roads, seismic lines, and oil and gas wells (Dyer et al. 2001). In other regions, petroleum exploration has reduced habitat quality for vertebrate species through increased poaching, predation, and the spread of invasive species in areas with extensive seismic line development (Fiori and Zalba 2003). The development of roads for resource extraction is also regarded as a major threat to biodiversity conservation through the alteration of landscape structure due to the creation of sharp ecotones that result in edge effects and influence animal movement (Miller et al. 1996). The shift in landscape structure due to resource extraction activities has also been found to occur over time as development reduces the mean size of mid to late seral patches and increases the size of early seral patches (Williams and Marcot 1991). A shift in landscape structure towards younger more fragmented forests has been found to cause an increase in bird species that require early successional habitat and a reduction in species that require contiguous mature forest (Benkman 1993; Drapeau et al. 2000). If changes in land-use due to independent resource extraction activities can have a major influence on both stand- and landscape-level biodiversity than the cumulative effects of multiple activities should accentuate changes to biodiversity over time. This is even more likely when resource activities work in near absentia from each other as is the case in the Peace River region (Schneider et al. 2003).
Study area
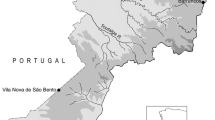
In this study, the cumulative effects of 35 years of oil and gas, forestry, and agricultural development on landscape structure and forest biodiversity are assessed within a 410,000 ha region within the Treaty 8 First Nation’s Traditional Territory in Northeast British Columbia (Fig. 1). The Peace-Moberly Study Region contains four broad ecosystems classified as biogeoclimatic (BEC) zones (Meidinger and Pojar 1991). The Boreal Black and White Spruce (BWBS) zone is the most dominant zone in the study area occupying 84.2% of the landscape. The Sub-Boreal Spruce (SBS) zone is the second most abundant ecosystem occupying 10.4% of the landscape. The Engelmann Spruce Subalpine Fir (ESSF) ecosystem occupies 5.3% of the landscape and the Alpine Tundra (AT) zone occupies only 0.1% of the landscape’s area.

Peace-Moberly study area within the British Columbian Treaty 8 First Nation’s Traditional Territory in Northeast British Columbia
Coniferous forests dominate 44% of the landscape with deciduous forests occupying 38% and mixed forests 18% respectively. Ten tree species are found within the study area: Hybrid white spruce (Picea glauca × engelmannii); white spruce (Picea glauca); black spruce (Picea mariana); trembling aspen (Populus tremuloides); Lodgepole pine (Pinus contorta var. latifolia); black cottonwood (Populus balsamifera ssp. Trichocarpa); paper birch (Betula papyrifera); subalpine fir (Abies lasiocarpa); western redcedar (Thuja plicata); and, tamarack (Larix laricina). Hybrid white spruce, lodgepole pine and trembling aspen are the dominant species with the region.
The Peace-Moberly study area is located within the Northern Rockies and Boreal Taiga Plains Bird Conservation Areas (Partners in Flight British Columbia and Yukon 2007). Partners in Flight British Columbia and Yukon (2007) and the Canadian Intermountain Joint Venture (2003) have identified 67 priority bird species that should be managed for within the study area. The B.C. Conservation Data Centre (BCDC) (2006) has also identified nine red-listed, 12 blue-listed and one identified wildlife species in the region. Four species that may be found within the study area are also listed in Canada’s Species at Risk Act (SARA) (BCDC 2006). The Treaty 8 First Nations have also identified two species that are culturally important, moose (Alces alces) and pine marten (Martes americanus). The study area is also within the geographic range of a species reliant on contiguous, mature-late seral forest, the southern red-backed vole (Clethrionomys gapperi) (Sullivan et al. 2001), which is also a major prey species for the pine marten and fisher (Martes pennanti) (Buskirk and Zielinski 2003). The Peace-Moberly is also within the geographic range of the brown-headed cowbird (Molothrus ater); the only obligate nest parasite in North America (Harris 1988; Campbell et al. 1997). These species are summarized in Table 1 and were used in this study to measure the cumulative effects of resource development on forest biodiversity.
Methods
A geographic information system (GIS) based analysis was used in this study to model the changes in landscape structure and the response of forest biodiversity over time. ArcGIS 9.2 (Environmental Systems Research Institute (ESRI) 2006) was used in conjunction with forest management and oil and gas development datasets to measure change in landscape structure and to model the suitability and distribution of each identified species on the Peace-Moberly landscape between two time periods. Raster coverages, with a spatial resolution of 0.25 ha (50 m × 50 m), were created for each variable used in the analysis. Table 2 summarizes the variables used and their characteristics.
Habitat variables
The raster coverages with a measure of zero or one were used to represent definitive habitat elements. This means that if a species requires a certain habitat element then it will only be found within the coverage of that habitat variable. For example, interior habitat is classified as 1.0 for interior-requiring species and zero for edge-requiring species. Edge habitat was defined as any area within 200 m of an existing urban or agricultural development, road, seismic, pipeline, power-line or harvested area with regeneration <5 m in height. Second-growth forests >5 m tall mediate edge effects on adjacent patches (Didham and Lawson 1999; Mesquita et al. 1999). Edge-effects penetration distances have been classified from 30 to 300 m with the most pronounced effects occurring within the first 100–200 m (Kroodsma 1984; Hansson 1994; Laurance et al. 2002). Any interior requiring species will not be found in areas defined as edge habitat. Riparian habitat was also classified as 1.0 for riparian obligate species while upland habitats were classified as 0.0. Riparian habitats were classified as any habitat within 100 m of a stream, lake or wetland since riparian obligate species can be present up to 100 m from riparian features although a significant decline is usually observed between 50 and 100 m (McComb et al. 1993; Gomez and Anthony 1996). This feature was also fixed over the modelled time periods as was the rock habitat cover and alpine habitat cover. For the habitat elements with a measure between zero and 100, a linear relationship between the proportion of tree species within a stand and habitat suitability was used until a threshold of 50% abundance was reached, at which point suitability was assumed to equal one. For shrub habitats, a suitability score of 100 was given to delineate areas since data on presence, not abundance, was available in the forest inventory database.
Past and current scenarios
The investigation into the cumulative effects of resource development requires the comparison of a past scenario to a current scenario. The current scenario represents an amalgamation of forest cover, topographic, predictive ecosystem mapping, forest road, and oil and gas development data from 1970 to 2004 for forest management based data and 1954 to 2007 for the oil and gas based data. The development of the past scenario was constructed to represent the state of the landscape pre-1970. The pre-1970 timeline was chosen because it represented the earliest recorded date of action within the forest cover database. All developments entered as occurring in 1970 were assumed to have occurred pre-1970. To represent past forest structure the age of each forest was reduced by 35 years (2004–1969). Crown closures were reclassified based on the average crown closure for stands with the same dominant species at the equivalent age in the current scenario. The resource development history of the landscape was used to identify which stands were harvested or cleared pre-1970. Stands that were harvested or cleared post-1970 were reconstructed using a neighbourhood analysis within ArcGIS. Seral stages were reclassified using the changes in age and crown closure based on the reconstructed landscape. Riparian, rock, alpine, and aspect coverages were kept fixed between scenarios. Edge habitat was created from the amalgamation of human activities on the landscape. For the current scenario: roads, pipeline, power lines, seismic lines, oil and gas well pads, harvested areas, and agricultural fields were combined. Roads and power lines, pipelines, not defined as polygons in any of the coverages, were assumed to have a 30 m right of way; a seismic line a width of 8 m, and oil and gas well pads a dimension of 120 × 120 m. The assumed widths for each linear variable were based on the average width of these features that are represented as polygons in the forest cover and predictive ecosystem databases. The well pad size is the maximum forest area allowed to be cleared for the well pad. These features were also used in the creation of the earl seral stage variable. The reconstructed pre-1970 forest landscape identified harvested and agriculture areas and some linear features. Oil and gas development data was used to identify the oil and gas wells that were developed pre-1970. Seismic lines were selected from these databases that were adjacent to and inclusive of the pre-170 well sites. Forest roads were removed from the forest road database that did not correspond to pre-1970 harvest blocks. This reconstruction of linear features was combined with the reconstructed forest landscape to create the pre-1970 edge/interior habitat map and seral stage distribution. Figure 2 illustrates the spatial change in edge and interior habitat between each time scenario while Fig. 3 summarises the landscape structure pre-1970 and in the current scenario.

Interior and edge habitat distribution for the (a) pre-1970 landscape; and (b) current developed landscape

Seral stage distribution, crown closure and amount of interior and edge forest habitat in the pre-1970 landscape and in the current landscape
Habitat modelling
Three types of habitat models were used to model species response: species-habitat matrices, habitat suitability modelling and habitat effectiveness modelling. Species-habitat matrices are tables that list vegetation types, seral stages, and habitat elements that wildlife species associate with and are based on expert opinion and/or quantitative studies (O’Neil et al. 2001). Habitat suitability modelling is a theoretical modelling approach that calculates single patch indices for each species based on a combination of habitat variables (Roloff et al. 2001). Habitat variables were mathematically combined using the raster calculator in ArcGIS to determine areas that represent unsuitable to optimum areas of habitat, respectively (United States Fish and Wildlife Service 1981). In this study, individual habitat models were created for each species from the species-habitat matrices. Eighty-six models were created for the 82 species; the winter ranges of moose, woodland caribou, wood bison and Rocky Mountain bighorn sheep were modelled separately from their non-winter ranges due to the critical importance of seasonal habitat supply for the conservation of the species (Lindenmayer and Franklin 2002). Habitat effectiveness modelling involves two components: (1) a measure of habitat quality; and, (2) a measure of habitat quantity (Roloff et al. 2001). The measure of habitat quality was calculated through the habitat suitability modelling described above, while the habitat quantity measure was calculated based on Davis et al. (2001). Habitat quantity was determined by classifying habitat into a quality index and then multiplying by the amount of habitat in each index to calculate habitat units (Roloff et al. 2001). The habitat suitability indexes calculated for each species were classified into four categories based on Davis et al. (2001):
-
1.
High Suitability = habitat suitability index of >67
-
2.
Medium Suitability = habitat suitability index > 33 to 67
-
3.
Low Suitability = habitat suitability index > 0 to 33
-
4.
Not Suitable = habitat suitability index = 0.00
Habitat units were then calculated for each modelled species using the following equation (Davis et al. 2001):
The total habitat units calculated for each species provide a quantitative estimate of the total amount of habitat present in hectares on a landscape (Davis et al. 2001).
Landscape-level richness
To determine if changes in stand- and landscape-level species richness has occurred over time an overlay analysis was conducted in ArcGIS using reclassified habitat suitability maps for all modeled species. Margalef’s Richness Index (Margalef 1958) was used to compare the change in stand-level species richness between time periods and then was weighted using the proportion of area occupied by stands with equal richness to get an overall estimate of landscape-level species richness. The equation for Margalef’s Richness Index is:
where S = number of species in an area and ln(N) = natural logarithm of the total number of species.
The weighted equation to determine landscape-level richness is:
where a = the area with S number of species and A = total area of the landscape.
Margalef’s Richness Index is purely a measure of species richness via presence or absence not species dominance as is the case with measures of diversity such as the Simpson’s, Brillouin and Shannon-Weiner Indexes (Chadwick and Canton 1984). Margalef’s richness index is sensitive to small sample sizes but with a sample size of 86 this issue should be mitigated in this study (Chadwick and Canton 1984).
Synergistic effects: risk of parasitism by the brown-headed cowbird
The risk of parasitism of passerine bird species within the study area by the brown-headed cowbird is a significant concern due the species response to land-use development. The cowbird has continually expanded its range since 1890 in British Columbia due to the opening up of forested landscapes by human development (Campbell et al. 2001). To measure change in the risk of brown-headed cowbird parasitism an overlay analysis was conducted in ArcGIS using bird species that are common hosts of the cowbird in British Columbia (Campbell et al. 2001) and the modelled habitat suitability of the cowbird. Risk of parasitism was classified by using the habitat suitability ratings as proxies of risk.
Species risk to cumulative effects
The World Conservation Union’s (IUCN) risk categories and criteria (IUCN 2001) were used to classify the degree of risk of cumulative effects to the modelled species. Species that suffered a reduction in habitat by ≥90% over the 35 years of development were classified as at extreme risk, very high risk if habitat reduction was ≥70%, high risk if a reduction of ≥30% occurred, and moderate if a reduction in habitat >0% and <30% occurred. Species that exhibit no change or an increase in habitat are classified as being of low risk, the category used to reflect “least concern” in this study.
Statistical analyses
To compare the pre-1970 landscape to the current landscape statistical tests were used to determine the significance of changes in area under development, species response, species richness, and the area at risk to parasitism. Statistical tests were also used to identify which habitat elements were independent of changes in the quantity of species habitat and which elements were not independent. A paired two sample of means t-test was performed to test for the significance of change in the area impacted by resource development and species richness between the pre-1970 and current scenarios. This t-test was selected because the landscape was tested twice and thus represented a natural pairing of observations. This t-test does not assume that the variances of both populations are equal and was used to determine if the means from each sampling period are distinct (Sokal and Rohlf 1995). A significance-level of 0.05 was used to infer a significant response. Single-factor Analysis of Variance (ANOVA) was used to determine if the habitat units between species with a negative and positive response over time were drawn from different sampling distributions of means (Tabachnick and Fidell 2001). The ANOVA test was also used to determine if the risk of brown-headed cowbird parasitism was different between time periods. Each time period was considered as an independent treatment. A significance-level of 0.05 was used to infer a significant response. A two-sample F-test for variances was used to test the difference in population variances for species habitat units between the two time periods (Sokal and Rohlf 1995). Each time period was used to represent a unique population. A one-tailed F-test was used because the response of the species in the current time period showed an increasing trend. A significance-level of 0.05 was used to infer a significant response. To determine which habitat elements best predicted changes in species habitat quantity two-way contingency tables were used to organize the categorical habitat element data (Gotelli and Ellison 2004). Thirty-one contingency tables were set-up to test the potential that change in a habitat element or group of habitat elements due to land-use development over time affected the habitat quantity of modeled species. The Pearson chi-square test for independence (Pearson 1900) was used to determine if a change in a habitat element was independent of a change in species habitat quantity (Gotelli and Ellison 2004). A significance-level of 0.05 was used to infer a significant response.
Results
Landscape structure
The cumulative effects of resource development over the last 35 years have resulted in a significant increase in the area affected by development (P: 0.0311). Figure 4 summarizes the change in area (hectares) over the last 35 years for each of the resource development activities included in this study. The increase in development coupled with the shift in seral stage distribution, crown closure and contiguous forest (Fig. 3) caused a change in the proportion of the landscape occupied by the habitat elements used in the modeling of forest biodiversity (Fig. 5). Land-use development coupled with natural changes in forest age have resulted in a decrease in mid-seral, moderate to closed forests and interior forest habitat. The cumulative effect of development has result in an 89% increase in edge habitat, 67% increase in early seral habitat, 49% increase in areas that could provide shrub habitat and a 47% increase in landscape openness. Changes in forest type also occurred with a large increase in mixed forests. The increase in stands that could provide habitat for both coniferous and deciduous forest species is responsible for the increase in stands that could support mixed-woods species. The increase in forest age of residual forest stands was responsible for the increase in mid to late seral stands. The change in habitat elements can provide a misleading picture of landscape structure. For example, Fig. 6 illustrates the change in landscape structure through the quantification of three metrics: mean patch size, maximum patch size, and number of patches. Thirty-five years of development has resulted in a decrease in mean patch size in all seral stages and a decrease in maximum patch size in each seral stage except the early seral stage. Development has also resulted in an increase in the number of patches within each seral stage. The decrease in mean and maximum patch size and increase in the number of patches also occurred for patches defined as interior forest habitat.

Change in area developed by resource extraction activities between pre-1970 and the current time period in the Peace-Moberly study area

Change in habitat elements between pre-1970 and the current time period in the Peace-Moberly study area

Change in landscape metrics between pre-1970 and the current time period in the Peace-Moberly study area
Response of forest biodiversity
Change in landscape structure had a significant impact on habitat quantity for the modeled species in this study (n = 86; P: 0.041). Of the 82 species modelled 22% experienced a decline in habitat quantity while 78% responded with an increase in habitat quantity. Ten percent of the species showed an increase of 5% or less. Figure 7 summarizes the responses of individual species. The change in habitat units between species that responded negatively to development and those that responded positively over time was found to be significantly different (F 19, 65; P < 0.001). Based on the IUCN (2001) risk classification criteria, four species would be considered to be at high risk and 14 a medium risk to the additive effects of 35 years of resource exploitation.

Response of forest biodiversity to 35 years of cumulative resource development: (a) species that responded with a decrease in habitat quantity; (b) species that responded with a 0.3–38% increase in habitat quantity; (c) species that responded with a 43–79% increase in habitat quantity; and (d) species that responded with a >79% increase in habitat quantity
Species richness
Species richness was significantly higher in the current time period compared to the pre-1970 scenario (P: 0.00574). Margalef’s richness index has increased from 8.7 to 10.8 over the last 35 years. The spatial distribution of species richness also changed over time in response to resource development; Fig. 8 shows the distribution of stand-level species richness over the landscape. In both time periods, riparian forests are the richest areas for forest faunal biodiversity. Richness has increased in regions that have undergone the most intensive development; predominately represented by the creation of edge habitat in the Boreal White and Black Spruce ecosystem.

Distribution of stand-level species richness across the Peace-Moberly landscape for (a) pre-1970; and (b) current developed landscape
Factors of habitat quantity change
The results of the chi-squared tests identified that 16 of the 31 habitat variables tested were not independent of species habitat quantity change. Table 3 summarizes the results of all 31 tests. The increase in early seral habitat, openness, edge habitat, mid-late seral habitat, and coniferous forest and the decrease in interior habitat and moderate to closed habitat where found to be the habitat variables that had a significant influence on species response. The decreases in interior-coniferous (interior habitat × coniferous forest interaction), interior-moderate-closed (interior habitat × moderate-closed stands interaction), and interior-mid-late seral forests (interior habitat × mid-late seral interaction) were found to be highly significant for species relying on these forest types. Decreases in coniferous forests with moderate to closed crown closure (coniferous forest × moderate-closed stands interaction) and increases in mid-late seral coniferous stands (coniferous forest × mid-later seral interaction) were also influential for species requiring these attributes in combination. Decrease in moderate to closed mid-seral stands (mid-late seral × moderate-closed stands interaction) also had a significant affect on changes in species habitat. Species requiring mid seral deciduous (mid seral × deciduous forest interaction) were also significantly impact by changes in this combination of habitat variables. Species that have a preference for open early seral habitats (early seral × openness interaction) responded to increases in the amount of this habitat combination on the landscape in the current scenario. Species responses to changes in the quantity of the remaining habitat elements or combination of habitat elements were found to be independent of their change.
Synergistic effects: development and parasitism risk
Figure 9 summarizes the species that are recognized as hosts of the brown-headed cowbird in British Columbia (Campbell et al. 2001) and the proportion of each bird species’ habitat at risk in both time periods. The area under risk to brown-headed cowbird parasitism was found to be significantly greater on the current developed landscape versus the pre-1970 landscape (P: 0.0489). In the pre-1970 landscape, 7.5% of the landscape was found to be at risk to parasitism which increased to 19% in the current developed landscape. Figure 10 illustrates the spatial distribution of the increase in parasitism risk due to cumulative resource development.

Proportion of bird species habitat at risk to nest parasitism by the brown-headed cowbird (BHCO) in the current and pre-1970 scenarios

Spatial distribution of brown-headed cowbird parasitism risk: (a) pre-1970 landscape; (b) current developed landscape
Discussion
Thirty-five years of observed land-use change has resulted in a significant transformation of landscape structure which in turn has caused additive, antagonistic, and synergistic cumulative effects within the Peace-Moberly landscape. The cumulative effects of resource development on landscape structure and the modeled response of forest biodiversity suggests that ecological integrity has been impacted. The measured decrease in mean patch size and interior forest and increase in the number of patches and edge habitat is congruent with other studies that investigated the impacts on development of forest landscapes (Tinker et al. 1998; McGarigal et al. 2001). The decrease in patch sizes of mid to late seral habitat and the increase in early seral habitat are also similar to studies that have quantified the effects of forest management and road development in forest landscapes (Williams and Marcot 1991; Miller et al 1996). The reduction in interior habitat and core patch size and the increase in the number of patches support the contention that habitat fragmentation has occurred over the last 35 years (Bender et al. 1998). Trzcinski et al. (1999) identified that the loss of forest cover, not fragmentation, as the most important factor that affects the distribution of forest species. In the Peace-Moberly habitat fragmentation has occurred in conjunction with habitat loss as industrial and agriculture development have replaced forest cover with open non-forested habitats. Bender et al. (1998) refer to the combination of habitat fragmentation and loss as habitat destruction which not only causes additive effects but synergistic effects on forest species. For example, nest parasitism and predation have been found to increase for both bird and ungulate species in landscapes fragmented by agricultural and industrial development (James and Stuart-Smith 2000; Lloyd et al. 2005). The measured increase in species richness in the Peace-Moberly is also a common observation in landscapes that have undergone increases in edge habitat and/or increases in open vegetation types due land-use development (Yahner 1988; Niemela et al. 1993). The increase in richness in this study was a result of an increase in open and early seral habitat in conjunction with edge habitat creation. Positive or neutral responses from species that prefer or are edge-interior habitat generalists are predicted in heavily developed landscapes (Bender et al. 1998).
This study has identified that the past 35 years of resource development has resulted in a significant increase in the amount of area developed which has resulted in changes in landscape structure and species richness. The modeled change in landscape structure and species richness is congruent with the responses observed in other landscapes in Western North America that have undergone intensive agricultural and industrial development. The cumulative effects of development on landscape structure are easy to detect and measure, the effects on forest species however are much more difficult to detect but important nonetheless for understanding changes in species assemblages that are masked by measures of species richness and diversity (Lindenmayer 1999).
Cumulative effects of development on forest species
The cumulative effects of human perturbation on faunal species at the landscape-level are difficult to detect or predict due to species reliance on specific habitat elements (Lindenmayer 1999). This means a species response over one time period to landscape alteration does not mean that its response in the future will be the same. This is particularly the case where multiple development activities have occured over a relatively short period of time (Lindenmayer 1999). Species specific habitat thresholds may be responsible in these cases. For example, species with high demographic potential can persist in a landscape with 25–50% suitable habitat, but species with low demographic potential may not be able to persist if suitable habitat falls below 80% in a landscape (With and Crist 1995). The lack of a negative response by some species may be attributed to past development not reducing suitable habitat below a threshold.
The decrease in interior habitat was one of the variables that significantly impacted the response of modeled species. Even when coupled with the measured increases in mid-late seral and coniferous habitat the reduction in interior habitat was a predominant cause of species decline. Schneider et al. (2003) identified that species requiring old-growth and interior habitats were likely to be the most effected by cumulative development, this study identified that interior species requiring mid-late seral coniferous habitat have already been adversely impacted by cumulative development. The negative response by interior species and the positive response by species that prefer or are edge-interior habitat generalists support the prediction of species response to landscape alteration by Bender et al. (1998). The response of species that require contiguous mature forest to the shift in landscape structure towards younger more open and fragmented forests is also supported by other studies that have investigated the impact of development on landscapes over time (Benkman 1993; Reed et al. 1996; Drapeau et al. 2000).
The modeling of species habitat suitability identified that species considered as threatened and culturally important have been negatively impacted by development. The blue-listed woodland caribou suffered a lost in winter and non-winter habitat. A decline in caribou habitat due to industrial development has been identified in adjacent boreal landscapes by Dyer et al. (2001). A decline in the habitat of the blue-listed grizzly bear, wolverine, fisher, surf scoter, and black-throated green warbler were also modeled. The decrease in habitat suitability was found to be linked to a reduction in contiguous mature and old-growth coniferous forests. The habitats for two culturally important species, moose and pine marten, were also modeled to have been reduced by development. Non-winter moose habitat was reduced by less than one percent but winter habitat was modeled to have declined by 7%. The reduction in winter habitat could have major impacts on moose populations since wintering habitats are a critical component of moose habitat on landscapes (Ahlén 1975; Peele et al. 1976). The reduction in contiguous mature coniferous forest was the cause of the decline in pine marten habitat. The Philadelphia vireo suffered the greatest reduction in habitat (63%) due a loss of mid seral deciduous forest. This species is being impacted by habitat loss not the fragmentation of the landscape and may serve as an example when the cumulative effects of habitat loss due to industrial development and forest management planning and operations do not consider the landscape-level impact of stand-based management. Habitat specialists are more sensitive to habitat fragmentation and loss than habitat generalists (Edenius and Sjöberg 1997). This study supports this hypothesis since species that have a narrow range of habitat requirements related to older intact forests were impacted the most. The pine grosbeak, red crossbill, Townsend’s warbler, varied thrush, yellow warbler, Wilson’s warbler, Vaux’s swift, and southern red-backed vole are species that all showed decline in habitat due to the cumulative effects of development. Of all these species, the red crossbill is the quintessential habitat specialist that has been found in other studies to be sensitive to habitat loss (Benkman 1993). This pattern remained the same in this study as did the response of old-growth and edge sensitive species (Bender et al. 1998).
For species that are not edge sensitive but require mature to old-growth forest a positive response to changes in landscape structure were detected. These responses should be considered with caution since they are driven by changes in forest age and structure. Even though these species exhibited an increase in habitat suitability their potential response was mitigated by land-use development. If future development continues as planned then habitat loss will likely increase for these species as continual increases in open, early seral habitat occurs at the expense of mature/old-growth forests. Species that should be considered at risk to future development includes the “identified” northern goshawk, the blue-listed Rocky Mountain bighorn sheep, northern long-eared myotis, and Canada warbler, and the red-listed wood bison, Connecticut, Cape May, and Bay-breasted warblers. The winter range for the wood bison is the critical habitat that would be impacted by habitat loss. Fifteen other species modeled in this study require mid-late or late seral forests and should be considered a risk to future habitat loss.
The effects of development on species requiring intact forest habitat was counterbalanced by the response of species that benefited from the increase in edge and/or open early seral habitat. The measured increase in species richness over time is driven by the response of these species. The increase in the habitat for these species and the decrease of species requiring intact forest habitat highlights the problem of only using indexes of species diversity or richness as a measure of impact. The use of only the richness index would have masked the cumulative effects of resource development on species in the Peace-Moberly (Lindenmayer 1999). The increase in species that favor open habitats comes at the cost of species that are adversely impacted by habitat destruction. Bennett (1990) identified a similar response in species assemblages in an intensively managed landscape in Australia. In Bennett’s study, resident species of the intact forest landscape were replaced by an array of new species adapted to the habitat characteristics of the developed landscape. The results of this study suggest a change in species assemblages is occurring at the landscape-level as habitat destruction alters the habitat in a negative manner for some species and positively for others. Campbell et al. (2001) suggested that the opening up of the boreal forest may increase the habitat for bird species requiring open habitats. They identified the mourning warbler, brown-headed cowbird and rose-breasted grosbeak as species that should respond positively to agricultural and industrial development. In this study, the habitat quantity for these species increased by 141%, 154% and 183%, respectively, supporting their hypothesis. The increase in brown-head cowbird habitat is a concern due to its role as an obligate nest parasite.
Many studies have been conducted to determine the impact of forest fragmentation on nest parasitism (Donovan et al. 1995, 1997; Hanski et al. 1996; Flaspohler et al. 2001). Hanski et al. (1996) and Flaspohler et al. (2001) found that nest predation had a greater impact on the nesting success of bird species versus nest parasitism. Flaspohler et al. found that 9.6% of studied nests were parasitized while Hanski et al. found that less than 3% of nests were parasitized in intact forest landscapes affected only by forest harvesting. Donovan et al. (1995) attributed increased nest failure in fragmented landscapes to increases in both nest predation and brown-headed cowbird parasitism. Donovan et al. (1997) found that brown-headed cowbird abundance is greater in highly fragmented landscapes but is correlated with host abundance. Despite differences in parasitism levels across North America it is generally accepted that increased fragmentation of landscapes will lead to reduced reproductive success for forest birds by increasing predation and parasitism (Donovan et al. 1995, 1997). In this study, the risk of nest parasitism was found to be significantly greater after 35 years of development that fragmented the Peace-Moberly landscape. The increased risk of nest parasitism due to an increase in habitat for both the brown-headed cowbird and it host species illustrates a potential synergistic affect on cumulative land-use development in the Peace-Moberly landscape. The increase in parasitism risk means that the habitat suitability modeling represents an increase in areas that may serve as ecological traps that will likely have a reduction in species abundance over time (Flaspohler et al. 2001). Inclusion of the increased rates of predation by avian and mammalian predators that are characteristic of fragmented landscapes than a significant amount of forest bird species are likely to be less abundant than the quantity of habitat that exists for them on the landscape suggests (Donovan et al. 1995; Flaspohler et al. 2001). The increase in predation in fragmented landscapes may also be an important determinant for the cause of observed decreases in caribou and moose within the region. James and Stuart-Smith (2000) found that increases in caribou mortality occurred due to increase in wolf and human predation in areas with roads, pipelines and seismic lines. Doak (1995) also found an increase in grizzly bear mortality in areas adjacent to roads. The increased access to areas provided by these linear structures is regarded as a serious conservation threat and it is likely that these linear disturbances are detrimental to other species subjected to predation, moose for example.
The Peace-Moberly region, like most of northeast British Columbia, is poised to undergo an accelerated increase in oil and gas development. The increased in development is expected to follow a “boom and bust” cycle over the next 20 years (Schneider et al. 2003). The increase in oil and gas exploration means more roads, seismic lines, pipelines and well sites. This will result in further habitat destruction occurring as forest habitats are lost and residual patches are fragmented even more. An increase in harvesting over the next five to ten years is also expected to occur in order to mitigate the impacts of a mountain pine beetle (Dendroctonus ponderosae) epidemic that is spreading throughout the region. Increases in coal mining and renewable energy development are also planned which may further impact forest ecosystems and forest biodiversity. Based on what has happened over the last 35 years we can therefore expect a reduction in interior forest habitat and an increase in open, early seral habitats. This means that species that have declined over the last 35 years will likely continue to decline and species that rely on mature forest habitats will begin to decline as habitat loss increases. Increased openness and edge habitats also means and increase in nest parasitism and predation as the landscape becomes one large ecological trap for forest species.
The objective of this study was to determine if resource development has had a cumulative effect on landscape-level ecological integrity. Changes in landscape structure, habitat destruction, change in forest biodiversity, an increase in parasitism risk, and a reduction in species habitat all support our hypothesis that agricultural and industrial development has had a additive and synergistic effect on the ecological integrity of the ecosystems within the Peace-Moberly study area. If changes in biodiversity do indeed represent a loss in ecological integrity than continual resource development over the foreseeable future will further erode ecological integrity. When the resource boom is over, the Peace-Moberly will be a much different landscape, lacking the ecological integrity needed to recover quickly to a state that can be used sustainably as a cultural, economical, social, and ecological resource by First Nations and stakeholders.
Conclusion
The increased rate of resource development in north-east British Columbia has placed amplified pressure on the ecological integrity of forest ecosystems within the region. To understand what the potential effects of future development it is important to incorporate the knowledge of past cumulative effects into decision making processes (Spaling and Smit 1993). Assessing the cumulative effects of multiple human perturbations on the ecological integrity of an ecosystem is one method that can be used to gain such knowledge. A change in forest biodiversity due to changes in landscape structure is one method that can be used for measuring ecological integrity. Understanding the role that cumulative effects have had on ecological integrity provides a foundation from which decisions and future actions can be evaluated. The incorporation of synergistic effects is also important for evaluating changes in ecological integrity since these impacts are likely to be greater than additive effects. The synergism between habitat loss and habitat fragmentation is example of how land-use changes can seriously impact forest biodiversity. The last 35 years of agricultural and industrial development in the Peace-Moberly region of northeast British Columbia can be defined as habitat destruction as landscape structure has been fragmented and converted to open, early seral and non-forested habitats. Some faunal species have undergone range reductions while others have had an expansion in potential habitat. These changes have resulted in a shift in species assemblages and an increase in species richness. Concurrent to these changes is an increase in risk to nest parasitism for some bird species and a likely increase in predation risk. If the past is anything to judge the future on then we can conclude that the cumulative effects of exploitive resource development has a significant impact on ecological integrity within the Peace-Moberly and that the future development will only exacerbate the loss of ecological integrity. For the decision makers in the areas under intensive development pressure, this study points to what has been done, and highlights the need for changes in management philosophies and approaches that incorporate the strategic concerns of those whose cultures rely on the ecological integrity of the region’s landscapes. If agricultural and industrial developments are not integrated into a sustainable management framework the future will be characterized by unabated habitat destruction leading to a loss in forest species and ecological integrity.
References
Ahlén I (1975) Winter habitats of moose and deer in relation to land use in Scandinavia. Swed Wildl 9:45–192
Angermeier PL, Karr JR (1994) Biological integrity versus biological diversity as policy directives: protecting biotic resources. Bioscience 44:690–687
B.C. Conservation Data Centre (2006) BC species at ecosystems explorer. B.C. Ministry of Environment. Victoria, BC. Available from http://srmapps.gov.bc.ca/apps/eswp/ (Accessed January 2007)
Bender DJ, Contreras TA, Fahrig L (1998) Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology 70:517–533
Benkman CW (1993) Logging, conifers, and the conservation of crossbills. Conserv Biol 7:473–479
Bennett AF (1990) Land use, forest fragmentation and the mammalian fauna at Naringal, south-western Victoria. Aust Wildl Res 17:325–347
Buskirk SW, Zielinski WJ (2003) Small and mid-sized carnivores. In: Zabel CJ, Athony RG (eds) Mammal community dynamics: management and conservation in the coniferous forests of Western North America. Cambridge University Press, Cambridge, UK, pp 207–249
Campbell RW, Smith GEJ, McNall MCE et al (1997) The birds of British Columbia, Volume 3: Passerines – Flycatchers through Vireos. UBC Press, Vancouver
Campbell RW, Dawe NK, McTaggert-Cowan I et al (2001) The birds of British Columbia, Volume 4: Wood warblers through Old World Sparrows. UBC Press, Vancouver
Canadian Intermountain Joint Venture (2003) Biological foundation and prospectus. British Columbia, Canada
Chadwick JW, Canton SP (1984) Inadequacy of diversity indices in discerning metal mine drainage effects on a stream invertebrate community. Water Air Soil Pollut 22:217–223
Dale VH, Beyeler SC (2001) Challenges in the development and use of ecological indicators. Ecol Indicators 1:3–10
Davis LS, Johnson KN, Bettinger PS et al (2001) Forest management: to sustain ecological, economic, and social values, 4th edn. Waveland Press, Long Grove, IL
Didman RK, Lawton JH (1999) Edge structure determines the magnitude of changes in microclimate and vegetation structure in tropical forest fragments. Biotropica 31:17–30
Dixon J, Montz BE (1995) From concept to practice: implementing cumulative impact assessment in New Zealand. Environ Manage 19:445–456
Doak DF (1995) Source-sink models and the problem of habitat degradation: general models and applications to the Yellowstone grizzly. Conserv Biol 9:1370–1379
Donovan TM, Thompson FR, Faaborg J et al (1995) Reproductive success of migratory birds in habitat sources and sinks. Conserv Biol 9:1380–1395
Donovan TM, Jones PW, Annand EM et al (1997) Variation in local-scale edge effects: mechanisms and landscape context. Ecology 78:2064–2075
Drapeau P, Leduc A, Giroux J-F et al (2000) Landscape-scale disturbances and changes in bird communities of the Boreal mix-wood forests. Ecol Monogr 70:423–444
Dyer SJ, O’Neill JP, Wasel SM et al (2001) Avoidance of industrial development by woodland caribou. J Wildl Manage 65:531–542
Edenius L, Sjöberg K (1997) Distribution of birds in natural landscape mosaics of old-growth forests in northern Sweden: relations to habitat area and landscape context. Ecography 20:425–431
Environmental Systems Research Institute (2006) ESRI ArcGIS 9.2, Environmental Systems Research Institute Ltd., USA
Fiori SM, Zalba SM (2003) Potential impacts of petroleum exploration and exploitation on biodiversity in a Patagonian nature reserve, Argentina. Biodivers Conserv 12:1261–1270
Flannery TF (1994) The future eaters. Reed New Holland, Australia
Flaspohler DJ, Temple SA, Rosenfield RN (2001) Species-specific edge effects on nest success and breeding bird density in a forested landscape. Ecol Appl 11:32–46
Gergel SE (2002) Assessing cumulative impacts of levees and dams on floodplain ponds: a neutral-terrain model approach. Ecol Appl 12:1740–1754
Gomez DM, Anthony RG (1996) Amphibian and reptile abundance in riparian and upslope areas of five forest types in western Oregon. Northwest Sci 70:109–119
Gotelli NJ, Ellison AM (2004) A primer of ecological statistics. Sinauer Associates, Inc. Sunderland, MA
Hansen AJ, Neilson RP, Dale VH et al (2001) Global change in forests: responses of species, communities and biomes. Bioscience 51:765–779
Hanski IK, Fenske TJ, Niemi GJ (1996) Lack of edge effect in nesting success of breeding birds in managed forest landscapes. Auk 113:578–585
Hansson L (1994) Vertebrate distributions relative to clear-cut edges in the boreal forest landscape. Landsc Ecol 9:105–115
Harris LD (1988) Edge effects and conservation of biotic diversity. Conserv Biol 2:330–332
James ARC, Stuart-Smith AK (2000) Distribution of caribou and wolves in relation to linear corridors. J Wildl Manage 64:154–159
Kimmins JP (2004) Forest ecology: a foundation for sustainable forest management and environmental ethics in forestry, 3rd edn. Prentice Hall, Upper Saddle River, NJ
Kroodsma RL (1984) Effect of edge on breeding forest bird species. Wilson Bull 96:426–436
Laurance WF, Lovejoy TE, Vasconcelos HJ et al (2002) Ecosystem decay of Amazonian forest fragments: a 22 year investigation. Conserv Biol 16:605–618
Lindenmayer DB (1999) Future directions for biodiversity conservation in managed forests: indicator species, impact studies and monitoring programs. For Ecol Manage 115:277–287
Lindenmayer DB, Franklin JF (2002) Conserving forest biodiversity: a comprehensive multiscaled approach. Island Press, Washington, DC
Lloyd P, Martin TE, Redmond RL et al (2005) Linking demographic effects of habitat fragmentation across landscapes to continental source-sink dynamics. Ecol Appl 15:1504–1514
MacDonald LH (2000) Evaluating and managing cumulative effects: processes and constraints. Environ Manage 26:299–315
Margalef DR (1958) Information theory in ecology. Gen Syst 3:36–71
McComb WC, Anthony RG, Newton M (1993) Small mammal and amphibian abundance in streamside and upslope habitats of mature Douglas-fir stands, western Oregon. Northwest Sci 76:7–15
McGarigal K, Romme WH, Crist M et al (2001) Cumulative effects of roads and logging on landscape structure in the San Juan Mountains, Colorado (USA). Landsc Ecol 16:327–349
Meidinger D, Pojar J (1991) Ecosystems of British Columbia. B.C. Ministry of Forests, Victoria, BC
Mesquita R, Delamônica P, Laurance WF (1999) Effects of surrounding vegetation on edge-related tree mortality in Amazonian forest fragments. Biol Conserv 91:129–134
Miller JR, Joyce LA, Knight RL et al (1996) Forest roads and landscape structure in the southern Rocky Mountains. Landsc Ecol 11:115–127
Niemela J, Langor D, Spence JR (1993) Effects of clear-cut harvesting on boreal ground-beetle assemblages (Coleoptera: Carabidae) in western Canada. Conserv Biol 7:551–561
O'Neil TA, Johnson DH, Barrett C et al (2001) Matrixes for wildlife-habitat relationships. In: Johnson DH, O’Neil TA (eds) Wildlife – habitat relationships in Oregon and Washington. Oregon State University Press, Corvallis
Partners in Flight British Columbia and Yukon (2007) Plans and implementation. Partners in Flight British Columbia and Yukon. Available from www.pifbcyukon.org (Accessed January 2007)
Pearson K (1900) Mathematical contribution to the theory of evolution VII: on the correlation of characters not quantitatively measurable. Philos Trans R Soc Ser A 195:1–47
Peele JM, Ulrich DL, Mackie JL (1976) Moose habitat selection and relationships to forest management in Northeastern, Minnesota. Wildl Monogr 48:1–65
Poore D (2003) Changing landscapes. International Tropical Timber Organisation and Earthscan Publications Ltd., London
Reed RA, Johnson-Barnard J, Baker WL (1996) Fragmentation of a forested Rocky Mountain Landscape, 1950–1993. Biol Conserv 75:267–277
Roloff GJ, Wilhere F, Quinn T et al (2001) An overview of models and their role in wildlife management. In: Johnson DH, O’Neil TA (eds) Wildlife–habitat relationships in Oregon and Washington. Oregon State University Press, Corvallis
Sala OE, Chapin FS, Armesto J et al (2000) Global Biodiversity Scenarios for the Year 2100. Science 287:1770–1774
Schneider RR, Stelfox JB, Boutin S et al (2003) Managing the cumulative impacts of land uses in the Western Canadian Sedimentary Basin: a modeling approach. Conserv Ecol 7:8
Sokal RR, Rohlf FJ (1995) Biometry: the principles and practice of statistics in biological research, 2nd edn. W. H. Freeman, New York
Spaling H, Smit B (1993) Cumulative environmental change: conceptual frameworks, evaluation approaches, and institutional perspectives. Environ Manage 17:587–600
Sullivan TP, Sullivan DS, Lindgren PMF (2001) Stand structure and small mammals in young lodgepole pine forest: 10-year results after thinning. Ecol Appl 11:1151–1173
Tabachnick BG, Fidell LS (2001) Using multivariate statistics, 4th edn. Allyn and Bacon, Boston, MA
The World Conservation Union (2001) IUCN Red List Categories and Criteria: Version 3.1. IUCN Species Survival Commission, Gland, Switzerland
Tinker BB, Resor CAC, Beauvais GP et al (1998) Watershed analysis of forest fragmentation by clearcuts and roads in a Wyoming forest. Landsc Ecol 13:149–165
Trzcinski MK, Fahrig L, Merriam G (1999) Independent effects of forest cover and fragmentation on the distribution of forest breeding birds. Ecol Appl 9:586–593
United States Fish and Wildlife Service (1981) Standards for development of habitat suitability index models. United States Fish and Wildlife Service Division of Ecological Services, ESM 103, Washington, DC
Williams B, Marcot B (1991) Use of biodiversity indicators for analyzing and managing forest landscapes. Trans N Am Wildl Nat Resour Conf 56:613–627
With KA, Crist TO (1995) Critical thresholds in species; responses to landscape structure. Ecology 76:2446–2459
Yahner RH (1988) Change in wildlife communities near edges. Conserv Biol 2:333–339
Acknowledgments
First, and most importantly, I would like to thank the British Columbian Treaty 8 Tribal Council for allowing me to conduct this research on their traditional territory and for providing information paramount to the success of this project. Their support and interests are at the foundation of this project. I would also like to thank the British Columbian government for providing information on forest and oil and gas development. I would also like to acknowledge the leadership and guidance of Dr. John Innes on this project. Finally, I would like to thank the Sustainable Forest Management Network for funding this research project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nitschke, C.R. The cumulative effects of resource development on biodiversity and ecological integrity in the Peace-Moberly region of Northeast British Columbia, Canada. Biodivers Conserv 17, 1715–1740 (2008). https://doi.org/10.1007/s10531-008-9376-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-008-9376-6