
Abstract
Guadalupe Island, an oceanic island in the northwest of Mexico, is an outlier of the California Floristic Province that has been disturbed by introduced goats for more than a century, with dramatic effects of goats on plant communities and local species extinctions. In 2004 the island went through a successful eradication program. Since then, six previously unrecorded species have been discovered and four supposed extinct species have been found again. Quantifying the true species richness of the island at the time of eradication, to set a benchmark for the future monitoring of this large-scale natural experiment, is both a challenge and a necessity. For this purpose, we estimated (a) current and (b) accumulated historical plant species richness of the island through accumulation functions. Estimation of current species richness was based on the geographical accumulation process of species richness (80 species) obtained from sampling 110 (50 m × 2 m) transects distributed along the island in year 2004. Historical species richness was estimated through the temporal accumulation of species richness (119 species) from botanical records (1,960 specimens reviewed) between 1875 and 2000. The predicted value of historical richness (213 species) is similar to known historical records (218 species), but estimation of current richness (203 species) is significantly higher than accepted extant plant richness (187 species). Our results suggest that currently there may be more plant species living in the island than estimated through recent botanical exploration. Future monitoring of the island as it recovers will clarify this hypothesis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
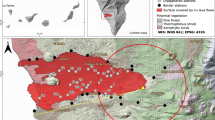
Guadalupe is an oceanic island of about 250 km2, with three small islets, located approximately 260 km off the coast of the Baja California peninsula in Mexico. It is the most remote and isolated unit of the California Islands (i.e., the Channel Islands of southern California, USA, and the Pacific islands of Baja California, Mexico; Fig. 1). Floristically, it is considered a geographic outlier of the California Floristic Province (Moran 1996).

The Pacific island corridor of the California Floristic Province, showing the regional location of Guadalupe Island
This remote volcanic island has become a fascinating natural experiment. Since its volcanic formation some 7 My ago (Moran 1996), Guadalupe has evolved in extreme isolation, resulting in a unique marine life with extraordinary local species like the Guadalupe fur seals (Arctocephalus townsendi, Hubbs and Rechnitzer 1958) and a highly endemic terrestrial biota, especially rich in vascular plants (Moran 1996; León de la Luz et al. 2003), lichens (Weber 1994), snails (Pilsbry 1927), and birds (Barrios-Quintana et al. 2006).
In 1602 Guadalupe Island was first-time sighted by the Spanish explorer Sebastián Vizcaíno (Berzunza 1950) and for approximately two centuries it served as occasional landfall for the Spanish galleons returning from the Philippines in their way to Cape San Lucas and Acapulco (du Petit-Thouars 1956). The biological consequences on the island of this early trans-Pacific trade are still unknown. From the late eighteenth century, and at least until 1830, the island was visited by fur seal hunters (Zavala-Gonzales and Mellink 2000), but it was in the mid-nineteenth century when the natural history of the island was dramatically changed forever when goats were introduced by sailors or fur seal hunters. Since then, the distinctive native flora and plant communities on the main island have been devastated by overgrazing, although, fortunately, the surrounding islets remained free of goats (Moran 1996; León de la Luz et al. 2003).
We never will know how Guadalupe Island was before its native flora was devastated by goat introduction. When Edward Palmer made the first botanical collections on the island in 1875, the destructive effects of goats on vegetation were already notorious (Watson 1876; Greene 1885; Francheschi 1893). Since then, scientific expeditions have visited periodically Guadalupe Island (see Moran 1996 for a detailed review). In 1996, Reid Moran published The flora of Guadalupe Island, Mexico, based on an extensive revision of previous botanical collections and on his twenty trips of floristic exploration during forty years (1948–1988).
In 2000, after a week-long bi-national expedition to Guadalupe Island organized by San Diego Natural History Museum, five new plant species were found (four exotic and one native species), but an alarming degradation of plant populations and communities was observed (Rebman et al. 2002; León de la Luz et al. 2003). An urgent call for goat eradication was made once more to avoid what seemed to be an impending ecological catastrophe of local extinction (León de la Luz et al. 2003).
Fortunately, a collaborative and multi-institutional program led by Mexico’s Instituto Nacional de Ecología (National Institute of Ecology a Mexican governmental research agency) and the Mexican Grupo de Ecología y Conservación de Islas (a non-governmental conservation group) was initiated in 2004 to eradicate the goats, restore the island, and manage it as a protected area. As a result, goats have been now completely eradicated, and in April 2005 Guadalupe Island was declared a Biosphere Reserve by the Mexican government.
As a fundamental part of this conservation plan, a vegetation baseline study was established in the spring of 2004, just before the eradication program began. Estimating the species richness of the island before eradication was considered an important reference point for future conservation, management, and long-term ecological studies. Since 1875, when Edward Palmer made the first botanical collections on the island, to present time there have been 223 different plant species documented on Guadalupe Island and its islets (Moran 1996; Rebman et al. 2002; León de la Luz et al. 2003; Junak et al. 2005; see Table 1). Due to overgrazing, two island endemics from this documented total are presumed to be extinct (Castilleja guadalupensis and Hesperalea palmeri), while 29 other species are supposed to have been extirpated from the island (locally extinct); 19 of them are native to the California Floristic Region and two of them are endemic to the California Islands. Thus, at present there is an estimated total of 192 vascular plant species on Guadalupe: 187 species are found on the main island, while five species survive exclusively on the islets (Moran 1996; Rebman et al. 2002; León de la Luz et al. 2003; Junak et al. 2005).
Observed species number has been widely recognized as a biased underestimation of the real species richness in a region (Hill 1973; Palmer 1990). Bias in floristic lists of known species from a region depends on how exhaustively explored and collected the area has been. In the case of Guadalupe Island, new species have been added in recent floristic expeditions: five previously unrecorded species were found in the 2000 expedition (Rebman et al. 2002; León de la Luz et al. 2003), other five new species were found by Steve Junak (Santa Barbara Botanic Garden Herbarium) in 2001–2005 (Junak et al. 2005), and one additional unrecorded species (Yabea microcarpa) was found in 2004 by Garcillán during field work. At the same time, it can be troublesome to clearly establish the fate of some plant species: four native species previously assumed to be extirpated from the island have been found in recent years (Junak et al. 2005).
These numbers suggest that some species may still be growing undetected in Guadalupe Island, and imply that, as the island vegetation recovers from goat overgrazing, new unrecorded species, or even recorded species presumed to be now extirpated or extinct may be found again. To address this question, in this paper we analyze (a) the extant and (b) the historical plant species richness through species-accumulation functions, based, respectively, on a systematic field sampling effort done in 2004 and on historical collection data obtained from herbarium records. Finally, we compare our results to accepted values of current and historical plant richness. We believe that these models may provide a powerful tool to understand and evaluate species extinction and extirpation in heavily degraded islands, and to guide restoration and conservation efforts.
Methods
Species-accumulation database
Extant plant species richness: geographical accumulation
We divided the island in 45 cells of 1.5′ latitude by 1.5′ longitude, and made three transects in each cell: one transect over the centroid of the cell, and the other two approximately 350 m north and south of the centroid (Fig. 2). Because of their rugged topography and/or small terrestrial area, some cells were sampled with only 1 or 2 transects, totaling 110 transects (50 m long × 2 m wide each) cumulatively covering 11,000 m2 (0.004% of the island’s total land surface). All transects were marked with a steel rod for future re-sampling and their exact location was registered with a GPS system. The presence of all vascular plant species was registered in each transect in May 2004, shortly after the end of a relatively moist winter-rain season. Identification of specimens was made with the help of Dr. Steve Junak (SBBG herbarium) and Dr. José Delgadillo (BCMEX herbarium). Voucher specimens were deposited in the BCMEX herbarium (Universidad Autónoma de Baja California–Ensenada). Nomenclature follows Moran (1996) and Jon Rebman’s unpublished taxonomic revision of the flora of Guadalupe Island (available from the Herbarium at the San Diego Natural History Museum). Because we were interested in generating a baseline estimate to monitor the success of the goat eradication program, we sampled only the main island and excluded the three islets from our study. A total of 80 plant species was recorded, and the data was organized into a presence/absence matrix of 80 species × 110 transects with a total of 1,173 presences.

Distribution of the 110 transects sampled in Guadalupe Island. Contour lines correspond to intervals of 200 m. Modified from INEGI (1998)
Historical plant species richness: temporal accumulation
We checked all specimens collected in the main island at four herbaria in California (CAS, DS, SD, and UC), which have been historically the main recipients of voucher specimens from research expeditions to the island, and registered information on the collector and collection date. After duplicates were eliminated, a total of 191 different species were found and 1,960 records registered. Because this analysis was aimed at estimating the historical flora of Guadalupe Island, we did not eliminate species currently considered extirpated (i.e., locally extinct). To condense the dataset, we sorted chronologically the 1,960 records and grouped them into 196 sequential groups of ten records each. We then constructed a frequency matrix of 191 species × 196 groups of ten time-ordered records. Each cell contained, for each species, the number of records with which the species was represented in a given chronological group of ten.
Data analysis: accumulation models
We tested four well-known accumulation models on the Guadalupe dataset: (a) power, (b) exponential, (c) Clench, and (d) negative exponential (see Tjørve 2003 for sources).
Extant plant species richness: geographical accumulation
We used number of transects as a measure of sampling effort. The expected species accumulation curve (sample-based rarefaction curves in the terminology of Gotelli and Colwell 2001) was built using the analytical method of Colwell et al. (2004) available in the computer program EstimateS version 7 (Colwell 2004). A rarefaction curve can be viewed as the statistical expectation of the corresponding accumulation curve over different random reorderings of the individuals or samples (Gotelli and Colwell 2001). The four models of species accumulation were tested against the rarefaction curve.
Historical plant species richness: temporal accumulation
We used historically-ordered collection specimens as a measure of sampling effort, and built the observed accumulation curve of number of species versus number of specimens collected over time. In this case, the specimens were not randomized because they maintain a sequential relationship through time. We used this sequential accumulation curve to fit the accumulation models.
Fitting models to our data
The predicted number of species by the Clench and negative exponential models corresponds to the value of their respective asymptotes, while the estimated richness for the power and exponential models is calculated through extrapolation. In the geographical accumulation process we extrapolated the sampled transect area to that of the whole area of the island. For the temporal accumulation process we arbitrarily extrapolated estimated species richness to a collection total of 2,940 specimens, that is, hypothesizing a 50% increase with respect to the current 1,960 collected herbarium specimens.
We compared the species predicted by the different models to the observed number of species for each accumulation process: 187 species for current species richness (geographical accumulation) and 218 species for historical species richness (temporal accumulation process). All models were fitted by direct numeric estimation of the parameters, using the Hooke and Jeeves algorithm of direct search (Ezcurra and Becerra 1987).
Results
Extant species richness: geographical accumulation
We recorded 80 plant species in our field transects (see Appendix for the species list). That is, 43% of the 187 species that conform the accepted current species pool was found in only 0.004% of the island’s total area. Mean richness per transect was 10.7 species, with a minimum value of 1 (in a transect heavily impacted by overgrazing and erosion) and a maximum value of 20. The distribution of species abundances, measured as the number of transects occupied by each species in the geographic dataset, and as the number of specimens collected for each species in the temporal dataset, were both extremely skewed. The majority of species for both datasets was ecologically rare: In the geographic dataset no species occurred in all transects and only one species (Hordeum murinum) was present in more than half of total sample (79 transects). In contrast, thirteen species (16% of the total number of observed species) were found in only one transect and another eight species (10% of the total) were found only in two transects.
The exponential model was the best descriptor of the geographical accumulation process, showing the highest R 2 value and a residual error significantly lower than those of any other model (Table 2, Fig. 3a). An extrapolated total of 203 species was predicted by this model, 16 species (8.6%) higher than the accepted current species richness of 187 species.

Accumulation curves. (a) Geographical accumulation against the number of transects: Fisher’s logarithmic model, (b) Temporal accumulation against the sequential number of collected specimens: Clench’s model. In (a) open dots show the rarefaction curve and black line shows the fitted model, and in (b) open dots show the accumulation curve and black line shows the fitted model
Historical species richness: temporal accumulation
We recorded a total of 1,960 vouchers, collected between years 1875 and 2000; containing 191 of the total historically recorded 218 plant species (88%). Twenty six species (14% of total 191 observed species) were represented by only one collected specimen in the herbaria consulted, 16 species (8% of total) had only two specimens, and 66 species (34%) were in the first decile (5 specimens or less) of the frequency distributions. Two species of particular interest to collectors (Perityle incana, a Guadalupe endemic, and Gilia nevinii, endemic to Guadalupe and the California Channel Islands) were represented by more than 40 herbarium specimens. The temporal accumulation curve showed a marked increase in accumulated species counts towards the end of the curve (Fig. 3b), caused by the collection of exotic species in the last recorded botanical expedition, organized by the San Diego Natural History Museum in 2000.
In this case, Clench’s model provided a significantly better fit, showing a significantly lower residual error, a larger R 2 value, and a more uniform distribution of the residuals than any other model (Table 1, Fig. 3b). The total of 213 plant species predicted by Clench’s model is very close to the historically recorded 218 plant species.
Discussion
Both datasets showed high levels of rarity; however, the species assemblages showing up as rare or abundant were different in both datasets (Fig. 4), suggesting that rarity in each accumulation process is determined by different factors. In the geographic accumulation process, rarity in the dataset is simply a result of the species abundance distribution in the field, and it can be explained by low overall abundances and/or patchy distributions of populations. In the case of historic accumulation, frequencies in the dataset seem to be highly influenced by the preference of collectors for some species that are judged to be more interesting for research, and possibly also by the access route to the island of different collection expeditions. As a general rule, endemic species that are rare and difficult to find in the field are overrepresented in the herbaria, while introduced weedy species that are common in the field are underrepresented in the collections.

Relative abundance of species in the herbaria dataset compared to the field dataset. Relative abundance is defined as number of specimens (herbarium data) or counts (field data) divided by the number of specimens or counts observed in the most abundant species in herbarium records or field transects, respectively. Black dots show weedy, non-native species, dark gray dots show species native to the California Floristic Province, light gray dots correspond to species endemic to the Pacific islands of the California Floristic Province, and white dots correspond to Guadalupe Island endemics. Note the bias in herbarium records for rare, endemic, or native species. The oval figure along the abscissa marks eight species that have maximum frequency in herbarium records but are very rare in the field: Cupressus guadalupensis, Spergularia macrotheca, Cryptantha foliosa, Deinandra greeneana and Perityle incana (Guadalupe endemics); Quercus tomentella, Phacelia floribunda and Gilia nevinii (Guadalupe and Channel Islands endemics)
Our fitted accumulation function for the field transects predicts that, currently, there are some 203 plant species living on the island, versus 187 species listed by collectors as still surviving there. This difference (16 species, or 8% of the currently accepted extant flora) between observed and predicted species richness suggests that (a) it is likely that more than the accepted 187 species may be surviving in Guadalupe Island at present, and that (b) some species considered extinct or extirpated may not have completely vanished, and/or that there may be species not previously collected.
In this sense, it is noteworthy that some native species previously assumed to be extirpated from the island have been found again in recent years, such as Allophyllum gilioides, Descurainia pinnata, Nicotiana attenuata, Satureja palmeri (Junak et al. 2005), and Juniperus californica (observed on the island by Garcillán and Vega in 2004). Furthermore, recent collecting has also added four previously unrecorded California-native species for Guadalupe Island (Ceanothus arborens, Claytonia parviflora, and Simmondsia chinensis reported by Junak et al. 2005; and Yabea microcarpa collected by Garcillán and Vega in 2004), and two exotic species (Lactuca serriola and Pennisetum setaceum reported by Junak et al. 2005).
It is very likely that species assumed to extirpated or not previously found may be surviving in remote and inaccessible parts of the island, where protection and refuge from goat grazing and trampling can be found, or that through overgrazing these species became scarce and sparsely distributed but now with goat eradication they may spread again. The two exotic species (Lactuca serriola and Pennisetum setaceum) found by Junak et al. (2005) in the proximity of the island’s airstrip are probably new invaders.
The full floristic listing of species collected on the island since 1875 (both extant and extirpated) is 218, while the historic species-accumulation model estimated a total of 213 species. The similarity between these two values, historically observed versus estimated richness, suggests that the historically collected flora is very close to the true historic flora. However, recently added new native species partially contradict this hypothesis. In spite of more than 125 years of botanical collections, some of the more rugged areas of Guadalupe Island may still be under-explored.
Our results show that although the historically accumulated species richness on Guadalupe Islands is most probably near or above 218 species, the extant species on the island probably add to more than the currently accepted number of 187. The accumulation functions we used predict that some 10-plus species assumed to be extirpated from, or previously unrecorded in the island may be surviving in remote areas. With the goats finally eradicated, as vegetation recovers future expeditions and monitoring will provide more insight into the predictive capacity of accumulation models and will help to confirm or reject hypotheses on the survival of plant species in both time and space within regions that are recovering from intense long-term perturbation.
References
Berzunza CR (1950) La Isla de Guadalupe. Bol Soc Mex Geogr Estad 70:7–62
Colwell, RK (2004) EstimateS: statistical estimation of species richness and shared species from samples. Version 7. User´s Guide and application. http://purl.oclc.org/estimates
du Petit-Thouars A (1956) Voyage of the Venus: soujourn in California. Excerpt from Voyage autour du monde sur la frégate Vénus pendant les années 1836–1839. Glen Dawson Press, Los Angeles
Ezcurra E, Becerra J (1987) Experimental decomposition of litter from the Tamaulipan cloud forest: a comparison of four simple models. Biotropica 19:290–296
Francheschi F (1893) Notes on the flora of Guadalupe Island. Zoe 4:130–139
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391
Greene EL (1885) Studies in the botany of California and parts adjacent. Bull Calif Acad Sci 1:179–228
Hill MO (1973) Diversity and evenness: a unifying notation and its consequences. Ecology 54:427–431
Hubbs CL, Rechnitzer AB (1958) A new fish, Chaetodon falcifer, from Guadalupe Island, Baja California, with notes on related species. Proc Calif Acad Sci Ser 4(29):273–313
INEGI (1998) Curvas de nivel de Baja California y Baja California Sur, escala 1: 250,000
Junak S, Keitt B, Tershy B, Croll D, Luna-Mendoza L, Aguirre-Muñoz A (2005) Esfuerzos recientes de conservación y apuntes sobre el estado actual de la flora de Isla Guadalupe. In: Santos del Prado K, Peters E (comp) Isla Guadalupe, restauración y conservación. Instituto Nacional de Ecología, pp 83–94
León de La Luz JL, Rebman J, Oberbauer T (2003) On the urgency of conservation on Guadalupe Island, Mexico: is it a Lost Paradise? Biodiv Conserv 12:1073–1082
Moran R (1996) The flora of Guadalupe Island, Mexico. Memoirs of the California Academy of Sciences, No. 19, California Academy of Sciences
Palmer MW (1990) The estimation of species richness by extrapolation. Ecology 71:1195–1198
Pilsbry HA (1927) Expedition to Guadalupe Island, Mexico, in 1922. Land and freshwater mollusks Proc Calif Acad Sci Ser 4(16):159–203
Qintana-Barrios L, Ruiz-Campos G, Unitt P, Erikson RA (2006) Update on the birds of Isla Guadalupe, Baja California. Western Birds 37:23–36
Rebman J, Oberbauer T, León de la Luz JL (2002) The flora of Toro islet and notes on Guadalupe Island, Baja California, Mexico. Madroño 49:145–149
Tjørve E (2003) Shapes and functions of species-area curves: a review of possible models. J Biogeogr 30:827–835
Watson S (1876) Botanical contributions. Proc Am Acad Arts Sci 11:105–148
Weber WA (1994) The lichen flora of Guadalupe Island, Mexico. In: Hochberg E (ed) Third California Islands symposium: recent advances in research on the California Islands. Santa Barbara Botanic Garden, Santa Barbara, CA, pp 65–71
Zavala González A, Mellink E (2000) Historical exploitation of the California Sea Lion, Zalophus californianus, in México. Mar Fish Rev 62(1):35–40
Acknowledgements
We are grateful to Luciana Luna and Francisco Casillas for their help in the field work, to Eulogio López for his assistance in checking the herbaria, to Charlotte E. González-Abraham for her work with figures, and to an anonymous reviewer for excellent criticism and advice. We also thank Secretaría de Marina de México and GECI for the facilities during field work. We are especially indebted to the CAS, DS, SD, and UC Herbaria staff for their invaluable help. The financial support that made possible this work was provided by Instituto Nacional de Ecología, México (INE). The second author (EE) thanks the financial support of the Packard Foundation and the Pew Fellowship Program on Marine Conservation.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
List of the 80 species found during field sampling in Guadalupe Island. Subspecies level is showed, although we used species as the taxonomic unit for our study. Among found species in the field only for Hordeum murinum have been recognized two different subspecies in Guadalupe Island (ssp. murinum and ssp. leporinum). Non-native taxa are marked with an asterisk (*).
Aizoaceae |
*Mesembryanthemum crystallinum |
*Mesembryanthemum nodiflorum |
Amaranthaceae |
Atriplex barclayana |
Aphanisma blitoides |
*Atriplex semibaccata |
*Chenopodium murale |
Apiaceae |
Lomatium insulare |
Arecaceae |
Brahea edulis |
Asteraceae |
Amblyopappus pusillus |
Ambrosia camphorata |
*Centaurea melitensis |
Deinandra greeneana |
*Hypochaeris glabra |
Logfia filaginoides |
Perityle emoryi |
Perityle incana |
*Sonchus oleraceus |
*Sonchus tenerrimus |
Uropappus lindleyi |
Boraginaceae |
*Capsella bursa-pastoris |
Guillenia lasiophylla |
Harpagonella palmeri |
Lepidium oblongum |
Pectocarya linearis ssp. ferocula |
*Plagiobothrys acanthocarpus |
*Plagiobothrys collinus |
*Sisymbrium orientale |
Thysanocarpus erectus |
Cactaceae |
Cylindropuntia prolifera |
Campanulaceae |
Githopsis diffusa |
Triodanis biflora |
Caryophyllaceae |
*Cerastium glomeratum |
*Herniaria hirsuta |
*Silene gallica ssp. capitatum |
Spergularia macrotheca |
Crassulaceae |
Crassula connata |
Cupressaceae |
Callitropsis guadalupensis |
Fabaceae |
Lotus grandiflorus |
Lupinus niveus |
Trifolium microcephalum |
Trifolium palmeri |
Trifolium gracilentum |
Vicia hassei |
Vicia ludoviciana ssp. ludoviciana |
Geraniaceae |
*Erodium cicutarium |
*Erodium brachycarpum |
*Erodium moschatum |
Hydrophyllaceae |
Emmenanthe penduliflora |
Eucrypta chrysanthemifolia |
Phacelia floribunda |
Phacelia phyllomanica |
Malvaceae |
*Malva parviflora |
Sphaeralcea palmeri |
Papaveraceae |
Eschscholzia elegans |
Plantaginaceae |
Plantago ovata |
Poaceae |
*Avena barbata |
*Bromus diandrus |
*Bromus hordeaceus |
*Bromus madritensis ssp. rubens |
*Bromus trinii |
*Hordeum murinum ssp. murinum/ssp. leporinum |
*Lamarckia aurea |
Muhlenbergia microsperma |
Phalaris caroliniana |
*Schismus barbatus |
*Vulpia myuros |
Vulpia octoflora |
Polemoniaceae |
Gilia nevinii |
Leptosiphon pygmaeus ssp. pygmaeus |
Polygonaceae |
Pterostegia drymarioides |
Portulacaceae |
Calandrinia ciliata |
Calandrinia maritima |
Claytonia perfoliata ssp. mexicana |
Resedaceae |
Oligomeris linifolia |
Rosaceae |
Aphanes occidentalis |
Rubiaceae |
*Galium aparine |
Solanaceae |
Lycium californicum |
*Nicotiana glauca |
Themidaeceae |
Dichelostemma capitatum ssp. cinerea |
Urticaceae |
Parietaria hespera |
Rights and permissions
About this article
Cite this article
Garcillán, P.P., Ezcurra, E. & Vega, E. Guadalupe Island: Lost paradise recovered? Overgrazing impact on extinction in a remote oceanic island as estimated through accumulation functions. Biodivers Conserv 17, 1613–1625 (2008). https://doi.org/10.1007/s10531-008-9370-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-008-9370-z