
Abstract
Programs for monitoring biological diversity over time are needed to detect changes that can constitute threats to biological resources. The convention on biological diversity regards effective monitoring as necessary to halt the ongoing erosion of biological variation, and such programs at the ecosystem and species levels are enforced in several countries. However, at the level of genetic biodiversity, little has been accomplished, and monitoring programs need to be developed. We define “conservation genetic monitoring” to imply the systematic, temporal study of genetic variation within particular species/populations with the aim to detect changes that indicate compromise or loss of such diversity. We also (i) identify basic starting points for conservation genetic monitoring, (ii) review the availability of such information using Sweden as an example, (iii) suggest categories of species for pilot monitoring programs, and (iv) identify some scientific and logistic issues that need to be addressed in the context of conservation genetic monitoring. We suggest that such programs are particularly warranted for species subject to large scale enhancement and harvest—operations that are known to potentially alter the genetic composition and reduce the variability of populations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Methods for monitoring biodiversity at different biotic levels, from genes to ecosystems, are necessary for meeting the primary goal of conserving and sustainably using biological resources as outlined in the convention on biological diversity (CBD; www.cbd.int). This convention, together with the United Nations Framework Convention on Climate Change, represent key agreements adopted at the 1992 Earth Summit in Rio de Janeiro. The CBD is presently the most important international political instrument dealing with the increasing threat of biodiversity loss. Currently, 190 nations are parties to the convention, and the CBD shapes the political process with respect to biological diversity—from the genetic level to ecosystems—for most of the world, including the European Union.
Article 7 of the CBD recognizes the need to “identify components of biological diversity” and to “monitor, through sampling and other techniques, the components of biological diversity” in order to reach the goal of conservation and sustainable use of biological resources. It will not be possible to detect negative changes and reductions of biological diversity unless the amount and distribution of this diversity is systematically studied over time.
Programs for monitoring biodiversity at the levels of ecosystems and species exist in several countries (Estonian Ministry of Environment 1997; Paulsen 1997; Rasmussen and Geertz-Hansen 1998; Norwegian Agency for Nature Conservation 1998; Swedish Environmental Protection Agency 1999; Martins et al. 2007), but programs for monitoring changes in genetic composition and diversity are missing, although the need has been stressed repeatedly (e.g. UNEP 1995; Laikre and Ryman 1997; Usher 2002; Aho and Laikre 2004; Andersson et al. 2007).
In this paper we discuss issues concerning the development of programs for monitoring gene level biodiversity within the CBD framework using Sweden as an example. There is a strong political focus on issues relating to biological diversity and the implementation of the CBD in Sweden. It also appears as though Sweden is relatively progressive with respect to recognizing the need for identifying and monitoring genetic diversity. Recently adopted political goals explicitly state the importance of retaining “sufficient genetic variation” to assure long term viability of particular species (Sweden’s environmental objectives; http://miljomal.nu/english/english.php; Andersson et al. 2007).
We identify information on genetic composition and spatial population genetic structure as important starting points for development of monitoring that aims to detect reductions of gene level variability. Such information describes the magnitude and the distribution of the genetic diversity to be monitored. Similarly, information on the degree of temporal stability of genetic structures is important to permit separation of “normal” variation over time from changes that may constitute a threat to the genetic resources. We review the extent of such data with respect to naturally occurring animal and plant populations in Sweden.
Many natural populations are subject to human activities that may negatively affect their genetic composition and variability. In addition to habitat alterations, large scale harvest and release programs are carried out for several forest trees, fishes, and wildlife populations. Such operations can change the natural genetic makeup through selective removal and/or addition of genotypes (Laikre and Ryman 1996; Ernande et al. 2003; Grift et al. 2003; Olsen et al. 2004). Also, over-harvest may result in extinction of local gene pools, or may reduce the genetically effective population size (N e ) through manipulation of the demographic characteristics of a population (Ryman et al. 1981; Laikre and Ryman 1996). Reductions of N e will result in elevated rates of genetic diversity loss.
Releases of captively cultivated individuals may similarly result in manipulation of reproductive rates which, in turn, may reduce N e (Ryman and Laikre 1991; Ryman et al. 1995; Wang and Ryman 2001). Recent studies show that extensive release of genetically alien populations is common in Sweden, but data regarding the potential effects of these manipulations are lacking (Laikre and Palmé 2005; Laikre et al. 2006, 2007). The situation is similar in many other countries.
We identify key categories for species for which we regard conservation genetic monitoring as highly warranted, and propose that monitoring species subject to large scale exploitation is particularly urgent. We suggest pilot species to observe for temporal genetic change based on our review of currently available genetic information. We also discuss gaps in the general understanding of short term temporal genetic change and suggest additional information needed in this context.
Conservation genetic monitoring
Genetic monitoring may be used for a wide variety of ecological and management purposes, all of which do not necessarily deal with the retention of genetic diversity (Schwartz et al. 2007). Here, we focus on the type of monitoring of interest for implementation of the CBD, i.e. for identifying and safeguarding gene level biodiversity. We define conservation genetic monitoring to imply the systematic survey of amount of genetic variation, genetic composition, and spatial genetic structure with the aim of detecting potential changes of these parameters that may reflect or result in loss of gene level variability.
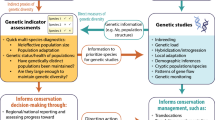
Figure 1 schematically illustrates major threats to genetic diversity and possible effects of these threats that conservation monitoring programs should aim to detect. This is largely comparable to the Category II type of monitoring identified by Schwartz et al. (2007) that focuses on population genetic parameters reflecting levels of genetic variation, rate of loss, admixture, population structure, and migration.

Schematic illustration of the threats to genetic variability and the possible effects that need to be monitored to meet the aims of the convention on biological diversity. Various human induced pressures may result in (i) loss of genetically distinct local populations or population segments, (ii) reduction of genetically effective population sizes (N e ) causing increased rates of loss of genetic variation through genetic drift, (iii) change of genetic composition and loss of genetic variation through selection, or (iv) inflow of alien genes
We regard information on the genetic composition of particular species over their geographic distribution an important prerequisite for conservation genetic monitoring. This information provides a starting point, which the genetic characteristics may be compared with at later points in time. In a next step, programs must be developed that permit detection of “unnatural” amounts of change potentially reflecting threats to genetic diversity (Fig. 1). It must be possible to separate normal rates of genetic change from elevated ones caused by anthropogenic pressures. In this respect, knowledge on “natural” rates of genetic change is important.
Collection of information
To obtain information on what is currently known regarding genetic composition and spatio-temporal patterns of natural animal and plant populations in Sweden we searched the literature using four databases: Science Citation Index (ISI Web of Science), BIOSIS, AGRIS, and CAB. As search criteria we used the following string: “natural population” OR “genetic variation” OR “genetic variability” OR “population genetic*” OR “genetic differen*” OR “population structure” OR “genetic distance” OR “genetic divergence” OR “genetic structure” to occur together with “Sweden” OR “Swedish” OR “Scandinavia” OR “Baltic”, in the title, abstract or keywords. The “*” signify wild card character(s). The search was conducted in August 2006.
Of course, it is difficult to construct search criteria that result in a completely exhaustive retrieval, and we do not claim that the present bibliography includes each and every study on genetic variability on natural animal and plant species in Sweden. We believe, however, that the general pattern of well studied and less well studied species is representative and that most of the scientifically published work is included here.
We were primarily interested in genetic variability of natural populations that may be monitored at particular gene loci, and publications on domesticated species/populations and quantitative genetic studies were therefore excluded. To obtain information on the extent to which the genetic composition of Swedish populations have been studied over time we screened references for the words “temporal”, “long-term”, “short-term”, “time” or “ancient” in the titles or abstracts of the previously retrieved references. We focused on species occurring “naturally” in Sweden, implying those that have existed as self sustaining populations in the wild for about 150 years or more (Berg and Nilsson 1997).
We were particularly interested in the degree of genetic information on species subject to large scale exploitation involving breeding-release and/or harvest. From the genetic perspective, harvest and enhancement represent the removal or addition of individuals with particular phenotypes and genotypes. Such manipulations may have profound effects on the genetic characteristics of the population (Laikre and Ryman 1996). The potential conservation genetic problems posed by these activities have been recognized primarily within the field of fisheries management (Laikre and Palmé 2005; Laikre et al. 2006, 2007), and mapping of the genetic variability patterns of natural and hatchery populations have in some cases been carried out by local authorities and hatcheries in Sweden. Similar screenings do not appear to occur for forest trees and game birds. We contacted the County Administrative Boards, that issue permits for local fish releases, to obtain information on genetic surveys of stocks used for release.
Information on spatial genetic structure
We found a total of 775 scientific publications that deal with the population genetic characteristics of one or several species that occur naturally in Sweden using various molecular genetic techniques. In total, 374 species have been studied in the 775 publications, and for the vast majority of species (246), there is only one single scientific publication. Only for 29 species are there more than four scientific studies (Fig. 2).

Number of scientific studies for the 29 Swedish species for which five publications or more were found. Four or fewer studies were found for another 345 species. A full reference list is available at www.zoologi.su.se/research/popgen/monitoring
The most frequently studied species in Sweden are brown trout Salmo trutta (68 publications), Scots pine Pinus sylvestris (61), Atlantic salmon Salmo salar (58), Arctic char Salvelinus alpinus (24), and Norway spruce Picea abies (21). Clearly, a few fishes and forest trees have dominated the population genetic research, whereas there are no, or very few, publications for most taxa. For instance, there are only 43 studies in total of birds (30 species), and 56 studies covering a total of 44 insect species. Figure 3 illustrates the number of genetic studies of different taxonomic groups. Lists of all references and the taxa studied are available at www.zoologi.su.se/research/popgen/monitoring.

The number of genetic studies for different taxonomic groups in Sweden. In total, 775 publications involving 374 species were found in the present literature search
Information on temporal genetic variation
With respect to studies including two or more temporally separated samples we found a total of 32 such studies involving natural Swedish populations of 15 different species (Table 1). About half (16) of the studies focus on bony fishes, and eight of those involve the brown trout. Other bony fishes studied temporally are Atlantic herring (3 studies), turbot (2), Atlantic cod (1), Arctic char (1), and European eel (1). Equally well studied (measured by the number of studies) as the brown trout is the grey wolf. Eight studies focus on change of diversity over time in the very small Swedish population (less than 100 individuals during most of the time span) and on the effects of those changes. Arctic fox (1 study) and brown bear (1) are the other mammals that have been studied temporally.
We found only a single temporal genetic study of a forest tree (Scots pine), as for perennial flowering plant (crow garlic), and two on each of birds (great reed warbler and willow grouse) and insects (spear-winged fly and fruit fly). The periods covered in these studies vary between 2 and 9,900 years, with ten investigations including 2–4 years. Several studies that cover long time spans involve comparisons between two or a few points in time. Only four studies systematically follow consecutive cohorts and thus provide detailed information on temporal genetic change. All of these are on the brown trout.
In summary, only a few species of the Swedish fauna and flora have been well studied with respect to genetic composition and variability patterns. For the vast majority of Swedish species, however, genetic information is missing completely (cf. Laikre and Ryman 1997). The lack of information on the temporal stability of observed structures is striking—of the 775 studies we retrieved, only 4% include temporally separated samples. Similarly, the extent of information for the 374 studied species varies considerably, but it appears that the number of studies for separate species largely reflect the level of knowledge for those species.
Information on species subject to large scale exploitation
Examples of species that are subject to large scale harvest and/or enhancement operations are listed in Table 2 together with the number of genetic studies that are available for these species. Clearly, basic information on spatio-temporal genetic structure is missing for most exploited species, making it difficult or impossible to evaluate the degree of genetic sustainability of these resources. For several highly exploited species, including the Atlantic salmon, Norway spruce, Atlantic cod, Atlantic herring, and brown trout, extensive genetic data are available (Fig. 2, Table 2). For these five species, conservation genetic monitoring programs should be developed to evaluate potential effects of the harvest and enhancement activities (cf. Laikre et al. 2006).
The fish species for which most release permits are issued in Sweden is the brown trout. During the period 1995–2001 over 60% of the issued permits for fish release (6,877/11,157) concerned brown trout (Laikre and Palmé 2005; Laikre et al. 2006), and around 800,000 brown trout are released annually, although the exact number of released fish is not recorded (cf. Laikre and Palmé 2005; Olsson et al. 2007).
The information collected from the County Administrative Boards, the National Board of Fisheries, and various published reports shows that more than 90 separate stocks of brown trout have been used for stocking. All of these stocks, except one (the Konnevesi stock from Finland), originate from Sweden. However, over 40% of the stocks have been released in geographic regions outside their original distribution. This is expected to cause genetic changes in recipient wild populations as the brown trout shows high levels of genetic divergence between watersystems and even between spawning areas within watersystems (e.g. Laikre 1999). Only 40% of the released brown trout stocks have been studied genetically.
The Atlantic salmon is subject to the most extensive fish stockings in Sweden in terms of number of fish released. At least 3 million individuals are released annually. The majority of these fish are stocked into rivers flowing into the Baltic Sea, where an estimated 80–90% of the total salmon population originates from hatcheries (WWF 2001; Swedish National Board of Fisheries 2007). Around 30 different stocks are used for these releases, and all of them have Swedish origin. However, as with the brown trout, Atlantic salmon is frequently released in non-native areas within Sweden. Genetic screening has been performed on 75% of these stocks, but the genetic effects of the large scale releases are not monitored. A full reference list and a table of the stocks of brown trout and Atlantic salmon used for release in Sweden is available at www.zoologi.su.se/research/popgen/monitoring.
Development of genetic monitoring programs
Clearly, within the foreseeable future it will not be possible to monitor all species for loss of intraspecific variability. Rather, conservation monitoring programs developed within the CBD framework will have to focus on particular cases. Pursuing along the lines that we have proposed previously (Laikre and Ryman 1997), we suggest the following categories of species for which monitoring genetic diversity is particularly urgent. Development of monitoring programs for individual target species within these categories seem to be a rational first step in this context. Examples of Swedish species within categories for which basic spatio-temporal genetic information appear to be available is also presented.
Category 1: Species subject to large scale release operations
Release of genetically alien populations into areas with wild conspecifics is expected to cause changes and possible losses of natural gene pools (Fig. 1; Ryman et al. 1995; Laikre and Ryman 1996; Allendorf and Luikart 2007). Similarly, in cases where local stocks are used for breeding and subsequent release, loss of genetic variability of the natural population may occur due to the so called supportive breeding effect—a reduction of effective population size resulting from demographic manipulation of family size (Ryman and Laikre 1991; Ryman et al. 1995; Wang and Ryman 2001).
Currently, the genetic effects of large scale release programs in Sweden, as elsewhere, are unknown (Laikre et al. 2006). Genetic monitoring is urgently needed in this context, and we suggest possible target species within this category to include brown trout (Salmo trutta), Norway spruce (Picea abies), Atlantic salmon (Salmo salar), and grey partridge (Perdix perdix) or mallard (Anas platyrhynchos). For the latter bird species genetic information is presently missing, but both the mallard and the grey partridge are subject to large annual releases for hunting purposes where a substantial proportion of the released birds are imported to Sweden (Laikre and Palmé 2005; Laikre et al. 2006). Monitoring potential genetic effects of these releases is highly warranted.
Category 2: Species subject to large scale harvesting
Harvest from natural populations is practiced for many animal and plant species and frequently constitutes a part of the management of those populations. Harvest implies the selective removal of particular phenotypes or demographic groupings, and such removal has been demonstrated to cause genetic effects (Ryman et al. 1981; Harris et al. 2002; Ernande et al. 2003; Grift et al. 2003; Olsen et al. 2004). Samples for genetic screening should be easy to obtain from species that are regularly killed in large numbers. Potential candidate species in this category include Atlantic cod (Gadus morhua), Atlantic herring (Clupea harengus), Atlantic salmon (Salmo salar), and moose (Alces alces).
Category 3: Species classified as “near threatened” or “least concern”
Declining population size and increased isolation of population segments is expected to be coupled with reduction of the genetically effective population size, and thus result in elevated levels of genetic drift and inbreeding. Species which are not yet identified as threatened but e.g. classified as near threatened (NT) or least concern (LT) following the IUCN threat categories (www.redlist.org) may nevertheless face loss of intraspecific variability that has not yet been recognized. Genetic monitoring of this category of species may be warranted to obtain an appropriate picture of the level of genetic threat. Targets in Sweden include species that are identified as near threatened, such as brown bear (Ursus arctos), great reed warbler (Acrocephalus arundinaceus), and turbot (Psetta maxima).
Category 4: Species with small population sizes
For species with small population sizes, around 1,000 individuals or less, we do not need genetic monitoring to conclude that genetic variability is lost. The genetically effective population sizes of such populations is necessarily considerably smaller and below the threshold value of 500–5000 recognized as necessary to retain long term evolutionary potential (Franklin and Frankham 1998; Lynch and Lande 1998; Allendorf and Ryman 2002). Thus, genetic monitoring of this type of species may be carried out primarily for evaluating the effects of various measures for increasing population size, avoiding removal of genetically important individuals, and for documenting effects of inbreeding.
Category 5: Species which are subject to other types of monitoring
Several species in Sweden are monitored with respect to occurrence and abundance or to reflect environmental contaminants. Such programs frequently involve regular collection of tissue samples and other information on the sampled individuals. For instance, within the scientific bird banding project in Sweden over 300,000 birds representing around 200 species are caught and banded annually. Similarly, aquatic organisms such as the benthic amphipods Monoporeia affinis and Pontoporeia femorata are regularly sampled for environmental monitoring of the Baltic Sea, and various fish species are caught in freshwater lakes subject to species monitoring programs conducted by the National Board of Fisheries. Extending such particular programs to also include genetic diversity monitoring would provide means for coupling various types of demographic and environmental information to genetic variability patterns and change of genetic profile. Possible target species in this category include Monoporeia affinis, collared flycatcher (Ficedula albicollis), brown trout (Salmo trutta) and Arctic char (Salvelinus alpinus). However, basic genetic information on population structure is presently available only for the last three species.
Category 6: Species for which extended time series of samples are available
There is a general lack of knowledge on microevolutionary processes including natural rates of genetic change over short time periods, and fluctuations of effective population sizes in natural populations. Increased scientific efforts are needed on these topics (see below). In this respect, important information may be obtained from monitoring species for which considerable time series of genetic data and samples exist. In Sweden, such data are available for e.g. several brown trout (Salmo trutta) populations.
Tissue collections
Tissue samples are regularly collected for particular species and stored at various institutions within the framework of several museum programs, research activities, and wildlife monitoring projects. For instance, collections of a large number of different species are kept by the National History Museum, the National Board of Fisheries, and several university departments in Sweden. These collections constitute important material from a conservation genetic monitoring perspective as they provide means for investigating temporal genetic change in the species concerned. We suggest that a centrally based record keeping system for documenting what is available at various institutions needs to be developed. This would constitute an important step towards the realization of genetic monitoring programs that would not require excessive amounts of funding.
Scientific and logistic issues in genetic monitoring
Conservation genetic monitoring to meet CBD goals imply the identification of conditions where natural gene pools are severely depleted or compromised. As illustrated in Fig. 1, this includes situations where (i) genetically distinct local populations or population segments have been or are at risk of being lost, (ii) inbreeding and genetic drift is considerably elevated, (iii) alien genes become established, or (iv) the genetic composition is altered due to direct human induced selection. Monitoring programs need to have the capacity to identify these types of changes and separate them, with a reasonable degree of reliability, from naturally occurring temporal genetic shifts that are due to genetic drift, gene flow, and natural selection.
This leads to a number of both logistic and scientific issues that need to be addressed. For example, what levels of inflow of alien genes do we need to detect? What reductions of effective population size (increased drift/inbreeding) are acceptable? How strong human induced selection is acceptable? What sample sizes, in terms of individuals, sampling locations, and gene loci, are required to obtain adequate statistical power to detect these changes? How can collection of material for conservation genetic monitoring be organized? What institutions should organize such collections? It is beyond the scope of this presentation to go into detail, but we stress the urgent need for further exploration of all these topics.
As of present, our knowledge of microevolutionary processes and short term temporal variability patterns of natural populations is limited (Hendry and Kinnison 1999; Laikre et al. 2005). This is exemplified here by the very few studies (4%) of Swedish species involving more than a single sampling occasion. General knowledge on rates of temporal genetic change is needed for testing strategies for sampling design, data collection, analytical procedures for interpreting observed temporal genetic changes, and identifying situations of loss of gene level biodiversity. We propose that research focusing on these issues should be prioritized.
The type of genetic markers used for studying genetic variation of Swedish species vary considerably. Most frequently used markers are allozymes, microsatellites, and mitochondrial DNA (mtDNA), but the number and types of loci used vary both among and within species. The interpretation of variation patterns based on different types of markers are frequently not straight forward (Ryman et al. 2006). Further, the efficiency of detecting various levels and types of substructuring varies due to differences in statistical power and may be more or less pronounced in particular situations (e.g. Larsson et al. 2007).
The population genetic structure of particular species is shaped by microevolutionary processes affected by landscape and environmental features (Manel et al. 2003). Yet, our understanding of how ecological processes affect genetic variability and the relationship between landscape variables and spatio-temporal population genetic structure is largely unknown (Holderegger and Wagner 2006; Storfer et al. 2006). Conservation genetic monitoring aims at detecting changes of genetic composition and levels of variability over time, and also to provide insights into the reasons for observed changes. It will be important to link temporal genetic processes to anthropogenically induced change, ecological processes, and environmental factors. Increased research into these areas is urgently needed.
References
Aho T, Laikre L (eds) (2004) Genetic diversity in commercially exploited fish species. Report from Expert-seminar at Hólar College, Iceland, October 6–8, 2004. Norden, Report from the Nordic Council of Ministers
Allendorf FW, Luikart G (2007) Conservation and the genetics of populations. Blackwell, Malden
Allendorf FW, Ryman N (2002) The role of genetics in population viability analysis. In: Beissinger SR, McCullough DR (eds) Population viability analysis. University of Chicago Press, Chicago, pp 50–85
Andersson A-C, Andersson S, Lönn M (2007) Genetic variation in natural populations of animals and plants in Sweden—a review of case studies, theory and some methods. The Swedish Environmental Protection Agency, Report 5712 (in Swedish, English summary)
Berg LM, Nilsson T (1997) Introduktion av främmande arter i svensk landmiljö. Omfattning och konsekvenser. Naturvårdsverket Rapport 4658 (Introduction of alien species in terrestrial environments in Sweden. Extent and consequences. Swedish Environmental Protection Agency (in Swedish))
Carlsson J, Nilsson J (2000) Population genetic structure of brown trout (Salmo trutta L.) within a northern boreal forest stream. Hereditas 132:173–181
Ceplitis A (2001) Genetic and environmental factors affecting reproductive variation in Allium vineale. J Evol Biol 14:721–730
Dannewitz J, Maes GE, Johansson L, Wickström H, Volckaert FAM, Järvi T (2005) Panmixia in the European eel: a matter of time. Proc R Soc B 272:1129–1137
Ellegren H, Savolainen P, Rosen B (1996) The genetical history of an isolated population of the endangered grey wolf Canis lupus: a study of nuclear and mitochondrial polymorphisms. Phil Trans R Soc Lond Ser B 351:1661–1669
Ernande B, Dieckmann U, Heino M (2003) Adaptive change in harvested populations: plasticity and evolution of age and size at maturation. Proc R Soc Lond Ser B 271:415–423
Estonian Ministry of Environment (1997) Eesti keskkonnaseire 1996 (Estonian environmental monitoring (In Estonian, English translation)). Eesti Vabariigi Keskkonnaministeerium, Info-ja Tehnokeskus, Estonian Ministry of Environment, Environmetal Information Centre, Tallinna Taamatutrükikoda
Flagstad Ø, Walker CW, Vilà C, Sundqvist AK, Fernholm B, Hufthammer AK, Wiig O, Koyola I, Ellegren H (2003) Two centuries of the Scandinavian wolf population: patterns of genetic variability and migration during an era of dramatic decline. Mol Ecol 12:869–880
Florin AB, Höglund J (2007) Absence of population structure of turbot (Psetta maxima) in the Baltic Sea. Mol Ecol 16:115–126
Franklin IR, Frankham R (1998) How large must populations be to retain evolutionary potential? Anim Conserv 1:69–70
Grift RE, Rijnsdorp AD, Barot S, Heino M, Dieckmann U (2003) Fisheries-induced trends in reaction norms for maturation in North Sea plaice. Mar Ecol Prog Ser 257:247–257
Gyllensten U (1985) Temporal allozyme frequency changes in density fluctuating populations of willow grouse (Lagopus lagopus L.). Evolution 39:115–121
Hansson B, Bensch S, Hasselquist D, Lillandt BG, Wennerberg L, von Schantz T (2000) Increase of genetic variation over time in a recently founded population of great reed warblers (Acrocephalus arundinaceus) revealed by microsatellites and DNA fingerprinting. Mol Ecol 9:1529–1538
Harris RB, Wall WA, Allendorf FW (2002) Genetic consequences of hunting: what do we know and what should we do? Wildl Soc Bull 30:634–643
Hendry AP, Kinnison MT 1999 The pace of modern life: measuring rates of contemporary microevolution. Evolution 53:1637–1653
Holdegger R, Wagner HH (2006) A brief guide to landscape genetics. Landsc Ecol 21:793–796
Jorde PE, Ryman N 1996 Demographic genetics of brown trout (Salmo trutta) and estimation of effective population size from temporal change of allele frequencies. Genetics 143:1369–1381
Jørgensen HBH, Hansen MM, Loeschcke V (2005a) Spring-spawning herring (Clupea harengus L.) in the southwestern Baltic Sea: do they form genetically distinct spawning waves? ICES J Mar Sci 62:1065–1075
Jørgensen HBH, Hansen MM, Bekkevold D, Ruzzante DE, Loeschcke V (2005b) Marine landscapes and population genetic structure of herring (Clupea harengus L.) in the Baltic Sea. Mol Ecol 14:3219–3234
Laikre L (ed) (1999) Conservation genetic management of brown trout (Salmo trutta) in Europe. Report by the Concerted action on identification, management and exploitation of genetic resources in the brown trout (Salmo trutta) “TROUTCONCERT”; EU FAIR CT97-3882; ISBN 87-987732-0-8 1999
Laikre L, Ryman N (1996) Effects on intraspecific biodiversity from harvesting and enhancing natural populations. Ambio 25:504–509
Laikre L, Ryman N (1997) Monitoring biological diversity at the gene level. Suggestions for actions and future research. Swedish Environmental Protection Agency, Report 4824 (in Swedish)
Laikre L, Palmé A (2005) Release of alien populations in Sweden. Swedish Environmental Protection Agency, Report 5475 (in Swedish, English summary)
Laikre L, Jorde PE, Ryman N (1998) Temporal change of mitochondrial DNA haplotype frequencies and female effective size in a brown trout (Salmo trutta) population. Evolution 52:910–915
Laikre L, Järvi T, Johansson L, Palm S, Rubin JF, Glimsäter CE, Landergren P, Ryman N (2002) Spatial and temporal population structure of sea trout at the Island of Gotland, Sweden, delineated from mitochondrial DNA. J Fish Biol 60:49–71
Laikre L, Palm S, Ryman N (2005) Genetic population structure of fishes—implications for coastal zone management. Ambio 34:111–119
Laikre L, Palmé A, Josefsson M, Utter FM, Ryman N (2006) Spread of alien populations in Sweden. Ambio 35:255–261
Laikre L, Palmé A, Larsson LC, Lindberg E (2007) Spread of genetically alien populations in Sweden. What do we know? What do we do? What are the effects? Swedish Environmental Protection Agency, Report 5683 (in Swedish, English summary)
Larsson LC, Laikre L, Palm S, André C, Carvalho G, Ryman N 2007 Concordance of allozyme and microsatellite differentiation in a marine fish, but evidence for selection at a microsatellite locus. Mol Ecol 16:1135–1147
Liberg O, Andrén H, Pedersen H, Sand H, Sejberg D, Wabakken P, Åkesson M, Bensch S (2005) Severe inbreeding depression in a wild wolf (Canis lupus) population. Biol Lett 1:17–20
Lynch M, Lande R (1998) The critical effective size for a genetically secure population. Anim Conserv 1:70–72
Manel S, Schwartz MK, Luikart G, Taberlet P (2003) Landscape genetics: combining landscape ecology and population genetics. TREE 18:189–197
Martins SS, Sanderson JG, Silva-Júnior JS (2007) Monitoring mammals in the Caxiuanã National Forest, Brazil—First results from the tropical ecology, assessment and monitoring (TEAM) program. Biodiv Conserv 16:857–870
Mestres F, Balañà J, Segarra C, Prevosti A, Serra L (1994) O chromosome inversion polymorphism in Northern and Atlantic Europe and its implications in the American colonization by Drosophila subobscura. Zeitschrift für Zoologische Systematik und Evolutionsforschung 32:108–116
Nielsen EE, Nielsen PH, Meldrup D, Hansen MM (2004) Genetic population structure of turbot (Scophthalmus maximus L.) supports the presence of multiple hybrid zones for marine fishes in the transition zone between the Baltic Sea and the North Sea. Mol Ecol 13:585–595
Niklasson M, Tomiuk J, Parker ED (2004) Maintenance of clonal diversity in Dipsa bifurcata (Fallen, 1810) (Diptera: Lonchopteridae). I. Fluctuating seasonal selection moulds long-term coexistence. Heredity 93:62–71
Nyman L, Ring O (1989) Effects of hatchery environment on three polymorphic loci in arctic char Salvelinus alpinus species complex. Nord J Freshw Res 65:34–43
Nyström V, Angerbjörn A, Dalén L (2006) Genetic consequences of a demographic bottleneck in the Scandinavian arctic fox. Oikos 114:84–94
Norwegian Agency for Nature Conservation (Direktoratet for naturforvaltning) (1998. Plan for overvåkning av biologisk mangfold (Plan for monitoring biological diversity (in Norwegian)). DN-rapport 1998–1
Olsen EM, Heino M, Lilly GR, Morgan MJ, Brattey J, Ernande B, Dieckmann U (2004) Maturation trends indicative of rapid evolution preceded the collapse of northern cod. Nature 428:932–935
Olsson J, Aho T, Florin A-B, Vainikka A, Bekkevold D, Dannewitz J, Hindar K, Koljonen M, Laikre L, Magnussen E, Pålsson S (2007) Genetic research on commercially exploited fish species in Nordic countries. Report, Nordic Council of Ministers
Palm S, Laikre L, Jorde PE, Ryman N (2003a) Effective population size and temporal genetic change in stream resident brown trout (Salmo trutta, L.). Conserv Genet 4:249–264
Palm S, Dannewitz J, Järvi T, Petersson E, Prestegaard T, Ryman N (2003b) Lack of molecular genetic divergence between sea-ranched and wild sea trout (Salmo trutta). Mol Ecol 12:2057–2071
Parducci L, Suyama Y, Lascoux M, Bennett KD (2005) Ancient DNA from pollen: a genetic record of population history in Scots pine. Mol Ecol 14:2873–2882
Paulsen GM (ed) (1997) Overvåkning av biologisk mangfold i åtte naturtyper. Forslag fra åtte arbeidsgrupper (Monitoring of biological diversity in eight habitat types. Suggestions from eight working groups (in Norwegian)). Utredning for DN 1997-7. Norwegian Agency for Nature Conservation, Direktoratet for naturforvaltning
Poulsen NA, Nielsen EE, Schierup MH, Loeschcke V, Grønkjær P (2006) Long-term stability and effective population size in North Sea and Baltic Sea cod (Gadus morhua). Mol Ecol 15:321–331
Rasmussen G, Geertz-Hansen P (1998) Stocking of fish in Denmark. In: Cowx IG (ed) Stocking and introduction of fish. Fishing News Books, Oxford, pp 14–21
Ruzzante DE, Mariani S, Bekkevold D, André C, Mosegaard H, Clausen LAW, Dahlgren TG, Hutchinson WF, Hatfield EMC, Torstensen E, Brigham J, Simmonds EJ, Laikre L, Larsson LC, Stet RJM, Ryman N, Carvalho GR (2006) Biocomplexity in a highly migratory pelagic marine fish, Atlantic herring. Proc R Soc B Sci 273:1459–1464
Ryman N (1983) Patterns of distribution of biochemical genetic variation in salmonids: differences between species. Aquaculture 33:1–21
Ryman N, Ståhl G (1980) Genetic changes in hatchery stocks of brown trout Salmo trutta. Can J Fish Aquat Sci 37:82–87
Ryman N, Laikre L (1991) Effects of supportive breeding on the genetically effective population size. Conserv Biol 5:325–329
Ryman N, Baccus R, Reuterwall C, Smith MH (1981) Effective population size, generation interval, and potential loss of genetic variability in game species under different hunting regimes. Oikos 36:257–266
Ryman N, Palm S, André C, Carvalho GR, Dahlgren TG, Jorde PE, Laikre L, Larsson LC, Palmé A, Ruzzante DE (2006) Power for detecting genetic divergence: differences between statistical methods and marker loci. Mol Ecol 15:231–245
Ryman N, Utter F, Allendorf FW, Busack C, Shaklee J (1993) Genetic concerns about hatchery populations—a comment on the conclusions of Nyman and Ring. J Fish Biol 42:471–480
Ryman N, Utter F, Laikre L (1995) Protection of intraspecific biodiversity of exploited fishes. Rev Fish Biol Fish 5:417–446
Schwartz MK, Luikart G, Waples RS (2007) Genetic monitoring as a promising tool for conservation and management. TREE 22:25–33
Seddon JM, Ellegren H (2004) A temporal analysis shows major histocompatibility complex loci in the Scandinavian wolf population are consistent with neutral evolution. Proc R Soc B Sci 271:2283–2291
Seddon JM, Parker HG, Ostrander EA, Ellegren H (2005) SNPs in ecological and conservation studies: a test in the Scandinavian wolf population. Mol Ecol 14:503–511
Seddon JM, Sundqvist AK, Björnerfeldt S, Ellegren H (2006) Genetic identification of immigrants to the Scandinavian wolf population. Conserv Genet 7:225–230
Storfer A, Murphy MA, Evans JS, Goldberg CS, Robinson S, Spear SF, Dezzani R, Delmelle E, Vierling L, Waits LP (2006) Putting the “landscape” in the landscape genetics. Heredity 98:128–142
Sundqvist AK, Ellegren H, Olivier M, Vilà C (2001) Y chromosome haplotyping in Scandinavian wolves (Canis lupus) based on microsatellite markers. Mol Ecol 10:1959–1966
Swedish Environmental Protection Agency (1999) Nytt program för miljöövervakning (A new programme for environmental monitoring (in Swedish, English summary)) Report 4980, Stockholm, Sweden
Swedish National Board of Fisheries (2007) Fiskbestånd och miljö i hav och sötvatten. Resurs-och miljööversikt 2007, Fiskeriverket (Fish stocks and sea and fresh water environments. Resource and environment survey 2007, Swedish National Board of Fisheries (in Swedish))
Tallmon DA, Bellemain E, Swenson JE, Taberlet P (2004) Genetic monitoring of Scandinavian brown bear effective population size and immigration. J Wildl Manage 68:960–965
UNEP (United Nations Environment Programme) (1995) Global Biodiversity Assessment Cambridge University Press, Cambridge
Usher M (2002) Edited reports from the four biodiversity conservation priority workshops In Scottish Biodiversity Committee—Action Plan and Science Group—Biodiversity Conservation Priority Workshop, Edinburgh, 1 August 2002, www.biodiversityscotland.gov.uk
Wang J, Ryman N (2001) Genetic effects of multiple generations of supportive breeding. Conserv Biol 15:1619–1631
Vilà C, Sundqvist AK, Flagstad Ø, Seddon J, Björnerfeldt S, Kojola I, Casulli A, Sand H, Wabakken P, Ellegren H (2003) Rescue of a severely bottlenecked wolf (Canis lupus) population by a single immigrant. Proc R Soc B Sci 270:91–97
WWF (2001) The Status of Wild Atlantic Salmon: a river by river assessment. WWF, World Wildlife Fund, 184 pp, (http://worldwildlife.org/salmon/pubs.cfm)
Acknowledgements
We thank Michael Schwartz for valuable comments on the manuscript. Financial contribution from the Swedish Environmental Protection Agency (LL), the Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning (LL), and the Swedish Research Council (LL, NR), is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Laikre, L., Larsson, L.C., Palmé, A. et al. Potentials for monitoring gene level biodiversity: using Sweden as an example. Biodivers Conserv 17, 893–910 (2008). https://doi.org/10.1007/s10531-008-9335-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-008-9335-2