
Abstract
Nonnative fish species introduced into new areas, especially when they develop into large populations, pose a threat to wild fauna. We used stable isotope analysis to quantify the ecological effects of introduced Bighead Carp (Aristichthys nobilis) stocking in Ulungur Lake on the basal food source composition, isotopic niche width and niche overlap of four dominant fish species including Bighead Carp, Pond Smelt (Hypomesus olidus), Common Bream (Abramis brama) and Common Carp (Cyprinus Carpio). Following Bighead Carp stocking, proportional contributions of phytoplankton to fishes significantly decreased for the other fishes (P < 0.05), and the contributions of particulate organic matter to the fishes significantly increased (P < 0.05) except for Common Bream. In addition, the total isotopic niche width of fishes increased from 12.01 to 13.06 and isotopic niche overlap between fishes increased from 22.75% to 42.50% following stocking; this change was particularly apparent in the large changes in the isotopic niche overlap between Common Bream and Bighead Carp (from 67 to 95%) and between Common Bream and Pond Smelt (from 65 to 100%), followed by that between Bighead Carp and Pond Smelt (from 0 to 90%). Furthermore, the obvious increase in isotopic niche overlap may be the reason for the reduced growth observed in Bighead Carp and in Pond Smelt. These findings highlighted that Bighead Carp stocking changed the availability of basal food sources for other sympatric fishes and increased the potential for interspecific competition among fishes. Overall, our study suggested the introduced Bighead Carp stocking had undesirable effects on fish community in Ulungur Lake.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The pace of global environmental change has increased substantially due to new environmental pressures that result from human activity (Pimm et al. 1995), and human-mediated introductions of nonnative species are among the most widespread and damaging of these pressures (Hooper 2005). The introduction of nonnative species can have strong ecological impacts on native natural resources, with the affected factors ranging from genes to ecosystems. In aquatic ecosystems, stocking is commonly used to enhance recreational catches, create new fisheries and improve water quality. However, stocking is a highly contentious and debated management practice for fisheries due to its high investment cost and limited scientific evaluation (Hunt and Jones 2017). Critical evaluation of stocking practices has only gained momentum in the last two decades (Leber 2002), but studies of documented effects focus on recapture and production rates following fish stocking (Holmlund and Hammer 2004). Much more attention needs to be paid to the ecological response following introduced fish stocking (e.g., availability in food resources, competitive interactions) (Johnston et al. 2018; Lorenzen 2014; Hunt and Jones 2017).
Haupt and Phelps (2016) and Wang et al. (2018) demonstrated that introducing the stocking of planktivorous Bighead Carp (Aristichthys nobilis) and Silver Carp (Hypophthalmichthys molitrix) affected the availability of phytoplankton to other filter-feeding fishes, which increased the potential for interspecific competition among the fishes and could lead to declines in the condition and abundance of native planktivores in the Missouri River, USA. Similarly, the introduced fish species Prussian Carp (Carassius gibelio) in the Karamenderes River, northwest Turkey, had extensive isotopic niche overlap with the native fish species and outcompeted coexisting species in resource utilization, which posed a serious threat to native fish fauna (Özdilek et al. 2019). However, without knowledge of the trophic niche size and diet composition of wild fish prior to introduced fish stocking, is impossible for researchers to determine if the current niche size and diet have changed after fish introduction. Thus, a before-after study design aiming to quantify the relative strength of the impacts of introduced stocking fish on other sympatric species is important.
The population niche provides a critical basis for understanding competitive interactions between species and populations (Wang et al. 2019). Traditional gut content analysis is insufficient to measure niche metrics, which require quantitative examination of the diets of many individuals in a population, and preferably over an extended time period (Syväranta and Jones 2008). However, stable isotope analysis offers time-integrated trophic information on consumers, which is an integrated measure of niche width, as the selection of tissues with appropriate turnover rates in consumers can be linked qualitatively to the variance among individuals in isotope space (Bearhop et al. 2004). Advances in stable isotope analysis have made quantifying isotopic niche metrics (including isotopic niche width and niche overlap) among and within populations increasingly practicable (Layman et al. 2007; Jackson et al. 2011; Swanson et al. 2015) and have been widely applied in the field of competitive interactions in recent decades (Kingsbury et al. 2020; Ying et al. 2020; Andrades et al. 2019; Hill et al. 2015).
Bighead Carp are large-bodied planktivorous cyprinids that comprise much of the Carp aquaculture production in China (Ke et al. 2008). The stocking of Bighead Carp for improving water quality or increasing economic yield is used in many waters, such as Taihu, Chaohu, Dianchi, Qiandaohu and Ulungur Lakes (Ke et al. 2008; Liu et al. 2007). Compared with other aquatic ecosystems, Bighead Carp is a nonnative species in Ulungur Lake that was introduced from the Yangtze Basin (Tang et al. 2012). Ulungur Lake, one of the most important fishing grounds in the Chinese inland area and one of China’s most northwestern desert lakes, exhibited different fish community patterns before and after Bighead Carp stocking (Tang et al. 2012). Before stocking, the yields of Pond Smelt (Hypomesus olidus), Common Bream (Abramis brama) and Northern Pike (Esox lucius) accounted for 40%, 30% and 18% of the total yield, respectively (Tang et al. 2012). Since the spring of 2012, an average of 20 million Bighead Carp fries and 172 t of Bighead Carp fingerlings and yearlings were stocked into Ulungur Lake annually, while captured 56.36 t Bighead Carp annually (Fig. S1), which makes Bighead Carp with the second-largest yield and makes Pond Smelt, Bighead Carp, Common Bream and Common Carp (Cyprinus Carpio) the dominant fish species (together accounting for 89.14% of the total catch). Thus, we expected to quantify the potential competition among these four dominant fishes with potential diet overlap following fish stocking, inferred through stable isotope analysis. Based on the increase in fish populations with similar prey groups (e.g., Pond Smelt and Bighead Carp), the working hypothesis was that the total isotopic niche width of fishes may increase to achieve coexistence and isotopic niche overlap between fishes may enhance to respond to the Bighead Carp stocking. The information discussed in this study will contribute important insights regarding the potential effects of introduced Bighead Carp stocking on the cooccurring fish and management of Ulungur Lake.
Materials and methods
Study site description
Ulungur Lake is located in the northern Xinjiang Uygur Autonomous Region in northwestern China (46°52′ ~ 47°28′N, 87°00′ ~ 87°34′E) and is the second largest lake (917.0 km2) in Xinjiang (Zhou et al. 2013). It is the downstream lake connecting Jelekol Lake and the Irtysh River, and its annual mean precipitation and evaporation capacities are 116.5 mm and 1844.4 mm, respectively (Zhou et al. 2013). For a complete description of the physical and biological characteristics of this station, see Ji et al. (2018). Ulungur Lake is surrounded by the Gobi Desert, where vegetation is exiguous. Scirpus validus, Phragmites australis, Potamogeton crispus, and Myriophyllum spicatum are the dominant aquatic plants, and the lake has experienced gradual losses of biodiversity and abundance, particularly in submerged plants. The sediments in the lake present a mixture of organic compounds, and the major sources of these organic compounds are inputs from anthropogenic activities (Yao et al. 2013). Yao et al. (2013) indicated that Ulungur Lake has been undergoing deterioration of the water quality and a decrease from the previously high level of biodiversity.
Sampling preparation and analysis
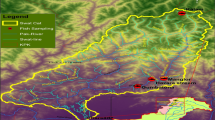
Samples for stable isotope analysis were collected in November of 2012 and 2016 from Ulungur Lake (Fig. 1). The samples collected in 2012 represented the starting point of fish stocking, while those collected in 2016 represented the after-stocking period. For the two studied years, basal sources include phytoplankton, particulate organic matter (POM), submerged plants, emergent plants and sedimentary organic matter (SOM), and the dominant fish species (Bighead Carp, Pond Smelt, Common Bream and Common Carp) were sampled and analyzed.

Sampling sites in 2012 (sites 2, 5 and 6) and 2016 (sites 1–6) in Ulungur Lake
The leaves of emergent plants (S. validus and P. australis) were collected by hand and washed with distilled water to remove detritus. Submerged plants (P. pectinatus, P. crispus, M. spicatum and Chara braunii) were obtained with grabs (50 cm × 38 cm) and subsequently washed with distilled water. Phytoplankton samples were obtained and collected from hauls of a 64-µm phytoplankton net and filtered through precalcined (450 °C for 6 h) Whatman GF/F filters. POM was collected from mixed water (from the upper layer, middle layer and lower layer of the lake) that was passed through a 64-µm phytoplankton net onto precalcined GF/F filters under a moderate vacuum (10 mbar) until clogging occurred (Andrades et al. 2019). The top 0–1 cm of the surface sediment was collected using a Peterson grab and was immediately placed into a sterile plastic bag (Beaudoin et al. 2001). Fishes were sampled using an anchored net (mesh size < 10 mm), a trawling net (mesh size 40–80 mm) and a three-floor hang net. Tissue samples were taken from the white muscle of fishes just between the dorsal fin and the lateral line after removal of all skin and scales (Pinnegar and Polunin 1999; Zanden et al. 1999). The muscle samples were immediately wrapped in acid-cleaned plastic bags, placed in an ice box, and transported to the laboratory (Li et al. 2015). All samples (fish muscle, aquatic plants, POM, SOM, phytoplankton) were dried in an oven at 60 °C for 48 h to a constant weight and then ground to homogenous powder using a glass mortar and pestle (Andrades et al. 2019). Prior to δ13C measurement, the POM, SOM and phytoplankton samples were treated with 1 N HCl to eliminate carbonates using a drop-by-drop technique. Each treated sample was then oven dried again at 60 °C and ground again to a fine powder. All samples used for the δ15N analyses did not undergo any acidification treatment (Zheng et al. 2018; Andrades et al. 2019).
The availability of food sources (phytoplankton, crustacean zooplankton and macrozoobenthos) before stocking referred to the literature (Liu et al. 2009; Yang et al. 2011; Zhao 2010), while the sample collections of them after stocking see Materials and Methods Section in Supporting Information.
Stable isotope analysis
Stable isotope analysis of all samples was carried out at the Key Laboratory of Crop Ecophysiology and Farming System in the Middle Reaches of the Yangtze River, Ministry of Agriculture, using a Vario ISOTOPE cube elemental analyzer coupled to an Isoprime-100 isotope mass spectrometer. The stable isotopic ratios were expressed in the conventional δ notation as parts per mil (‰), according to the following equation:
where R is 13C or 15N and X is 13C/12C or 15N/14N. Atmospheric nitrogen (for δ15N) and Pee Dee Belemnite (PDB) (for δ13C) were used as standards (Andrades et al. 2019). The analytical precision was within 0.1‰ and 0.2‰ for the carbon and nitrogen isotope measurements, respectively.
All stable isotope data analyses were performed in R 3.3.3 to estimate the contributions of multiple sources to each consumer species, isotopic niche width of each fish species and isotopic niche overlap among the four fish species. The relative contribution of primary production resources assimilated by aquatic consumers was estimated using a Bayesian stable isotope mixing model (Bond and Diamond 2011) in the SIAR package. The isotopic niche widths of the fish species were calculated from the δ13C and δ15N data as the standard ellipse area corrected for the sample size (SEAc), which is less sensitive to outliers (Jackson et al. 2011), using Stable Isotope Bayesian Ellipses in the SIBER package. A probabilistic method developed by Swanson et al. (2015) was used to test whether the isotopic niche overlap was not equivalently weighted among species in the nicheROVER package. The method measured a given 95% (or user-defined α) probability isotopic niche size and provided directional estimates of the pairwise isotopic niche overlap in multivariate space. The isotopic niche overlap of species A onto species B was defined as the probability that an individual from species A is found in the niche of B and vice versa (Swanson et al. 2015).
Statistical analysis
Statistical analyses were conducted with the 19.0 version of SPSS software (Chicago, IL, USA). Kruskal–Wallis tests were used to test for differences in the δ13C and δ15N signatures among the basal food sources and fishes. The Mann–Whitney U test was executed to compare the difference between the isotopic signatures of the same basal source or the same fish species between 2012 and 2016. All tests maintained a comparison wise type I error rate of 0.05.
Results
Stable isotope composition
Phytoplankton, POM, submerged plants, emergent plants and SOM were the primary carbon sources in Ulungur Lake between 2012 and 2016. The δ13C and δ15N values of these sources are presented in Fig. 2 and Table S1. In 2012, the basal food sources were distinguishable based on their δ13C values, ranging from − 26.70 ± 3.87‰ to − 19.78 ± 0.09‰. The δ15N values of the basal food sources varied from 0.06 ± 0.01‰ to 5.30 ± 0.76‰. In 2016, the basal food sources spanned a δ13C range of − 27.39 ± 1.63 to − 18.80 ± 2.27‰ and a δ15N range of 2.94 ± 2.16 to 5.10 ± 0.60‰. Great differences were observed among the δ13C values of the basal food sources in 2012 (Kruskal–Wallis analysis, P < 0.01) and 2016 (P < 0.01), which allowed for the calculation of the proportional contribution to consumers of each basal food resource using the Bayesian mixing model. Among the five potential food sources (phytoplankton, POM, submerged plants, emergent plants and SOM), there were significant differences in the δ13C values of submerged plants (Mann–Whitney U test, P < 0.05) between 2012 and 2016 and highly significant differences in the δ13C values of SOM (Mann–Whitney U test, P < 0.01) between the two years. Likewise, significant differences were observed among the δ15N values of the basal sources in 2012 (Kruskal–Wallis analysis, P < 0.01) and 2016 (P < 0.01). There were highly significant differences in the δ15N values of POM and SOM between 2012 and 2016 (Mann–Whitney U test; POM: P < 0.01; SOM: P < 0.01).

δ13C—δ15N diagrams of the basal food sources and the fish species in Ulungur Lake between 2012 (a) and 2016 (b). Basal food sources are denoted by red squares; fish are denoted by blue diamonds. Each square or diamond represents one species. The species codes are as follows: Phy, phytoplankton: POM, particulate organic matter; Eme, emergent plants; Sub, submerged plants; SOM, sediment organic matter; BHC, Bighead Carp; CMC, Common Carp; PDS, Pond Smelt; and CMB, Common Bream
The δ13C values of the fishes ranged from − 24.32 ± 1.27‰ to − 21.63 ± 2.33‰ and the δ15N values varied from 7.36 ± 0.39‰ to 9.53 ± 0.09‰ in 2012. In 2016, the four fish species spanned a δ13C signature range of − 24.24 ± 1.87‰ to − 21.53 ± 1.98‰ and a δ15N signature range of 8.02 ± 0.70‰ to 9.88 ± 0.73‰ (Fig. 2, Table S1). Great differences were observed among the four fish species in the δ15N values in 2012 (Kruskal–Wallis analysis, P < 0.01), and there were also significant differences in the δ13C values (Kruskal–Wallis analysis, P < 0.01) and δ15N values (P < 0.01) among the four fish species in 2016. Significant differences in the δ13C values of Bighead Carp were observed between 2012 and 2016 (Mann–Whitney U test, P < 0.05), and significant differences in the δ13C values of Common Bream were also observed (Mann–Whitney U test, P < 0.01). Furthermore, the δ15N of all four fish species changed significantly between the two years (Mann–Whitney U test; Bighead Carp: P < 0.01; Common Bream: P < 0.01; Common Carp: P < 0.05; Pond Smelt: P < 0.05).
Changes in the contributions of the basal food sources to the four fish species
Wide variation in the contributions of the basal food sources to the four fish species were observed between 2012 and 2016 (Fig. 3). Specifically, compared to 2012, the contributions of phytoplankton in 2016 significantly decreased from 35 to 21% for Common Carp, Pond Smelt and Common Bream (Mann–Whitney U test, P < 0.05), while that for Bighead Carp increased slightly from 30 to 32%. Additionally, the contributions of POM significantly increased from 11 to 18% for Common Carp, Pond Smelt and Bighead Carp (Mann–Whitney U test, P < 0.05), while that for Common Bream decreased from 11 to 6%. The estimated contributions of SOM significantly increased from 13 to 38% from 2012 to 2016 (Mann–Whitney U test, P < 0.05). Furthermore, the contribution of emergent plants to the four fishes decreased from 20 to 16%, while that of submerged plants significantly decreased from 22 to 7% from 2012 to 2016 (Mann–Whitney U test, P < 0.05; Fig. 3).

Bayesian mixing model solutions for the proportions of the basal food sources for the four fish species in Ulungur Lake between 2012 and 2016. Boxes indicate the 50%, 75% and 95% Bayesian credibility intervals. The species codes are consistent with those in Fig. 2
Changes in isotopic niche width and niche overlap
All species except for Common Bream showed consistent upward trends in isotopic niche width (SEAc). Compared to those in 2012, the isotopic niche width of Bighead Carp, Common Carp, and Pond Smelt increased from 2.14 to 2.15, 1.47 to 4.15, and 0.07 to 0.44, respectively, in 2016, while the isotopic niche width of Common Bream decreased from 8.33 to 6.32 (Fig. 4).

Density plot showing the credibility intervals of the standard ellipse areas (SEAs) in 2012 and 2016. Black circles are the mode SEAs, and boxes indicate the 50%, 75% and 95% credible intervals from wider to thinner. The green Xs are the sample-size-corrected SEAs (SEAc). The species codes are consistent with those in Fig. 2
The average isotopic niche overlap among the four fish species was as high as 22.75% in 2012, which was considerably lower than the average overlap among the fish species in 2016 (42.5%). Compared with that in 2012, the isotopic niche overlap among the fish species increased to different extents in 2016, except for the overlap between Common Bream and Common Carp, which decreased from 74 to 68%. In 2012, the isotopic niche overlap among the four fish species varied from 0 to 74%. Among the four fish species, the average isotopic niche overlap between Common Bream and the other fishes was highest (mean = 68.67%). The highest overlap values were all involved with Common Bream; the overlap values between Common Bream and Common Carp, Bighead Carp, and Pond Smelt were 74%, 67% and 65%, respectively. However, there was almost no overlap between Pond Smelt and other fishes. In 2016, the isotopic niche overlap among the four fish species varied from 0 to 100%. Interestingly, the mean isotopic niche overlap between Common Bream and other fishes was also highest in this year (mean = 87.67%), and, similarly to the trend seen in 2012, the mean overlaps between Pond Smelt and the other fishes were the lowest (mean = 11.50%). Specifically, the highest overlaps in 2016 were observed between Common Bream and Pond Smelt (100%) and between Common Bream and Bighead Carp (95%). In addition, the overlap between Bighead Carp and Pond Smelt was as high as 90% in 2016. However, in this year, the overlap between Pond Smelt and Common Carp was lowest (Fig. 5a, b).

Probabilistic isotopic niche overlap (%) for a standard eclipse isotopic niche space of 95% in Ulungur Lake in 2012 (a) and 2016 (b). The means and 95% intervals are displayed in blue. The species codes are consistent with those in Fig. 2
Discussion
This study is apparently the first to quantify the changes in diet composition, isotopic niche width and isotopic niche overlap in the dominant fish species in Ulungur Lake using stable isotope analysis following the introduction of fish stocking. Our findings suggest that the changed patterns in fish assemblages driven by the Bighead Carp stocking influenced food sources availability and the distribution of the nutritional contributions. Furthermore, our results support our initial hypotheses that the total isotopic niche width of fishes and isotopic niche overlap between fish species increased following the Bighead Carp stocking. The increased isotopic niche overlap indicated the potential for increased interspecific competition following stocking in Ulungur Lake. These findings will provide deeper insight into the potential effects of Bighead Carp stocking on cooccurring fish and offer essential input for a sustainable management strategy in Ulungur Lake.
In general, shifts in the relative contributions of the food sources of fish species respond to changes in the availability of basal food sources (Correa and Winemiller 2016). Conducting fish stocking can lead to changes in species composition and abundance and in the use of native resources in aquatic systems (Eby et al. 2006). Horn (2003) demonstrated that stocking of planktivorous fishes caused enormous predation pressure on the zooplankton group, with obvious increases in the concentrations of nano and picophytoplankton, which were too small to be effectively preyed upon by other fishes. In Ulungur Lake in 2016, compared with the period before-stocking Bighead Carp, an increase of 10.41 times in the cell density and an increase of only 1.11 times in the biomass of phytoplankton in 2016 indicated that phytoplankton communities tended to be miniaturized (Liu et al. 2009, Table S2) following stocking. The miniaturization of phytoplankton could make it difficult for the affected phytoplankton to be eaten as prey by other fishes; this may partially explain the significantly decreased nutritional contribution of phytoplankton to the fish species (except for Bighead Carp). Similar cases have been studied in Taihu Lake in China (Ke et al. 2008) and in Mississippi River in the USA (Wang et al. 2018). In Taihu Lake, after the stocking of Bighead Carp and Silver Carp, the main diets of both species shifted from zooplankton to phytoplankton, and the presence of the stocked species ultimately increased the concentration of small phytoplankton individuals (Ke et al. 2008). In Mississippi River, Wang et al. (2018) reported that the phytoplankton community was miniaturized after the introduction of Bighead Carp and Silver Carp and the miniaturized phytoplankton could not be effectively preyed upon by other planktivorous fishes. Furthermore, the turbidity in Ulungur Lake was higher after the stocking of Bighead Carp than before (Ji et al. 2018), which may reflect the increased availability of POM and partly explain the obviously increased nutritional contributions of POM to the fishes. Similarly, Yu et al. (2016) demonstrated that the concentration of POM increased significantly after the stocking of new species and that the nutritional contributions of POM were improved for sympatric species. Thus, our study supports other data and indicates that planktivorous fish stocking could change the community structure and availability of basal food sources, especially phytoplankton and POM, which is a crucial factor affecting the variety of the nutritional contributions of basal food sources to fish species.
Consumers with overlapping diets cope with changes in shared food sources and regulate their diet width to achieve coexistence during periods of food scarcity in aquatic ecosystems (Correa and Winemiller 2016; Leray et al. 2019). When preferred basal sources become limited, consumer species will broaden their diets (i.e., niche expansion) to include alternative foods to meet their dietary requirements (Perry and Pianka 1997; Svanback and Bolnick 2005). Such a change in foraging behavior in a given species is, however, likely to affect the interactions of that species with other consumer species via increased niche overlap and potential for competition (Correa and Winemiller 2016; Pianka 1974, 1976). In Ulungur Lake, compared with the period before-stocking Bighead Carp, the phytoplankton communities were obviously diminished following stocking (Liu et al. 2009, Table S2); Accordingly, the average density of crustacean zooplankton increased from 2.41 ind./L before stocking to 28.47 ind./L after stocking and biomass decreased from 0.066 to 0.046 mg/L (Yang et al. 2011; Table S3). In addition, the macroinvertebrates density and biomass decreased significantly after stocking (Mann–Whitney U test; density: P < 0.05; biomass: P < 0.05, Zhao 2010, Fig. S2), in which the average value of macroinvertebrates density decreased from 1048.13 to 233.64 ind./m2 and biomass decreased from 10.77 to 4.59 g/m2 (Fig. S2). On the other hand, the population density of nonnative Bighead Carp could increase following continue stocking practices (Fig. S1). In the present study, compared the results before stocking, the total niche width increased from 12.01 to 13.06 following stocking, in a pattern consistent with the niche variation hypothesis (Van Valen 1965; Bolnick et al. 2010). Furthermore, the average isotopic niche overlap between the fishes increased from 22.75% in 2012 to 42.5% in 2016 (Fig. 5). The above results indicated that after Bighead Carp stocking, the fish species changed the isotopic niche width with the potential for interspecific competition enhanced under the reduced availability in food sources and the increased abundance of nonnative species. Similarly, Taylor et al. (2018) measured isotopic niche width and overlap from stable isotope analysis to examine potential competitive interactions arising from Eastern King Prawns (Melicertus plebejus) stocking, and indicated that the isotopic niche width of sympatric fish species varied in response to Eastern King Prawns stocking and competitive interactions may occurr, especially between Eastern King Prawns and Palaemonid Prawn (Macrobrachium intermedium) with the greatest isotopic niche overlap. Özdilek et al. (2019) also found that the introduction of Prussian Carp into the Karamenderes River in Turkey led to extensive isotopic niche overlaps between Prussian Carp and other fish species and may pose a serious threat to other fish fauna. Overall, our findings and previous studies indicate that stocking of introduced fish could result in the changes in isotopic niche width of fishes and enhance the potential for interspecific competition between fishes.
Intense interspecific competition among fish species may result in reverse effects on different fishes, especially for fishes at competitive disadvantages (Britton et al. 2018; Munday et al. 2001; Irons et al. 2007). One of the primary considerations is that outcompeting consumer species will limit the growth of weakly competitive fish species when sources are limiting, particularly at higher levels of fish species density (Kindinger 2018). In Ulungur Lake, the availability of macrozoobenthos significantly decreased after stocking (Fig. S2). In addition, the population density of Common Bream gradually increased and the yield of Common Bream reached 158.7 t/year followed by the yields of Pond Smelt and Bighead Carp, and the catch of Bighead Carp, Pond Smelt and Common Bream accounted for 86.57% of the total catch. Despite the high abundance of Common Bream, Hu (2014) illustrated that Common Bream was miniaturized and the number of individuals with body length less than 15 cm accounted for more than 66% of the total sampled individuals. Furthermore, the diminished Common Bream population feeds mainly on zooplankton (length < 15 cm) (Biró 1991). The limited food resources and the population characteristics of Common Bream may partly explain the highest isotopic niche overlap between Common Bream and the other fish species (a mean of 68.67% in 2012 and 87.67% in 2016), particularly between Common Bream and Bighead Carp (increased from 67 to 95%) and between Common Bream and Pond Smelt (increased from 65 to 100%). Moreover, the overlap changed obviously between Bighead Carp and Pond Smelt (from 0 to 90%) (Fig. 5). These results suggested the increase in potential for interspecific competition among the zooplanktivorous fishes (Pond Smelt, Bighead Carp, Common Bream with body length less than 15 cm) and the highest isotopic niche overlap was observed among them indicating the largest potential for competition following stocking (Morrison et al. 2021, Fig. 5). Furthermore, the diminished crustacean zooplankton community responded to the shifts in predation pressure and high isotopic niche overlap among Common Bream, Bighead Carp and Pond Smelt (Yang et al. 2011, Table S3). Thus, the obvious increase isotopic niche overlap among the zooplanktivorous fishes under the limiting food sources and high abundance of fish species in Ulungur Lake was potentially responsible for the slower growth of Bighead Carp (Zheng et al. 2002; Gao et al. 2012; Zhu et al. 2013, Table 1) and the smaller Pond Smelt following stocking in Ulungur Lake (Table 2). Similarly, Morrison et al. (2021) qualified the impacts of Zebra Mussels (Dreissena polymorpha) on isotopic niche size and niche overlap among fish species in a mesotrophic lake, in which the isotopic niche overlap between species increased significantly (1.2-fold) post Zebra Mussels and that could ultimately result in reduced abundance of species less able to utilize littoral energy sources. Additionally, in the Mississippi River, after the introduction and rapid growth of Bighead Carp and Silver Carp there was high isotopic overlap between the introduced species and the native planktivores, and potential competition between them may lead to declines in the body conditions of the native fishes, including the Bigmouth Buffalo (Ictiobus cyprinellus) and the Gizzard Shad (Dorosoma cepedianum) (Irons et al. 2007; Wang et al. 2018). Therefore, our study indicates that under the limiting food sources and high abundance of fish species, the high isotopic niche overlap between fishes may be the reason for the reduced growth observed in Bighead Carp and in Pond Smelt following stocking in Ulungur Lake.
Conclusions
In the present study, changes in the basal food source composition, isotopic niche width and niche overlap of four dominant fish species (Bighead Carp, Pond Smelt, Common Bream and Common Carp) were investigated following the introduction of Bighead Carp stocking in Ulungur Lake. The work by 2012 and 2016 demonstrated that the stocking of Bighead Carp affected the variety of basal resources consumed by other dominant species in the lake. In addition, the total isotopic niche width of fishes and isotopic niche overlap between fish species enhanced to respond to the Bighead Carp stocking. The high isotopic niche overlap among zooplanktivorous fishes could be potentially responsible for the reduced growth observed in Bighead Carp and in Pond Smelt following stocking in Ulungur Lake. Thus, with the implementation of an annual release plan, long-term data will be critical to understanding how introduced fish stocking can influence fish population dynamics (e.g., growth and abundance) and what ecological problems may occur over time in Ulungur Lake.
Availability of data and material
All data generated or analysed during this study are included in this published article [and its supplementary information files].
References
Andrades R, Jackson AL, Macieira RM et al (2019) Niche-related processes in island intertidal communities inferred from stable isotopes data. Ecol Indic 104:648–658
Bearhop S, Adams CE, Waldron S et al (2004) Determining trophic niche width: a novel approach using stable isotope analysis. J Anim Ecol 73:1007–1012
Beaudoin CP, Prepas EE, Tonn WM et al (2001) A stable carbon and nitrogen isotope study of lake food webs in Canada’s Boreal Plain. Freshwater Biol 46:5–77
Biró P, Sadek SE, Paulovits G (1991) The food of Bream (Abramis brama L.) in two basins of Lake Balaton of different trophic status. Hydrobiologia 209:51–58
Bolnick DI, Ingram T, Stutz WE et al (2010) Ecological release from interspecific competition leads to decoupled changes in population and individual niche width. Proc Biol Sci 277:1789–1797
Bond AL, Diamond AW (2011) Recent Bayesian stable-isotope mixing models are highly sensitive to variation in discrimination factors. Ecol Appl 21:1017–1023
Britton RJ, Ruiznavarro A, Verreycken H et al (2018) Trophic consequences of introduced species: Comparative impacts of increased interspecific versus intraspecific competitive interactions. Funct Ecol 32:486–495
Correa SB, Winemiller KO (2016) Niche partitioning among frugivorous fishes in response to fluctuating resources in the Amazonian floodplain forest. Ecology 95:210–224
Eby LA, Roach WJ, Crowder LB et al (2006) Effects of stocking-up freshwater food webs. Trends Ecol Evol 21:576–584
Gao ZJ, Yu HX, Ma CX et al (2012) The preliminary study on the diversity and community structure of the fish in Xiquanyan reservoir. Chi J Fish 25:19–24 (in Chinese with an English abstract)
Haupt KJ, Phelps QE (2016) Mesohabitat associations in the Mississippi River Basin: a long-term study on the catch rates and physical habitat associations of juvenile Silver Carp and two native planktivores. Aquat Invasions 11: 93–99
Hill JM, Jones RW, Hill MP (2015) Comparisons of isotopic niche widths of some invasive and indigenous fauna in a South African river. Freshwater Biol 60:893–902
Holmlund CM, Hammer M (2004) Effects of fish stocking on ecosystem services: an overview and case study using the Stockholm Archipelago. Environ Manage 33:799–820
Hooper DU, Chapin FS, Ewel JJ et al (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monogr 75:3–35
Horn W (2003) Long-term development of the crustacean plankton in the Saidenbach Reservoir (Germany)-changes, causes, consequences. Hydrobiologia 504:185–192
Hu SD (2014) Age, growth and reproduction of the common bream Abramis brama in Ulungur Lake, Xinjiang, China. Dissertation, Huazhong Agricultural University
Hunt TL, Jones P (2017) Informing the great fish stocking debate: an Australian case study. Rev Fish Sci Aquac 26:275–308
Irons KS, Sass GG, McClelland MA et al (2007) Reduced condition factor of two native fish species coincident with invasion of non-native Asian Carps in the Illinois River, USA Is this evidence for competition and reduced fitness? J Fish Biol 71:258–273
Jackson AL, Inger R, Parnell AC et al (2011) Comparing isotopic niche widths among and within communities: SIBER-Stable Isotope Bayesian Ellipses in R. J Anim Ecol 80:595–602
Ji FF, Shen JZ, Ma XF et al (2018) Changes of the water quality and the underlying causes analysis in the Ulungur Lake. J Hydrogeol 39:61–66 (in Chinese with an English abstract)
Johnston FD, Allen MS, Beardmore B et al (2018) How ecological processes shape the outcomes of stock enhancement and harvest regulations in recreational fisheries. Ecol Appl 28:2033–2054
Ke Z, Xie P, Guo L (2008) In situ study on effect of food competition on diet shifts and growth of Silver and Bighead Carps in large biomanipulation fish pens in Meiliang Bay, Lake Taihu. J Appl Ichthyol 24:263–268
Kindinger TL (2018) Invasive predator tips the balance of symmetrical competition between native coral-reef fishes. Ecology 99:792–800
Kingsbury KM, Gillanders BM, Booth DJ et al (2020) Trophic niche segregation allows range-extending coral reef fishes to co-exist with temperate species under climate change. Global Change Biol 26:721–733
Layman CA, Quattrochi JP, Peyer CM et al (2007) Niche width collapse in a resilient top predator following ecosystem fragmentation. Ecol Lett 10:937–944
Leber KM (2002) Advances in marine stock enhancement: shifting emphasis to theory and countability. In: Stickney RR, Mcvey JP (eds) Responsible Marine Aquaculture. CABI Publishing, New York, pp 79–90
Leray M, Alldredge AL, Yang JY et al (2019) Dietary partitioning promotes the coexistence of planktivorous species on coral reefs. Mol Ecol 28:2694–2710
Li J, Zhou Q, Yuan G et al (2015) Mercury bioaccumulation in the food web of Three Gorges Reservoir (China): tempo-spatial patterns and effect of reservoir management. Sci Total Environ 527:203–210
Liu QG, Chen Y, Li JL et al (2007) The food web structure and ecosystem properties of a filter-feeding Carps dominated deep reservoir ecosystem. Ecol Model 203:279–289
Liu Y, Shen JZ, Li H et al (2009) Phytoplankton community structure in Lake Ulungur in Xinjiang Uygur Autonomous Region. J Lake Sci 21:855–864 (in Chinese with an English abstract)
Lorenzen K (2014) Understanding and managing enhancements: Why fisheries scientists should care. J Fish Biol 85:1807–1829
Morrison AL, Thelen MA, Howe SE et al (2021) Impacts of zebra mussels (Dreissena polymorpha) on isotopic niche size and niche overlap among fish species in a mesotrophic lake. Biol Invasions 1–18
Munday PL, Jones GP, Caley MJ (2001) Interspecific competition and coexistence in a Guild of Coral-Dwelling Fishes. Ecology 82:2177–2189
Özdilek ŞY, Partal N, Jones RI (2019) An invasive species, Carassius gibelio, alters the native fish community through trophic niche competition. Aquat Sci 81:29
Perry G, Pianka E (1997) Animal foraging: past, present and future. Trends Ecol Evol 12:360–364
Pianka ER (1974) Niche overlap and diffuse competition. P Natl Acad Sci 71:2141–2145
Pianka ER (1976) Competition and niche theory. In: May RM (ed) Theoretical ecology: principles and applications. Blackwell Scientific, Oxford, pp 114–141
Pimm SL, Russell GJ, Gittleman JL et al (1995) The future of biodiversity. Science 269:347–350
Pinnegar JK, Polunin NVC (1999) Differential fractionation of δ13C and δ15N among fish tissues: implications for the study of trophic interactions. Funct Ecol 13:225–231
Svanback R, Bolnick DI (2005) Intraspecific competition affects the strength of individual specialization: an optimal diet theory method. Evol Ecol Res 7:993–1012
Swanson HK, Lysy M, Power M et al (2015) A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecology 96:318–324
Syväranta J, Jones RI (2008) Changes in feeding niche widths of perch and roach following biomanipulation, revealed by stable isotope analysis. Freshwater Biol 53:425–434
Tang FJ, Brown A, Keerjiang A (2012) Fish community successions in Lake Ulungur: a case of fish invasions in fragile oasis. Russ J Biol Invasions 3:76–80
Taylor MD, Lowry MB, Becker A (2018) Evaluating potential competitive interactions following stocking through trophic niche breadth. Mar Freshwater Res 69:1614–1625
Van Valen L (1965) Morphological variation and width of ecological niche. Am Nat 99:377–390
Wang JZ, Chapman D, Xu J et al (2018) Isotope niche dimension and trophic overlap between bigheaded Carps and native filter-feeding fish in the lower Missouri River, USA. PLoS ONE 13:e0197584
Wang YY, Huan Z, Wei CY (2019) Trophic niche width and overlap of three benthic living fish species in Poyang Lake: a stable isotope approach. Wetlands 39:17–23
Yang LL, Zhou XY, Liu QG et al (2011) Seasonal succession of crustacean zooplankton in relation to the major environmental factors in Lake Ulungur, Xinjiang. Acta Ecol Sin 31:4468–4476
Yao X, Lu J, Liu Z (2013) Distribution of sterols and the sources of pollution in surface sediments of Ulungur lake, Xinjiang. Water Sci Technol 67:2342–2349
Ying R, Cao Y, Yin F et al (2020) Trophic structure and functional diversity reveal pelagic-benthic coupling dynamic in the coastal ecosystem of Daya Bay, China. Ecol Indic 113:106241
Yu HB, Gao QF, Dong SL et al (2016) Effect of co-culture of Chinese shrimp (Fenneropenaeus chinensis) and sea cucumber (Apostichopus japonicus Selenka) on pond environment. J Ocean Univ 15:905–910
Zanden MJV, Casselman JM, Rasmussen JB (1999) Stable isotope evidence for the food web consequences of species invasions in lakes. Nature 401:464–467
Zhao YJ (2010) Studies on community structure of benthic macroinvertebrates in Ulungur Lake. Dissertation, Huazhong Agricultural University
Zheng SP, Peng BC, Xin J (2002) Study on the biology of the Silver Carp and Big-head in the Talagan Reservoir. Inner Mongolia Agr Sci Tech 2:18–20 (in Chinese with an English abstract)
Zheng YY, Niu JG, Zhou Q et al (2018) Effects of resource availability and hydrological regime on autochthonous and allochthonous carbon in the food web of a large cross-border river (China). Sci Total Environ 612:501–512
Zhou XY, Hu ZJ, Liu QG et al (2013) Feeding ecology of the non-indigenous fish Hypomesus nipponensis in Lake Ulungur, China: insight into the relationship between its introduction and the collapse of the native Eurasian perch population. Marine and freshwater research. 64(6):549–557
Zhu FY, Liu JS, Li ZJ et al (2013) The growth performance of Hypophthalmichthys molitrix and Aristichthys mobilis in Lake Kuilei and their fisheries utilization. Freshw Fish 43:32–35 (in Chinese with an English abstract)
Acknowledgements
This study was supported by Special Funds for the Foundation Work of Science and Technology, Ministry of Science and Technology of the People’s Republic of China (2012FY112700). We are grateful that the Key Laboratory of Crop Ecophysiology and Farming System for the Middle Reach of the Yangtze River (Huazhong Agricultural University) provided technical assistance on sample analysis. We thank Lei Xiong, Peng Xie and Xiuhua Sun for field sampling and laboratory analyses.
Author information
Authors and Affiliations
Contributions
Conceptualization: FJ and JS. Formal analysis and investigation: FJ, XM, LQ and ZK. Writing—original draft preparation: FJ. Writing—review and editing: JS. Funding acquisition: JS. Supervision: JS and XM.
Corresponding author
Ethics declarations
Conflict of interest statement
The authors declare no actual or potential competing financial interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Ji, F., Ma, X., Qiu, L. et al. Quantifying the effects of introduced Bighead Carp (Cyprinidae; Aristichthys nobilis) stocking on dominant fish species in the Ulungur Lake, China. Biol Invasions 24, 1253–1265 (2022). https://doi.org/10.1007/s10530-021-02715-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-021-02715-0