
Abstract
A key question in invasion biology is why some regions have more alien species than others. Here, we provide a general framework to answer this. We model alien species richness as a function of the number of species introduced (colonization pressure) and the probability that each species establishes, which is a function of propagule pressure (the number of introduction events and the number of individuals per event) and the probability that a founding individual leaves a surviving lineage (lineage survival probability). With this model we show that: (a) alien species richness is most sensitive to variation in colonization pressure; (b) heterogeneity in lineage survival probability lowers the probability of population establishment, implying alien richness should be lower in more spatially or temporally variable environments; (c) heterogeneity in lineage survival probability leads to higher alien richness when a given propagule pressure is divided into more introduction events, each involving fewer individuals; and (d) we cannot quantify how specific components, such as lineage survival probability, influence alien species richness without data on other components and knowledge of how these covary. Overall, the model provides novel insights into the factors influencing alien species richness, and shows why we expect anthropogenic effects to be critical to this.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
One of the primary ways that humans are driving global environmental change is by introducing species to areas beyond their natural biogeographic ranges. Such species—here termed aliens—are now a feature of virtually every biological community worldwide (McGeoch et al. 2010). With the increasing volume and globalisation of human trade and transport networks, the numbers of successfully established, or naturalised, alien species continues to grow at ever-increasing rates (Seebens et al. 2017; Sikes et al. 2018). Central to understanding the problem of invasive alien species is explaining why the richness and composition of alien species varies so dramatically in different parts of the world (Elton 1958). In just a few centuries, the processes that underpin the arrival, establishment and spread of alien species have generated broad-scale patterns in alien species richness that both parallel and contrast with patterns in native species richness (Case 1996; Sax et al. 2002; Dyer et al. 2017). Yet, despite their potential for revealing the processes driving invasions, documenting and understanding patterns in alien species richness remains a challenge.
On the one hand, the richness (and other features) of alien assemblages is likely to be influenced by the biotic and abiotic features of the recipient regions (Shea and Chesson 2002) on the basis that alien species may respond in the same way as native species to geographic drivers known to structure assemblages in predictable ways (e.g., higher species richness in tropical relative to temperate regions). However, natural ecological processes are not the only drivers of community assembly in invasion biology—the number and composition of alien species in different regions is strongly influenced by human activities. In particular, biological invasion is a multi-stage process: a species must successfully pass through a series of sequential stages to become an invasive alien species (Blackburn et al. 2011). This sequential nature of the invasion process, means that the characteristics of an alien assemblage at any given stage depends on the characteristics of the assemblage in previous stages, along with those processes that act to determine which species successfully transition between any two stages. These filters include factors that depend on human actions, such as decisions about which species to transport purposefully to new regions, or the extent of trade and transportation between regions that entrains accidental introductions (Hulme 2009). Consequently, to understand the characteristics of alien assemblages, one has to understand the role of human actions, particularly in determining which species were transported and introduced to where. Since those species that establish and become invasive are a subset of those introduced (Williamson 1996; Blackburn et al. 2011), this historical human context is likely to leave a strong imprint on the composition (Williamson 1996; Cassey et al. 2004a; Diez et al. 2009) and richness (Dyer et al. 2017) of alien species assemblages.
While an increasing number of studies are using geographic data to study how alien species richness varies around the globe (Van Kleunen et al. 2015a, b; Dawson et al. 2017; Dyer et al. 2017), most studies do so in the absence of a clear conceptual framework to guide interpretation. Analyses of alien species richness need to be set in the context of the invasion pathway, specifically accounting for the number of alien species introduced to an area (colonization pressure; Lockwood et al. 2009). Here, we outline a model that describes the processes that determine the number of alien species successfully established in a region, given knowledge of colonization pressure. First, we derive an equation for alien species richness in terms of colonization pressure and the probability that each introduced species establishes a self-sustaining population, with this probability described in terms of the number of introduction events, the number of individuals in each event (which together comprise propagule pressure; Lockwood et al. 2005) and the likelihood a single individual will leave a surviving lineage and hence found a population (which we term ‘lineage survival probability’). Colonization pressure and propagule pressure depend on human actions, while lineage survival probability depends on how favourable the introduction location is to the arriving species, which determines whether individuals can survive and reproduce, and ultimately establish a self-sustaining population. Second, we use simulations to examine how variation in these parameters is expected to influence alien species richness. Finally, we discuss the implications of our findings in relation to the causes of variation in alien species richness.
A general model for alien species richness
Lonsdale (1999) proposed that the richness (R) of alien species in a region, R = p̂S; where S is the number of species introduced (colonization pressure) and p̂ is the proportion of introduced species that survive and establish a self-sustaining population (note that we have altered Lonsdale’s original notation to avoid confusion below). Rewriting Lonsdale’s (1999) equation in terms of the probability, Pi, that the ith introduced species establishes:
The probability, Pi, that an introduced species will establish is well known to depend on propagule pressure: the number of attempts to introduce a species and the number of individuals associated with each attempt (Lockwood et al. 2005; Hayes and Barry 2008; Blackburn et al. 2009; Simberloff 2009; Duncan et al. 2014; Cassey et al. 2018). For the ith species, a model for establishment probability as a function of propagule pressure is:
where Ii is the number of introduction events for species i (where an event is the introduction of some number of individuals), Nij is the number of individuals of species i introduced at event j, and pij is lineage survival probability: the probability that an individual of species i introduced at event j leaves a surviving lineage and thus founds a population (Leung et al. 2004; Duncan et al. 2014). The total propagule pressure for species i is the sum over Ii introduction events of Nij individuals per event, or the product of Ii and Nij, when Nij is constant across events. Combining Eqs. 1 and 2 gives:
Equation 3 identifies three components underlying variation in alien species richness. First, alien species richness should be higher when colonization pressure is greater. All else being equal, introducing more species (higher S) should result in more species establishing. Second, alien species richness should be higher when average propagule pressure is greater (the number of introduction events per species, Ii, and the number of individuals introduced per event, Nij) because more founding populations or greater founding population size means a higher chance that at least one founding population succeeds. Finally, alien species richness should be higher when lineage survival probability is higher (higher pij), which implies that conditions at the introduction site (e.g., biotic and abiotic variables) are more suited to the persistence (i.e., survival, growth, reproduction) of the species, given its ecological, life history and other demographic characteristics.
We expect two of the three components underlying variation in alien species richness to be strongly associated with human actions in the introduction process: colonization pressure (S) and propagule pressure (I and N; from now on we drop the subscripts for convenience). For intentionally introduced species, these components will be determined largely by human behaviour, regarding which species to introduce and how many individuals to transport and release. For accidentally introduced species, these components will be a feature of the pathways by which introductions occur, which will determine the number and type of species, and the number of individuals, accidentally entrained and transported to new locations (Wilson et al. 2009). The third component, lineage survival probability, p, reflects the interaction between the traits of a species and features of the environment to which it is introduced. Individuals introduced to environments that are favourable for population growth, and that are not subject to large fluctuations in population size, due to demographic, genetic or environmental stochasticity, are more likely to leave surviving lineages and thus establish, than are individuals introduced to less favourable environments (Dennis 2002; Lande et al. 2003; Schreiber and Lloyd-Smith 2009). While it is sometimes possible to obtain data on colonization pressure and propagule pressure (e.g., from knowledge of the numbers and types of species introduced to regions), it is much more difficult to determine lineage survival probability for species prior to introduction because it is hard to determine how species will behave in a new environment, the composition of individuals (i.e., demographic, phenotypic, genotypic), and the precise conditions they will face on arrival.
Nevertheless, we can identify key components that will underlie differences in lineage survival probability, which helps clarify some important invasion concepts (Richardson and Pyšek 2006). Specifically, p could vary predictably among species due to differences in traits that increase or decrease lineage survival probability, and hence establishment success, in a range of environments (Kolar and Lodge 2001; Hayes and Barry 2008). This would result in some species being classed as more invasive than others because they possess traits that result in individuals having a higher probability of leaving a surviving lineage at a greater range of locations, increasing the likelihood that founding populations will establish. Similarly, p could vary predictably among locations such that some locations are more likely than others to be invaded by many species because those locations have favourable environments where founding individuals of many species have a high chance of leaving a surviving lineage (Stohlgren et al. 1997; Shea and Chesson 2002). These definitions highlight that the concepts of species invasiveness and location invasibility reflect predictable differences among species and locations in lineage survival probability, which is the relevant measure of performance independent of colonization and propagule pressures (Lonsdale 1999; Duncan et al. 2003; Catford et al. 2012).
We expect p to vary depending on species and location, and between introduction events of the same species to the same location if conditions vary over time (Norris et al. 2002). To explore how this heterogeneity in p might influence alien richness, we extend Eq. 3 to allow for variation in p among introduction events. We can model heterogeneity in p using a beta distribution, which constrains p to values between 0 and 1, and provides a flexible distribution able to capture plausible ways in which p might vary, including bimodal distributions where some species or events have high and others low lineage survival probability. When heterogeneity in p is modelled using a beta distribution, the equation for expected alien richness becomes (see Duncan et al. 2014):
where B is the beta function, and α and β are the parameters of a beta distribution describing the distribution of p values among introduction events. Across all introduction events, individuals have a mean probability of leaving a surviving lineage, \(\bar{p}\) = α/(α + β) with variance \(\sigma_{p}\) = αβ/[(α + β)2(α + β + 1)]. While it may be difficult to estimate lineage survival probability for a given introduction event (Duncan 2016), Eq. 4 provides a model that allows the influence of variation in lineage survival probability on alien species richness to be explored.
Sensitivity of alien richness to different drivers
Understanding how and why alien species richness varies among locations or through time involves understanding the contributions of the components we have identified in driving richness patterns. As a starting point, we therefore analysed the sensitivity of alien species richness to variation in each of the three components (colonization pressure, S, propagule pressure, I and N, and lineage survival probability, p). Given that there will be variation in lineage survival probability among events, this term has two components: a mean (\(\bar{p}\)) and variance (\(\sigma_{p}\)) (see above). Using Eq. 4, we examined the sensitivity of alien richness to varying four parameters in turn: S, I, N and \(\bar{p}\) given four values for \(\sigma_{p}^{2}\) (0, 1 × 10−4, 0.001 and 0.005), noting that when \(\sigma_{p}^{2}\) = 0, Eq. 4 reduces to Eq. 3.
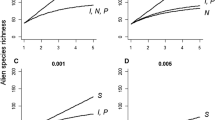
The results of the sensitivity analysis are shown in Fig. 1. All else being equal, greater heterogeneity in p (i.e., higher \(\sigma_{p}^{2}\) for the same \(\bar{p}\)) reduces the probability of establishment overall, and thus results in lower alien species richness R (compare Fig. 1a with no heterogeneity to the other panels in Fig. 1). This is because \(\bar{p}\) typically takes small values so that any increase in heterogeneity resulting in some larger p values leads to a skewed distribution with disproportionally more small p values to offset this, lowering the probability of establishment for most species.

Sensitivity analysis showing how alien species richness changes with a proportional increase in the parameters of Eqs. 3 and 4: colonization pressure S, propagule pressure I and N, and mean lineage survival probability \(\bar{p}\). We specified initial values of number of species introduced to a location S = 100, the number of introduction events per species I = 10, the number of individuals introduced per event N = 10, and lineage survival probability \(\bar{p}\) = 0.005. With these parameters, Eq. 3 gives the expected alien species richness R = 39. We then measured the effect on R of increasing each parameter, in turn, by a proportion ranging from 1 to 5 (i.e., increasing each parameter by up to 5 times the initial value), while keeping the other parameters fixed at the initial values. Choosing different initial values results in qualitatively the same outcomes. a is the sensitivity analysis assuming no heterogeneity in lineage survival probability (Eq. 3; \(\sigma_{p}^{2}\) = 0), while b–d assume increasing levels of heterogeneity in \(\bar{p}\) (Eq. 4 with \(\sigma_{p}^{2}\) = 0.0001, 0.001 and 0.005, respectively; with these values shown above each plot)
Regardless of the value of \(\sigma_{p}^{2}\), a proportional change in the number of introduction events I has the same effect on species richness R, as the same proportional change in the mean probability of lineage survival \(\bar{p}\): all else being equal, doubling I increases alien species richness R by the same amount as doubling \(\bar{p}\). However, when there is heterogeneity in p (\(\sigma_{p}^{2}\) > 0), R is less sensitive to changes in the number of individuals per introduction attempt N than it is to changes in I and \(\bar{p}\), and becomes increasingly less sensitive as \(\sigma_{p}^{2}\) increases. This means that, when there is variation in lineage survival probability among events within species, alien species richness will be greater if the total propagule pressure for each species (I × N) is divided into more introduction events I involving fewer individuals per event N (Fig. 1b-d). When there is no heterogeneity in p, it makes no difference to alien species richness whether individuals are introduced in one large event or several smaller events (Fig. 1a). This sensitivity of outcomes to variation in p may explain why different studies have reached different conclusions regarding the importance of I versus N in controlling establishment success (Hopper and Roush 1993; Haccou and Iwasa 1996; Grevstad 1999; Haccou and Vatutin 2003; Cassey et al. 2014).
Variation in alien species richness R was most sensitive to variation in colonization pressure S. When S is fixed and any of I, N or p increases, R cannot exceed the limit at R = S, when every introduced species succeeds in establishing. This constrains by how much alien species richness can increase as we increase I, N and p, and hence limits the sensitivity of R to variation in these parameters. In contrast, alien species richness is more sensitive to altering colonization pressure: doubling S increases R by a greater proportion than doubling the other parameters because alien species richness increases as a linear (rather than asymptotic) function of the number of species introduced. This implies that differences among locations in alien species richness will be more sensitive to varying colonization pressure than to varying propagule pressure or lineage survival probability, if other things stay constant. Other things, however, are unlikely to remain constant: we consider next how this might affect establishment outcomes.
Covariation between colonization pressure, propagule pressure and lineage survival
In our sensitivity analysis we varied each parameter while maintaining others constant at their initial values. In reality, however, parameters are likely to covary, potentially in predictable ways. For example, with intentional introductions the environment at a location may affect people’s choices about the number and type of species to introduce, and the effort expended in introducing different types of species, causing propagule pressure to be positively correlated with lineage survival probability. Horticulturalists, for example, often select ornamental species on the basis of how well matched these are to the local climate (e.g., using hardiness zones), leading to well-matched species (those likely to have higher p) being planted more often and more extensively (higher I and N) (Maurel et al. 2016). This may also mean that more species are introduced to locations with relatively benign climates compared to those with harsher climates, such that S and p are positively correlated.
In addition to covariation, parameters could also vary predictably through time. Globally, alien species richness is often positively correlated with measures of trade (Meyerson and Mooney 2007; Westphal et al. 2007; Hulme 2009; Sikes et al. 2018). This is expected, because trade is a key conduit for accidental introductions, with greater trade volumes leading to more individuals being accidentally transported to new locations (Wilson et al. 2009). Lockwood et al. (2009) demonstrated that when individuals are randomly sampled from a source community, with an underlying log series abundance distribution, increasing the number of individuals per sample increased both colonization pressure and mean propagule pressure, meaning S, I and N are positively correlated, reinforcing the association between trade and alien species richness (see Fig. 2). Here we use simulations to extend these findings. We show that when few individuals are sampled from the source community, increasing the number of individuals leads to a faster increase in colonization pressure than mean propagule pressure (Fig. 2b). This suggests that increasing numbers of new species arriving (greater colonization pressure), rather than increasing propagule pressure, drives the increase in alien species richness at low trade volumes. At higher trade volumes, however, colonization pressure will asymptote as samples become large enough to include most species. As trade volume increases beyond this point, propagule pressure continues to increase as more individuals of the same species are included in each sample. As a consequence, rather than slowing as colonization pressure asymptotes, alien species richness may continue to increase steadily with increasing trade volume due to propagule pressure taking over from colonization pressure as the driver of establishment (Fig. 2c). How the form of the relationship between colonization and propagule pressure differs for other realistic species-abundance distributions is unknown but, regardless of the particular assumptions, S, I and N are likely to covary in ways that alter the drivers of alien species establishment as trade volumes vary in time and/or space. Modelling the consequences of this covariation for alien species richness is a clear avenue for future work.

Output from a simulation where different numbers of individuals (10, 50, 100, 500, 1000, 2000, 3000, 4000, 5000, 10,000, 25,000, 50,000, 75,000 and 100,000 on the x-axis) were each sampled 1000 times from a community with the species abundance distribution in a (a Poisson-lognormal distribution with 1000 species having (log) mean abundance = 0.5 and standard deviation = 2). This community can be considered the source pool of alien species from which the different numbers of individuals were sampled; b shows the median number of species per sample as a function of the total number of individuals per sample, with the maximum value of 1000 species shown as a dotted line; c shows the median number of individuals per species as a function of the total number of individuals per sample; d shows median alien species richness (calculated using Eq. 3 with p = 0.005) as a function of the total number of individuals per sample
Other features of the introduction process could further result in parameter covariation, or cause parameters to change in predictable ways through time. For intentional introductions, less interest in pursuing later introductions once some alien species have established may cause propagule pressure to decline (I and N are negatively correlated with S). This could explain declines in the annual rate at which some species, particularly birds and mammals, have established in many countries following earlier efforts at acclimatisation (Duncan and Blackburn 2002; Seebens et al. 2017), particularly once the negative impacts of established species became apparent and efforts shifted to preventing new introductions through biosecurity measures (McDowall 1994). Nevertheless, declining establishment rates through time could be due to other processes. One possibility is that the pool of species available for introduction is being exhausted (Levine and D’Antonio 2003), although Fig. 2 shows that even when this occurs, establishment rates can still rise when increasing trade volume continues to increase propagule pressure. Recent analysis further suggests that the pool of potential alien species is increasing for most unintentionally transported taxa due to widening trade connections (Seebens et al. 2018). Another possibility invokes the idea of saturation: as alien species accumulate in a region it becomes increasingly difficult for new species to establish, due to niche pre-emption, which implies that p is negatively correlated with R independent of variation in propagule pressure (Moulton and Pimm 1983; Duncan 1997; Tilman 2004). Identifying which components of alien species richness (colonization pressure, propagule pressure or lineage survival probability) are driving variation in establishment rates across time and space is clearly challenging because it requires separating out the effects of covarying components.
Implications of the model for alien species richness
Lonsdale (1999) highlighted that the number of alien species in a region is a function of both the number of species introduced and the likelihood that a species establishes a self-sustaining population. Establishment probability is in turn a function of propagule pressure, allowing us to extend Lonsdale’s model and identify the three critical components underlying variation in alien species richness: colonization pressure, propagule pressure and lineage survival probability. Our model (Eqs. 3, 4) has at least two important implications for the study of alien species richness.
First, as we indicated above, it is not possible to quantify how specific components, such as lineage survival probability, influence alien species richness without quantifying the other components, particularly colonization pressure (Williamson 1996; Lonsdale 1999). Our sensitivity analysis (Fig. 1) implies that variation in colonization pressure is most likely a key component of variation in alien richness, with the number of established species increasing as a linear function of the number introduced, in contrast to other parameters that vary asymptotically. The logic of this difference is straightforward: for a given number of introduced species, increasing the number of introduced individuals per species (I and N) or the likelihood that any one individual leaves a surviving lineage (p) can increase establishment success, but can only ever increase alien species richness up to the limit set by colonization pressure (S). Introducing more species increases this limit, and hence alien species richness is more sensitive to changes in S than I, N and p. The influence of propagule pressure on invasion outcomes has already been argued to be a ‘null model for biological invasions’ (Colautti et al. 2006; Cassey et al. 2018), in the sense that it should be included in any attempt to understand and model the invasion process. We propose that colonization pressure should also be considered a null model for biological invasions. Nevertheless, like propagule pressure it is often difficult to quantify colonization pressure: in most regions we do not know how many alien species have been introduced, but subsequently failed to establish. Unfortunately this means that, in the absence of data for S, it is very difficult to unravel what underlies variation in alien species richness because we are missing information on what is probably the most important driver (Diez et al. 2009; García-Díaz et al. 2015). It may be possible to use proxies for S, such as trade volume, albeit advising similar caution as for the use of proxies for propagule pressure because of the potential for spurious correlations (Pigot et al. 2018).
Second, our model shows how colonization pressure, propagule pressure, and lineage survival probability combine to determine alien species richness (Eqs. 3, 4), but we need to understand how these components covary in real situations. Such covariation could lead to reinforcement (e.g., a positive correlation between propagule pressure and lineage survival probability for intentionally introduced species), substitution (e.g., propagule pressure taking over from colonization pressure as trade volume increases) or the cancelling out (e.g., negative correlation between lineage survival probability and colonization pressure due to niche pre-emption) of effects. More generally, our model provides a framework for understanding how specific factors are likely to affect alien richness through their combined influence on colonization pressure, propagule pressure and lineage survival probability. For example, we have argued that both colonization and propagule pressure should be positively correlated with trade volume (Fig. 2) but that alien richness should be more sensitive to changes in colonization pressure. This implies that, for a given trade volume, a country should be at greater risk of alien invasion if it trades with a wide range of partners (high colonization pressure through exposure to many partners, but lower propagule pressure per partner) than if trade is concentrated in fewer partners (higher propagule pressure per partner but lower colonization pressure), an outcome supported by simulation (Fig. 3). More generally, by specifying how factors such as trade interact with the different components of alien species richness, and understanding the sensitivity of these different components (Fig. 1), we should be able to identify the most important interactions underlying variation in alien species richness, generating predictions that can be empirically tested.

Output from a simulation where different numbers of individuals (10, 50, 100, 500, 1000, 2000, 3000, 4000, 5000, 10,000, 25,000, 50,000, 75,000 and 100,000 on the x-axis) were sampled from either 1, 2, 5 or 10 different communities (the different lines). The total number of individuals could represent the number of aliens arriving in a country (a function of trade volume), and each community the source pool of aliens from a different trading partner. Each community is characterised by the species abundance distribution shown in Fig. 2a (a Poisson-lognormal distribution with 1000 species having (log) mean abundance = 0.5 and standard deviation = 2). We modelled arrival such that the total number of arriving individuals (x-axis) was evenly distributed among the number of trading partners (1, 2, 5 or 10). Median alien species richness (calculated using Eq. 3 with p = 0.005 from 1000 simulations) is higher when there are more trading partners (and thus fewer arriving individuals per partner), than when there are fewer partners (and thus more arriving individuals per partner)
Overall, anthropogenic effects are likely to be strong determinants of alien species richness by directly influencing colonization pressure and propagule pressure, and by changing lineage survival probability to favour alien species through environmental modification (e.g., land use or climate change). As we have identified, differences among locations in their inherent invasibility must result from differences in lineage survival probability, independent of variation in colonization pressure or propagule pressure (Catford et al. 2012). Given that colonization and propagule pressure can vary widely, locations would have to show very strong and consistent differences in lineage survival probability for this to be a major driver of variation in alien species richness. A recent global analysis of alien bird species richness supports this finding, with variation in alien richness primarily explained by variation in anthropogenic factors, especially a strong positive effect of colonization pressure, but with some variance explained by a positive effect of native species richness at the recipient location, which could reflect differences in the suitability of locations for both native and alien species persistence (Dyer et al. 2017).
Species traits could also predictably influence lineage survival probability, but perhaps more important are likely to be the relationships between traits and propagule pressure and (indirectly) colonization pressure (Maurel et al. 2016; Peoples and Goforth 2017). In general, widespread, abundant species are more likely to be introduced to new environments, while large-bodied animal species are more likely to be deliberately translocated (Blackburn and Duncan 2001; Jeschke and Strayer 2006; Tingley et al. 2010; Blackburn et al. 2017). Globally, propagule pressure is positively related to geographic range size, annual fecundity, body mass and migratory tendency in birds (Cassey et al. 2004b). The characteristics of species entrained in the invasion pathway are therefore a biased subset relative to the pool of all potential alien species. Thus, the traditional view of the influence of species’ traits on alien species richness—that these affect lineage survival probability by influencing the ability of species to survive and reproduce at new localities—may be less important than the manner in which traits affect how humans perceive and interact with species, directly through deliberate trade or indirectly through the likelihood of accidental translocation (Chapple et al. 2012).
Equation 3 is a general model describing alien richness given knowledge of the species introduced, the number of introduction events and numbers of individuals of each species introduced, and the probability a founding individual in each event would leave a surviving lineage. This generic model can include a wide range of more specific processes that can be modelled by specifying particular relationships among the parameters (e.g., Fig. 3). For example, according to MacArthur and Wilson’s (1963, 1967) equilibrium theory of island biogeography, the rate at which species arrive and establish on an island declines as a function of the number of species already present due to exhaustion of the mainland species pool and competition with residents. Within our framework, this could be modelled by specifying a negative relationship between R and either S or p, or both. Similarly, Allee effects, which can be important in the dynamics of invading populations (Drake 2004; Tobin et al. 2011), would imply that p becomes small whenever N falls below a particular threshold. Nevertheless, while Eq. 3 describes a generic and very flexible model, fitting the model to data on alien species richness with the aim of quantifying the main drivers of variation requires data on some of the key components, particularly colonization and propagule pressure. Data on lineage survival probability are difficult to obtain, if not impossible, despite this being the key parameter to assess differences in invasion risk among species or locations. Consequently, studies typically aim to estimate this parameter and how it varies given data on the remaining components (Sol et al. 2012). Equation 4 may assist with this because it allows for generic modelling of variation via a distribution, rather than having to estimate lineage survival separately for each species or introduction event (for examples, see Duncan et al. 2014; Duncan 2016). Additionally, lineage survival probability can be modelled as a function of general environmental (e.g. seasonality) or species (e.g. life-history) attributes. Nevertheless, we emphasise that without data on at least some components, particularly colonization pressure, it is impossible to quantify the relative importance of other components in driving variation in alien species richness (Lonsdale 1999). This is particularly so where covariates, often used to explain variation in alien species richness (including anthropogenic factors such as trade volume), can influence outcomes via several pathways.
Acknowledging heterogeneity in lineage survival probability p (Eq. 4) also has important implications for understanding variation in alien species richness. First, heterogeneity in lineage survival probability reduces overall establishment probability and lowers alien species richness across the range of parameter values we simulated (Fig. 1). Second, when there is heterogeneity in lineage survival probability among locations, increasing the number of introduction events I has the same effect on alien species richness as an equivalent proportional increase in lineage survival probability, but alien species richness is less sensitive to an equivalent proportional increase in the number of individuals per introduction attempt N (Fig. 1). This implies that when there is heterogeneity in lineage survival probability due, for example, to spatial and/or temporal variation in the suitability of locations for establishment, then establishment probability (and alien species richness) will be higher when total propagule pressure for a species is divided into more introductions of fewer individuals. This occurs because, with environmental heterogeneity, it is better to try many locations, at least one of which may be suitable for establishment with even a few individuals, rather than risking all individuals at one or a few locations that may turn out to be unsuitable (see also Haccou and Iwasa 1996; Grevstad 1999; Haccou and Vatutin 2003).
The above finding implies that if two regions have the same mean lineage survival probability, then the region with greater spatial and temporal heterogeneity in lineage survival probability should have lower alien species richness, suggesting that regions with low environmental heterogeneity might be easier to invade. This negative effect of environmental heterogeneity on alien establishment may appear surprising given that environmental heterogeneity is generally expected to promote diversity by stabilising coexistence. An intriguing possibility is that the higher alien species richness found on islands, relative to continental regions (Dawson et al. 2017), could then be a consequence of lower environmental heterogeneity in suitable habitats, due to their small size and oceanic climate. If so, a further implication would be that, relative to continental regions, introductions of alien species to islands would be less sensitive to how total propagule pressure was distributed across introduction events. Conversely, many introductions of few individuals to continents should increase success, because there are more places or times where introductions would fail on continents, relative to islands, due to low values of p. Other areas with low environmental heterogeneity may include cities, especially in temperate climates, where the urban heat island effect reduces climatic variability, and where anthropogenic food sources (e.g., rubbish dumps, bins, supplemental feeding) may reduce periods of food scarcity. Alien species are particularly prevalent in cities and on islands (van Kleunen et al. 2015a, b; Dawson et al. 2017; Gaertner et al. 2017), although this could equally be due to variation in colonization and propagule pressure (Blackburn et al. 2008; Dawson et al. 2017; Dyer et al. 2017).
In summary, we have outlined a generic model to describe variation in alien species richness, highlighting three critical components—colonization pressure, propagule pressure and lineage survival probability—that underlie this variation. Substantial research effort in invasion biology has been devoted to understanding the characteristics that make some locations more invasible and some species more likely to become alien invaders. Indeed, these were the two key questions posed by the influential SCOPE program on alien species (Drake et al. 1989; Williamson 1996). Our model highlights that location and species-level traits are each components of lineage survival probability p, and that this combines with propagule and colonization pressure to determine establishment success and alien species richness (Fig. 3). While it remains important to identify the traits that underlie variation in lineage survival probability, it is clear that as much effort, if not more, needs to be focussed on understanding determinants of variation in colonization pressure S and propagule pressure (N × I). Traits that determine which species are introduced more often and in larger numbers may matter more in driving the distribution and richness of alien species than traits that increase the likelihood that an individual will leave a surviving lineage in a novel environment. Our model further highlights the need to understand how these components covary, and emphasises that it is impossible to infer the importance of one component without some understanding of the others. Given the importance of colonization pressure as a driver of alien species richness, an extension of our model would be to consider more thoroughly how colonization pressure can vary, particularly in relation to the pool of native species available as potential new alien introductions, the characteristics of the native pool (in terms of species abundances and exposure to humans that might influence the likelihood species will be picked up and transported to new locations), and how anthropogenic factors such as trade influence rates of species arrival (for a start on this, see Leung et al. 2012). We suspect that much of the global variation in alien species richness will be best explained by these factors.
References
Blackburn TM, Duncan RP (2001) Establishment patterns of exotic birds are constrained by non-random patterns in introduction. J Biogeogr 28:927–939. https://doi.org/10.1046/j.1365-2699.2001.00597.x
Blackburn TM, Cassey P, Lockwood JL (2008) The island biogeography of exotic bird species. Glob Ecol Biogeogr 17:246–251. https://doi.org/10.1111/j.1466-8238.2007.00361.x
Blackburn TM, Lockwood JL, Cassey P (2009) Avian invasions: the ecology and evolution of exotic birds. Oxford University Press, Oxford
Blackburn TM, Pyšek P, Bacher S et al (2011) A proposed unified framework for biological invasions. Trends Ecol Evol 26:333–339. https://doi.org/10.1016/j.tree.2011.03.023
Blackburn TM, Scrivens SL, Heinrich S, Cassey P (2017) Patterns of selectivity in introductions of mammal species worldwide. NeoBiota 33:33–51. https://doi.org/10.3897/neobiota.33.10471
Case TJ (1996) Global patterns in the establishment and distribution of exotic birds. Biol Conserv 78:69–96. https://doi.org/10.1016/0006-3207(96)00019-5
Cassey P, Blackburn TM, Jones KE, Lockwood JL (2004a) Mistakes in the analysis of exotic species establishment: source pool designation and correlates of introduction success among parrots (Aves: Psittaciformes) of the world. J Biogeogr 31:277–284. https://doi.org/10.1046/j.0305-0270.2003.00979.x
Cassey P, Blackburn TM, Sol D et al (2004b) Global patterns of introduction effort and establishment success in birds. Proc R Soc Lond Ser B 271:S405–S408. https://doi.org/10.1098/rsbl.2004.0199
Cassey P, Prowse TAA, Blackburn TM (2014) A population model for predicting the successful establishment of introduced bird species. Oecologia 175:417–428. https://doi.org/10.1007/s00442-014-2902-1
Cassey P, Delean S, Lockwood JL et al (2018) Dissecting the null model for biological invasions: a meta-analysis of the propagule pressure effect. PLoS Biol 16:1–15. https://doi.org/10.1371/journal.pbio.2005987
Catford JA, Vesk PA, Richardson DM, Pyšek P (2012) Quantifying levels of biological invasion: towards the objective classification of invaded and invasible ecosystems. Glob Change Biol 18:44–62. https://doi.org/10.1111/j.1365-2486.2011.02549.x
Chapple DG, Simmonds SM, Wong BBM (2012) Can behavioral and personality traits influence the success of unintentional species introductions? Trends Ecol Evol 27:57–62. https://doi.org/10.1016/j.tree.2011.09.010
Colautti RI, Grigorovich IA, MacIsaac HJ (2006) Propagule pressure: a null model for biological invasions. Biol Invasions 8:1023–1037. https://doi.org/10.1007/s10530-005-3735-y
Dawson W, Moser D, Van Kleunen M et al (2017) Global hotspots and correlates of alien species richness across taxonomic groups. Nat Ecol Evol 1:1–7. https://doi.org/10.1038/s41559-017-0186
Dennis B (2002) Allee effects in stochastic populations. Oikos 96:389–401. https://doi.org/10.1034/j.1600-0706.2002.960301.x
Diez JM, Williams PAA, Randall RP et al (2009) Learning from failures: testing broad taxonomic hypotheses about plant naturalization. Ecol Lett 12:1174–1183. https://doi.org/10.1111/j.1461-0248.2009.01376.x
Drake JM (2004) Allee effects and the risk of biological invasion. Risk Anal 24:795–802. https://doi.org/10.1111/j.0272-4332.2004.00479.x
Drake JA, di Castri F, Groves RH et al (1989) Biological invasions. A global perspective. John Wiley & Sons, Chichester
Duncan RP (1997) The role of competition and introduction effort in the success of passeriform birds introduced to New Zealand. Am Nat 149:903–915. https://doi.org/10.1086/286029
Duncan RP (2016) How propagule size and environmental suitability jointly determine establishment success: a test using dung beetle introductions. Biol Invasions 18:985–996. https://doi.org/10.1007/s10530-016-1083-8
Duncan RP, Blackburn TM (2002) Morphological over-dispersion in game birds (Aves: Galliformes) successfully introduced to New Zealand was not caused by interspecific competition. Evol Ecol Res 4:551–561
Duncan RP, Blackburn TM, Sol D (2003) The ecology of bird introductions. Annu Rev Ecol Evol Syst 34:71–98. https://doi.org/10.1146/annurev.ecolsys.34.011802.132353
Duncan RP, Blackburn TM, Rossinelli S, Bacher S (2014) Quantifying invasion risk: the relationship between establishment probability and founding population size. Methods Ecol Evol 5:1255–1263. https://doi.org/10.1111/2041-210X.12288
Dyer EE, Cassey P, Redding DW et al (2017) The global distribution and drivers of alien bird species richness. PLoS Biol 15:1–25. https://doi.org/10.1371/journal.pbio.2000942
Elton C (1958) The ecology of invasions by animals and plants. Methuen, London
Gaertner M, Wilson JRU, Cadotte MW et al (2017) Non-native species in urban environments: patterns, processes, impacts and challenges. Biol Invasions 19:3461–3469. https://doi.org/10.1007/s10530-017-1598-7
García-Díaz P, Ross JV, Ayres C, Cassey P (2015) Understanding the biological invasion risk posed by the global wildlife trade: propagule pressure drives the introduction and establishment of Nearctic turtles. Glob Change Biol 21:1078–1091. https://doi.org/10.1111/gcb.12790
Grevstad FS (1999) Factors influencing the chance of population establishment: implications for release strategies in biocontrol. Ecol Appl 9:1439–1447
Haccou P, Iwasa Y (1996) Establishment probability in fluctuating environments: a branching process model. Theor Popul Biol 50:254–280. https://doi.org/10.1006/tpbi.1996.0031
Haccou P, Vatutin V (2003) Establishment success and extinction risk in autocorrelated environments. Theor Popul Biol 64:303–314. https://doi.org/10.1016/S0040-5809(03)00092-3
Hayes KR, Barry SC (2008) Are there any consistent predictors of invasion success? Biol Invasions 10:483–506
Hopper KR, Roush RT (1993) Mate finding, dispersal, number released, and the success of biological control introductions. Ecol Entomol 18:321–331. https://doi.org/10.1111/j.1365-2311.1993.tb01108.x
Hulme PE (2009) Trade, transport and trouble: managing invasive species pathways in an era of globalization. J Appl Ecol 46:10–18. https://doi.org/10.1111/j.1365-2664.2008.01600.x
Jeschke JM, Strayer DL (2006) Determinants of vertebrate invasion success in Europe and North America. Glob Change Biol 12:1608–1619. https://doi.org/10.1111/j.1365-2486.2006.01213.x
Kolar CS, Lodge DM (2001) Progress in invasion biology: predicting invaders. Trends Ecol Evol 16:199–204. https://doi.org/10.1016/s0169-5347(01)02101-2
Lande R, Engen S, Sæther B-E (2003) Stochastic population dynamics in ecology and conservation. Oxford University Press, Oxford
Leung B, Drake JM, Lodge DM (2004) Predicting invasions: propagule pressure and the gravity of Allee effects. Ecology 85:1651–1660. https://doi.org/10.1890/02-0571
Leung B, Roura-Pascual N, Bacher S et al (2012) TEASIng apart alien species risk assessments: a framework for best practices. Ecol Lett 15:1475–1493. https://doi.org/10.1111/ele.12003
Levine JM, D’Antonio CM (2003) Forecasting biological invasions with increasing international trade. Conserv Biol 17:322–326. https://doi.org/10.1046/j.1523-1739.2003.02038.x
Lockwood JL, Cassey P, Blackburn TM (2005) The role of propagule pressure in explaining species invasions. Trends Ecol Evol 20:223–228
Lockwood JL, Cassey P, Blackburn TM (2009) The more you introduce the more you get: the role of colonization pressure and propagule pressure in invasion ecology. Divers Distrib 15:904–910. https://doi.org/10.1111/j.1472-4642.2009.00594.x
Lonsdale M (1999) Global patterns of plant invasions and the concept of invasibility. Ecology 80:1522–1536
MacArthur RH, Wilson EO (1963) An equilibrium theory of insular zoogeography. Evolution (N Y) 17:373–387. https://doi.org/10.2307/2407089
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
Maurel N, Hanspach J, Kühn I et al (2016) Introduction bias affects relationships between the characteristics of ornamental alien plants and their naturalization success. Glob Ecol Biogeogr. https://doi.org/10.1111/geb.12520
McDowall RM (1994) Gamekeepers for the nation: the story of New Zealand’s acclimatisation societies 1861–1990. Canterbury University Press, Christchurch
McGeoch MA, Butchart SHM, Spear D et al (2010) Global indicators of biological invasion: species numbers, biodiversity impact and policy responses. Divers Distrib 16:95–108. https://doi.org/10.1111/j.1472-4642.2009.00633.x
Meyerson LA, Mooney HA (2007) Invasive alien species in an era of globalization. Front Ecol Environ 5:199–208. https://doi.org/10.1890/1540-9295(2007)5%5b199:IASIAE%5d2.0.CO;2
Moulton MP, Pimm SL (1983) The introduced Hawaiian avifauna: biogeographic evidence for competition. Am Nat 121:669–690. https://doi.org/10.1086/284094
Norris RJ, Memmott J, Lovell DJ (2002) The effect of rainfall on the survivorship and establishment of a biocontrol agent. J Appl Ecol 39:226–234. https://doi.org/10.1046/j.1365-2664.2002.00712.x
Peoples BK, Goforth RR (2017) The indirect role of species-level factors in biological invasions. Glob Ecol Biogeogr 26:524–532. https://doi.org/10.1111/geb.12567
Pigot AL, Cassey P, Blackburn TM (2018) How to incorporate information on propagule pressure in the analysis of alien establishment success. Methods Ecol Evol 9:1097–1108. https://doi.org/10.1111/2041-210X.12930
Richardson DM, Pyšek P (2006) Plant invasions: merging the concepts of species invasiveness and community invasibility. Prog Phys Geogr 30:409–431. https://doi.org/10.1191/0309133306pp490pr
Sax DF, Gaines SD, Brown JH (2002) Species invasions exceed extinctions on islands worldwide: a comparative study of plants and birds. Am Nat 160:766–783. https://doi.org/10.1086/343877
Schreiber SJ, Lloyd-Smith JO (2009) Invasion dynamics in spatially heterogeneous environments. Am Nat 174:490–505
Seebens H, Blackburn TM, Dyer EE et al (2017) No saturation in the accumulation of alien species worldwide. Nat Commun 8:1–9. https://doi.org/10.1038/ncomms14435
Seebens H, Blackburn TM, Dyer EE et al (2018) Global rise in emerging alien species results from increased accessibility of new source pools. Proc Natl Acad Sci 115:201719429. https://doi.org/10.1073/pnas.1719429115
Shea K, Chesson PL (2002) Community ecology theory as a framework for biological invasions. Trends Ecol Evol 17:170–176. https://doi.org/10.1016/s0169-5347(02)02495-3
Sikes BA, Bufford JL, Hulme PE et al (2018) Import volumes and biosecurity interventions shape the arrival rate of fungal pathogens. PLoS Biol 16:100–101. https://doi.org/10.1371/journal.pbio.2006025
Simberloff D (2009) The role of propagule pressure in biological invasions. Annu Rev Ecol Evol Syst 40:81–102. https://doi.org/10.1146/annurev.ecolsys.110308.120304
Sol D, Maspons J, Vall-Llosera M et al (2012) Unravelling the life history of successful invaders. Science 337:580–583. https://doi.org/10.1126/science.1221523
Stohlgren TJ, Barnett DT, Kartesz JT (1997) The rich get richer: patterns of plant invasions in the United States. Front Ecol Environ 1:11–14. https://doi.org/10.1016/B978-0-7506-5727-3.50005-5
Tilman D (2004) Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. Proc Natl Acad Sci USA 101:10854–10861. https://doi.org/10.1073/pnas.0403458101
Tingley R, Romagosa CM, Kraus F et al (2010) The frog filter: amphibian introduction bias driven by taxonomy, body size and biogeography. Glob Ecol Biogeogr 19:496–503. https://doi.org/10.1111/j.1466-8238.2010.00530.x
Tobin PC, Berec L, Liebhold AM (2011) Exploiting Allee effects for managing biological invasions. Ecol Lett 14:615–624. https://doi.org/10.1111/j.1461-0248.2011.01614.x
van Kleunen M, Dawson W, Essl F et al (2015a) Global exchange and accumulation of non-native plants. Nature 525:100–103. https://doi.org/10.1038/nature14910
van Kleunen M, Dawson W, Maurel N (2015b) Characteristics of successful alien plants. Mol Ecol. https://doi.org/10.1111/mec.13013
Westphal MI, Browne M, MacKinnon K, Noble I (2007) The link between international trade and the global distribution of invasive alien species. Biol Invasions 10:391–398. https://doi.org/10.1007/s10530-007-9138-5
Williamson M (1996) Biological invasions. Chapman & Hall, London
Wilson JRU, Dormontt EE, Prentis PJ et al (2009) Something in the way you move: dispersal pathways affect invasion success. Trends Ecol Evol 24:136–144. https://doi.org/10.1016/j.tree.2008.10.007
Acknowledgements
We thank Julie Lockwood for helpful comments on an earlier version of this manuscript. This work was supported by Australian Research Council Discovery Project grant DP150101839 to RPD, Centre for Invasive Species Solutions project funding (PO1-I-002) to PC, and Leverhulme Trust Grant RPG-2015-073 to TMB, ALP and PC.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Duncan, R.P., Cassey, P., Pigot, A.L. et al. A general model for alien species richness. Biol Invasions 21, 2665–2677 (2019). https://doi.org/10.1007/s10530-019-02003-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-019-02003-y