
Abstract
After interest in keeping crayfish (Crustacea, Decapoda, Astacida) in home aquaria peaked in the mid-2000s, the aquarium trade has become a novel introduction pathway for non-native crayfish species in Germany. Here, we provide an update on the ornamental crayfish trade approximately one decade after the ‘crayfish hype’ to explore the long-term implications in terms of bio-invasion risk. Specifically, species’ availability in e-commerce and potential invasiveness were assessed and compared to previous studies. Morphological and ecological traits of the offered species were compiled and related to their long-term availability (covering 2005–2015). In July 2015, a total of 31 online shops offered 28 crayfish species, which represents a decline of 24 % in species diversity compared to the late 2000s. The estimated rate of import of new species has considerably flattened and approaches pre-hype values (<1 species year−1). However, the risk associated with the offered species, as assessed by a risk screening tool, has not decreased compared to the late 2000s. Long-term availability in the trade was primarily determined by bright coloration, the ability to reproduce under warm aquarium conditions, and a preference for lentic habitats. Species featuring such traits are likely to persist in the aquarium trade and include four high-risk species, most notably invasive and crayfish plague-carrying red swamp crayfish (Procambarus clarkii) and Marmorkrebs (Procambarus fallax f. virginalis). Persistent propagule pressure from aquaria has substantially contributed to the establishment of both species in Germany, stressing the need for more effective pathway management.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
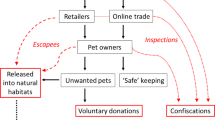
The growing trade with ornamental animals and plants has been identified as a major introduction pathway of aquatic non-native species (Padilla and Williams 2004; Duggan et al. 2006; Laister et al. 2014; Patoka et al. 2015). New trends among aquarium hobbyists that spur an interest in new taxonomic groups can create new opportunities for import and release of aquatic non-native organisms (Chucholl 2013). A prime example for this phenomenon is the trade with ornamental freshwater crayfish (Crustacea, Decapoda, Astacida). The popularity of crayfish as aquarium pets has mainly increased in the last two decades (Chucholl 2013; Patoka et al. 2014a). About 120 species have been recently moved between continents and countries for this purpose, including endangered or potentially invasive species (Chucholl 2015 and citations therein). Several of the traded ornamental species have been deliberately introduced from aquaria into nature or escaped from garden ponds and some species eventually became invasive (Jones et al. 2009; Chucholl et al. 2012; Chucholl 2013; Patoka et al. 2014b).
The stage for much of the trade with ornamental crayfish is Central Europe, in particular Germany (summarized in Chucholl 2015). Keeping of exotic crayfish in home aquaria for ornamental purposes started first in Germany in the late 1980s. At this time, exotic crayfish from the live food trade began to made sporadic appearances in the aquarium trade (Lietsch 1984; Schlüter 1989; Werner 1993). Between the mid- and late-1990s, keeping of crayfish in home aquaria became increasingly popular, which prompted the import of several new species of North American and Australasian origin (Werner 1998; Gonella 1999). By the late 1990s, approximately 15 species were available for home aquaria. In the early 2000s, keeping of exotic crayfish had eventually turned into a widespread trend among aquarium hobbyists in Germany and Austria. To meet the growing market demand, new species were imported at a very high rate from Central and North America as well as the western part of New Guinea (Pekny and Lukhaup 2005). Around the same time, a considerable number of freshwater shrimps and mollusks, mostly from the Afrotropical, Indomalayan, and Australasian ecozones were also imported and marketed (Pekny and Lukhaup 2005). This phenomenon was later dubbed as ‘invertebrates boom’ (Bitter 2006) and by 2005, a total number of 74 non-native crayfish species has been imported into Germany or Austria (Pekny and Lukhaup 2005). Most recently, it was shown that until mid-2009, a total number of 120 crayfish species has been available for home aquaria (Chucholl 2013).
This number represents roughly one fifth of the global species pool of crayfish and 42-fold the number of native crayfish species to be found in Germany (i.e. three; Chucholl 2015 and citations therein). Many of the imported species, however, have been only rarely available and only approximately 20 species have been commonly offered (Chucholl 2013). Similar findings were reported from other European countries, including the Netherlands (Soes and Koese 2010), Great Britain (Peay et al. 2010), the Czech Republic (Patoka et al. 2014a, b), Slovakia (Lipták and Vitázková 2015), Greece (Papavlasopoulou et al. 2014) and Italy (Tricarico et al. 2010), albeit these involved typically fewer species.
Using a risk assessment tool (Freshwater Invertebrate Invasiveness Scoring Kit), 4 out of the 20 common crayfish species in the German ornamental trade have been assessed as high-risk species (Chucholl 2013). These four species, namely Procambarus clarkii, Procambarus alleni, Procambarus fallax f. virginalis, and Cherax destructor are widespread in the trade and have already been deliberately introduced from aquaria into European inland waters (Chucholl 2015 and citations therein). Such releases have raised great concern because their omnivorous feeding habits, foraging and burrowing activity, and size make crayfish key players in aquatic ecosystems and invasive non-native crayfish can wreak ecological havoc (Nyström 1999; Rodríguez et al. 2005; Vaeßen and Hollert 2015). Crayfish of North American origin, such as Procambarus, are also notorious for displacing native European crayfish due to interference competition and transmission of crayfish plague, a fatal disease of native crayfish (Holdich et al. 2009; Svoboda et al. 2016; Chucholl 2016).
Species popularity among aquarium hobbyists can be dynamic, however, which might affect the availability of high-risk species (cf. Chucholl 2015). The main purpose of this study was therefore twofold: (1) we provide an update on the ornamental crayfish trade in Germany approximately one decade after the ‘crayfish hype’ in order to explore the long-term implications in terms of bio-invasion risk. Specifically, species’ availability and potential invasiveness were assessed and the results compared to those of previous studies. (2) A dataset on morphological and ecological species traits was used to identify the determinants of long-term availability in the trade, which was taken as surrogate measure for species popularity. Understanding the drivers of species popularity is important for risk assessment because popularity has been shown to be positively related to the likelihood of release into nature, i.e. propagule pressure (Duggan et al. 2006; Chucholl 2013).
Methods
Data collection
Following Chucholl (2013), online shops offering ornamental crayfish were searched using the German terms for crayfish, crustacean, shrimp, and invertebrates. From each identified shop, all of the offered crayfish species were recorded. Species’ availability was subsequently assessed as the proportion of shops that offered the respective species. As in previous studies, availability in the trade was taken as surrogate measure for the popularity in home aquaria (Duggan et al. 2006; Chucholl 2013; Patoka et al. 2014a). Online shops were chosen because they are more easily accessible than conventional pet stores and because online availability has been previously shown to be positively correlated with general availability in the pet trade (Chucholl 2013). The shops were analyzed over a period of 1 month, beginning in July 2015.
In the meantime, Cherax ‘Orange Tip’, an ornamental crayfish from West Papua (Indonesia), was described as new species Cherax snowden (Lukhaup et al. 2015). At the time of data collection, Cherax ‘Orange Tip’ was considered a color variety of Cherax holthuisi, its closest related species (Lukhaup et al. 2015), and is here therefore included within C. holthuisi (also for consistency with previous studies; cf. Chucholl 2013).
Risk assessment
The potential invasiveness and risk of the offered species was rated using the Freshwater Invertebrate Invasiveness Scoring Kit (FI-ISK, v.1.19; Tricarico et al. 2010). FI-ISK is an adaption of the Weed Risk Assessment for freshwater invertebrates and assesses species’ biogeography and history, the presence of ‘undesirable traits’, as well as species’ biology and ecology based on 49 questions. The FI-ISK output represents a risk score, with high numbers indicating high-risk species with a high potential invasiveness in the given reference region (Germany in the case of this study). The FI-ISK scoring system was calibrated for crayfish by Tricarico et al. (2010), who found a statistically appropriate threshold score for high-risk species of ≥16. The tool has been previously used to assess the potential invasiveness of ornamental crayfish (Chucholl 2013; Papavlasopoulou et al. 2014; Patoka et al. 2014a).
Determinants of availability
To assess the drivers of crayfish species’ availability in the pet trade we first compiled a list of traded species. Specifically, we used a dataset on ornamental crayfish imported into Germany until 2009 as starting point (Chucholl 2013). The dataset was updated with crayfish species imported after 2009 and long-term general availability (covering one decade, i.e. 2005–2015) was reassessed for all species on an ordinal scale as either ‘very rare’ (only available for a short time and in low numbers), ‘rare’ (available occasionally in low numbers), ‘common’ (frequently available in low numbers), or ‘very common’ (always available) following Chucholl (2013) and Patoka et al. (2014a).
Based on related studies (Zeng et al. 2015) and probable preferences of aquarium hobbyists (inferred from personal communications), a total of 11 species traits was then considered to predict crayfish species’ general availability in the trade. Specifically, species’ maximum size (carapace length in mm; continuous variable), maximum fecundity (pleopodal eggs female−1; continuous variable), habitat (categorical with the levels ‘lentic’, ‘lotic’, ‘indifferent’, and ‘semi-terrestrial’), native range (biogeographic region; categorical), taxonomic position (family; categorical) and availability in the live food trade (binary variable) were used as independent variables. In addition, four potentially appealing traits were assessed and included as binary predictors: multicolored (presence of at least two distinct body colors), color pattern (presence of prominent stripes or spots), bright coloration (presence of vivid body color), and impressive chelae (presence of large, massive chelae). Color and chelae traits were recorded by the same person by examining color pictures of adult crayfish in crayfish reference books (Lukhaup and Pekny 2005, 2008) or online shops. Finally, a species’ ability to reproduce under typical warm-water conditions in home aquaria (i.e. mean temperature >20 °C) was included as binary predictor. This ability was considered as given when the species is known to reproduce in captivity at constantly warm temperatures (cf. Lukhaup and Pekny 2005). In the absence of data from captivity, the ability to breed in constantly warm water was rated as given for species originating from tropical or subtropical climates or species that breed during summer (excluding temperate species that lay eggs in fall or spring, i.e. when water temperatures are low). Ecological, biogeographic and phylogenetic data were compiled from Hobbs (1974, 1989), Holthuis (1986), Hobbs et al. (1989), Holdich (2002), Taylor and Schuster (2004), Beatty et al. (2005), Seitz et al. (2005), Lukhaup and Pekny (2005, 2006, 2008), Souty-Grosset et al. (2006), Adams (2008), Lukhaup and Herbert (2008), Holdich et al. (2009), and Zeng et al. (2015), and information on all of the above mentioned traits could be obtained for a total of 81 species.
Statistical analyses
All statistical analyses were performed in the software package R (version 3.1.3; R Core Team 2015) and figures were produced in SigmaPlot 10.0. The cumulative number of crayfish species available for home aquaria in Germany was described by a sigmoidal four-parameter regression curve. The rate of import of new species was subsequently estimated from the first derivative of the regression fit. To assess possible changes in the bio-invasion risk posed by the traded species, FI-ISK scores of the species offered in online stores were compared between the years 2015 and 2009 (data taken from Chucholl 2013) using a Mann–Whitney rank sum test. To account for different availability of the species, the same test was also performed on weighted FI-ISK scores, with weights obtained from species availability in the respective study year. To further characterize the pool of species offered online, we calculated the Shannon Index (H′) and the species evenness (E) and compared the indices between the years 2009 and 2015 (cf. Heip et al. 1998). Species availability in the different years was taken as measure of abundance.
Determinants of general availability in the trade were explored using two complementary statistical approaches, namely recursive partitioning for classification and ordered logistic regression, to see if different methods arrive at a similar conclusion, thus making it more robust. For both analyses, general availability was implemented as an ordered response variable with four levels.
Firstly, we used recursive partitioning for classification (Breiman et al. 1984; Strobl et al. 2009). The method is widely used for multivariable analysis and Data Mining, and generates a binary decision tree that recursively splits the data into subsets. The resulting classification tree predicts membership of objects in the classes of the response variable (see Breiman et al. 1984 for a comprehensive description). The tree was constructed in R using the ‘rpart’ function of the ‘rpart’ package (Therneau et al. 2015) and the 11 species traits outlined above were entered as independent variables. The minimum number of observations in a node before attempting a split was set to 5 and the cost complexity factor was left at the default setting of 0.001. To avoid any overfitting of the data, the tree was pruned using the complexity parameter so that the 10-fold cross-validated error was least (cf. Breiman et al. 1984; Therneau et al. 2015).
Secondly, we used a more classical regression approach, i.e. ordered logistic regression as implemented by the function ‘polr’ of the R package ‘MASS’ (Venables and Ripley 2002), to predict general availability from the 11 candidate variables. To arrive at the optimal model, uninformative independent variables were removed from the full model containing all variables by stepwise backward selection based on the Akaike information criterion (AIC). The AIC is a measure of the goodness of model fit with a penalty for model complexity and is widely used for model selection. If the removal of a variable improved the AIC, it was dropped from the candidate model. Otherwise it was retained and the next variable was removed. The significance of the predictor variables contained in the final optimal model was assessed by analysis of deviance (likelihood ratio tests).
As a measure of accuracy, the overall accuracy rate and Cohen’s Kappa statistic (κ) were calculated for both statistical methods. κ is a robust measure of performance that is little affected by the prevalence of the categorical classes (levels). Values range between −1 and 1, with 1 indicating perfect agreement and values close to or below 0 indicating a performance no better or equal than a random model (Cohen 1960). In addition, a one-sided test was performed to assess if the accuracy is better than the ‘no information rate’, which is taken to be the largest class percentage in the data (cf. Kuhn 2008). Using analysis of deviance, the significance of the ordered logistic regression model was also tested against a null model containing only a constant predictor.
Results
Available species
The online survey yielded a total of 31 online shops, which offered 28 crayfish species (Fig. 1 and Fig. S1 in the supplementary material, Online Resource 1). Only two of those species were imported after 2009, indicating that the rate of import has considerably flattened compared to the late 2000s and approaches pre-hype values (<1 species year−1) (summarized in Fig. 2a).

Data for 2009 were taken from Chucholl (2013)
Availability (left panel) and potential invasiveness (as assessed by FI-ISK; right panel) of freshwater crayfish offered through German online shops in 2009 (white bars) and 2015 (grey bars). Black bars represent high-risk species that exceed the respective FI-ISK threshold (score >16; hatched grey line). ‘CPO’ means Cambarellus patzcuarensis ‘Orange’.

The ‘crayfish hype’ among German aquarium hobbyists in terms of imported species (a) and the resulting propagule pressure of Marmorkrebs (Procambarus fallax f. virginalis) (b), as assessed by independent records of free-living individuals. a Cumulative number of crayfish species available for home aquaria in Germany (black dots) in relation to time. Solid blue line denotes a sigmoidal four-parameter regression curve fitted to the data (adjusted R 2 > 0.99; P < 0.001) and dashed lines represent the 95 %-confidence band. The solid red curve shows the estimated rate of import (species year−1), as assessed as first derivative of the regression fit. Data sources (by year) are as follows: 1984—Lietsch (1984); 1989—Schlüter (1989); 1993—Werner (1993); 1999—Werner (1998) and Gonella (1999); 2005—Pekny and Lukhaup (2005); 2009—Chucholl (2013); 2012—Chucholl (2015); and 2015—present study. b Cumulative number of verified Marmorkrebs records in Germany (based on Chucholl et al. 2012 and updated with new data). All records of free-living Marmorkrebs (picture) in Germany most likely originate from releases from aquaria
Compared to 2009, the number of crayfish species offered online has declined by 24 % (Fig. 3a). H′ (2.8) and E (0.6) of the 2015 sample were both lower than the corresponding indices for the 2009 sample (3.2 and 0.7, respectively), corroborating a trend towards fewer species, of which most are only rarely available (cf. Fig. 1). Specifically, seven previously offered Procambarus species were not present in the 2015 sample, along with, two Cambarellus, two Orconectes, one Cambarus and one Cherax species (cf. Fig. 3a). The 2015 sample, by contrast, included native noble crayfish (Astacus astacus), the old invader signal crayfish (Pacifastacus leniusculus) and two recently imported Cherax species from New Guinea (C. monticola and C. communis) as new species. Except for noble crayfish, however, all of the newly offered species were only available from <5 % of the analyzed shops (cf. Fig. 1).

Data for 2009 were taken from Chucholl (2013)
Number of species per genus (a) and potential invasiveness (b), as assessed by FI-ISK, of freshwater crayfish offered through online shops in Germany in 2009 and 2015. The hatched horizontal line in b indicates the FI-ISK score threshold above which species are considered as high risk. Hatched lines in boxes show the mean and outliers are shown as dots. Other genera include Astacus, Pacifastacus, Orconectes, and Cambarus spp.
The five most commonly available crayfish species in 2015 were Cambarellus patzcuarensis ‘Orange’ (commonly referred to as ‘CPO’; Fig. S1A) (offered by 97 % of shops), Procambarus clarkii (offered by 63 % of shops; Fig. S1C,D), Procambarus alleni (blue color morph; Fig. S1B) (offered by 50 % of shops), Cambarellus diminutus (offered by 47 % of shops), and A. astacus and Cherax holthuisi (both offered by 33 % of shops). Except for A. astacus and C. diminutus, all of these species were also among the five most commonly offered species in 2009 (see Fig. 1). Most strikingly, CPO was the most frequently offered species in both study years.
Risk assessment
The bio-invasion risk associated with the offered species, as assessed by FI-ISK, did not differ between 2015 and 2009 (Mann–Whitney rank sum test: U = 488.5, P = 0.429; Fig. 3b). Similarly, the weighted FI-ISK scores, accounting for differences in availability among species, did not vary between years (Mann–Whitney rank sum test: U = 591.5, P = 0.613). The 2015 sample included a total of six high-risk species, namely signal crayfish (FI-ISK score = 39), P. clarkii (FI-ISK score = 33), Orconectes limosus (FI-ISK score = 24), P. alleni (FI-ISK score = 22), C. destructor (FI-ISK score = 22), and P. fallax f. virginalis (FI-ISK score = 20). Except for signal crayfish, all of these high-risk species have been also available in 2009, in addition to Orconectes neglectus (FI-ISK score = 18), which was not offered in 2015. An overview of the risk assessments and the species availability in the two study years is given in Fig. 1.
Determinants of availability
The classification tree with species traits to predict general availability is shown in Fig. 4, and the outcome of the ordered logistic regression analysis is summarized in Table 1. Accuracy of the regression tree was 0.88 (95 % CI: 0.79–0.94) and κ was 0.75. Accuracy of the optimal regression model was 0.78 (95 % CI: 0.67–0.86) and κ was 0.54. Both statistical methods performed better than the ‘no information rate’ (P < 0.001 and P = 0.020 for the classification tree and the regression model, respectively). The regression model (AIC = 101.8, residual deviance = 77.8) performed also significantly better than the respective null model with a constant predictor (AIC Null = 168.2, residual devianceNull = 162.2; likelihood ratio test: P < 0.001).

Classification tree with crayfish species traits to predict general availability in the German pet trade (Accuracy = 0.88, κ = 0.75, and P < 0.001; one-sided test against no information rate). Numbers on top of bars indicate the respective number of species. Abbreviations for general availability are as follows: VR very rare, R rare, C common, and VC very common
Both analyses concordantly suggest that long-term availability of crayfish in the pet trade is primarily determined by bright coloration, the ability to reproduce under warm aquarium conditions, and a preference for lentic habitats (cf. Fig. 4; Table 1). In addition, species’ size (carapace length) was negatively related to general availability in both models. Other explanatory variables were less important and differed between the two statistical methods. The classification tree produced a split based on availability in the live food trade (Fig. 4), while the optimal ordered regression model included crayfish family and impressive chelae as further explanatory variables (Table 1).
Discussion
Our results indicate that ornamental crayfish have become a permanent element of the German pet trade, notwithstanding that the trend of keeping crayfish as pets may have passed its peak. The associated bio-invasion risk has remained at a high level due to positive selection of several high-risk species by the trade. Compared to the late 2000s, the number of offered species has declined by about one fourth and the estimated rate of import of new species has approached values before the onset of the trend in the mid-1990s (Fig. 2a). In addition, most of the available species were only rarely offered (cf. Fig. 1). Specifically, only three species were offered by more than 50 % of the analyzed shops, namely CPO, P. clarkii, and P. alleni. However, six out of the 28 available species (i.e. 20 %) were rated as high-risk, indicating a high potential invasiveness, and four out of the six high-risk species (i.e. 67 %) were found to exhibit traits that promote popularity, thereby favoring their long-term persistence in the pet trade. Indeed, two out of the three most commonly offered species were high-risk species (P. clarkii and P. alleni).
The contemporary trade of ornamental crayfish in Germany, as assessed by the present shop survey, is dominated by only a few species. Interestingly, this resembles the situation that has been previously reported for other EU countries. Soes and Koese (2010) analyzed classified online sites in the Netherlands and report that Procambarus and Cambarellus were the most popular genera, followed by Cherax. Among Procambarus, P. clarkii was the most commonly offered species, followed by P. fallax f. virginalis and P. alleni. In Great Britain, Peay et al. (2010) found that 13 non-native crayfish species had been imported for the ornamental trade. These species included five Cherax spp., three Procambarus spp. (namely P. alleni, P. clarkii, and P. fallax f. virginalis), and two dwarf crayfish species (Cambarellus spp.). Papavlasopoulou et al. (2014) found that a total of eight ornamental crayfish species were available through e-commerce in Greece. Each three of the available species were from the genera Cambarellus (C. shufeldtii, C. montezumae, and CPO) and Cherax (C. destructor, C. peknyi, and C. quadricarinatus), and two belonged to the genus Procambarus (P. clarkii and P. alleni). Most recently, Patoka et al. (2014a, b) reported a total of 28 crayfish species to be marketed in the Czech Republic. Only ten of these species were classified as either common or very common in the trade. Seven of those species were from the genus Cherax, two belonged to Procambarus (P. clarkii and P. fallax f. virginalis), and one was a dwarf crayfish (C. patzcuarensis). Beyond Europe, Faulkes (2015a) reported a largely comparable set of ornamental crayfish species to be traded in North America.
Overall, the ornamental crayfish trade is strongly dominated by a few species in the genera Cambarellus, Cherax and Procambarus (Pekny and Lukhaup 2005; Peay et al. 2010; Chucholl 2013; Papavlasopoulou et al. 2014; Patoka et al. 2014a; Faulkes 2015a, b; cf. Fig. S1). Among Cambarellus, CPO is clearly the most popular species (Chucholl 2013; Patoka et al. 2014a). Among Cherax species, C. quadricarinatus and C. peknyi are widespread (Chucholl 2013; Papavlasopoulou et al. 2014; Patoka et al. 2014a); however, other species are also commonly offered, including high-risk C. destructor (Chucholl 2013; Patoka et al. 2014a). Among Procambarus species, P. clarkii, P. alleni, and P. fallax f. virginalis are clearly the most available species (Pekny and Lukhaup 2005; Peay et al. 2010; Soes and Koese 2010; Chucholl 2013; Papavlasopoulou et al. 2014; Patoka et al. 2014a). These rather consistent trends in species’ availability and, therefore, popularity among the analyzed countries suggest that availability in the trade is shaped by a common mechanism.
In line with this inference, our analysis indicates that long-term availability in the trade can be directly related to species’ traits. As intuitively expected (cf. Lukhaup and Pekny 2005; Chucholl 2015; Faulkes 2015b), popular ornamental crayfish species feature bright body coloration and are able to reproduce at typical warm-water aquarium conditions (cf. Fig. S1). A small body size and a preference or tolerance of lentic habitats further increase the probability of a species to become popular, in particular in the absence of bright body coloration (cf. Fig. 4). These determinants of availability were supported by both statistical methods (classification tree and ordered regression), indicating a robust pattern. The classification tree further indicated that availability in the live food trade can increase availability in the ornamental trade. This finding is consistent with early reports on ornamental crayfish in Germany, stating that species from the food trade, namely red swamp crayfish and the Australian species Yabby (Cherax destructor) and red claw (Cherax quadricarinatus), made sporadic appearances in the aquarium trade (Schlüter 1989; Werner 1993). Similar drivers of popularity can be expected for other animal groups, such as ornamental fish, shrimps or mollusks (cf. Bitter 2006; Maceda-Veiga et al. 2016).
Crayfish species featuring traits that promote popularity as pets are likely to be positively selected and to persist in the aquarium trade. As a worrying fact, species featuring such traits include four species with a high potential invasiveness (as assessed by FI-ISK), most notably P. clarkii, P. alleni, and P. fallax f. virginalis. All of these Procambarus were rated as high-risk species and captive populations are known reservoir hosts of crayfish plague (Keller et al. 2014; Mrugała et al. 2014; Svoboda et al. 2016). Persistent propagule pressure from aquaria has substantially contributed to the establishment of both P. clarkii and P. fallax f. virginalis in Germany (Chucholl 2013, 2015, and citations therein). The estimated contemporary propagule pressure of P. fallax f. virginalis exceeds two independent release events per year (see Fig. 2b). In addition, the “Florida lobster” (blue color morph of P. alleni), was recently reported from the Rhine River (Gross 2013), further corroborating the results of Chucholl (2013) whereby widely available pet crayfish exceeding a certain size threshold are likely to be released into nature. Finally, albeit rarely offered, the availability of well-known high-risk invaders, such as signal crayfish (P. leniusculus) and spiny-cheek crayfish (Orconectes limosus), in the ornamental trade is further disturbing (cf. Holdich et al. 2009; Chucholl 2016).
Although there are several high-risk species among popular pet crayfish, some of the traits that promote popularity may reduce species’ potential invasiveness. First, the selection for reproduction at warm temperatures might decrease species’ potential invasiveness in temperate climates. This holds true for some tropical Cherax species (e.g., C. quadricarinatus; Veselý et al. 2015); however, some species of subtropical or tropical origin, such as P. clarkii, P. fallax f. virginalis, and C. destructor, have the ability to withstand winter conditions in temperate zone (Veselý et al. 2015) and to establish reproducing populations (Chucholl 2011; Chucholl et al. 2012). Second, bright color morphs, such as C. patzcuarensis ‘Orange’ or blue P. alleni (Fig. S1A,B), may experience a higher risk of predation than wild type colored individuals (cf. Faulkes 2015a), which might lower their probability of establishment. There is, however, little empirical evidence to support this notion, and the naturally bright body coloration of P. clarkii (Fig. S1C) does not seem to interfere with its remarkable success as invasive species (Souty-Grosset et al. 2006).
‘Beautiful invaders’, such as P. clarkii, may be also hard to be substituted with benign low-risk species as part of a voluntary self-regulation of the trade (cf. Chucholl 2013; Mengedoht 2014), if the low-risk species featured few or no traits that promote popularity. Ideally, exotic species should be substituted with native species. Native European crayfish species, such as noble crayfish, however, are unlikely to replace exotic crayfish as pets due to their cool temperature requirements for reproduction and unappealing coloration (Pekny and Lukhaup 2005; Fig. S1H). Low- to medium-risk Cherax species from Australasia are more promising in this regard (Chucholl 2013; Patoka et al. 2014a) but have as yet failed to supersede the high-risk Procambarus species as most commonly offered species (cf. Fig. 1). This is despite public education efforts to favor Cherax species over high-risk, crayfish plague-carrying Procambarus species (e.g., Mengedoht 2014).
Conclusion
Pre-introduction selection of high-risk species based on traits that promote popularity as pets hampers risk mitigation and calls for effective pathway management (cf. Zeng et al. 2015). If no action is taken, the availability of high-risk species positively selected by the trade, such as P. clarkii, will remain high. This would be an undesirable situation because availability in the pet trade is positively related to propagule pressure (Duggan et al. 2006) and promotes thereby introduction and establishment (Lockwood et al. 2005; Chucholl 2015).
Effective pathway management requires a cut-back of the availability of high risk species (cf. Duggan et al. 2006; Chucholl 2013). Trade regulation of high-risk species is a promising tool in this regard and the new EU regulation on invasive alien species (IAS) allows for this risk mitigation measure (EU Regulation 1143/2014). Red swamp crayfish (P. clarkii), Marmorkrebs (P. fallax f. virginalis), signal crayfish (P. leniusculus), and spiny-cheek crayfish (O. limosus) are included on the list of IAS of Union concern (European Commission 2016). Species on this ‘black list’ can be subjected to trade regulations, as well as a ban on import and keeping. The need for these risk mitigation measures is strongly supported by the outcome of the present study. Finally, the data on contemporary species availability may serve as important baseline to assess the effectiveness of potential Union-wide regulations in the future.
References
Adams SB (2008) Cambarellus shufeldtii. Version 1.0. USDA Forest Service, Crayfishes of Mississippi website, Oxford, MS. http://maps.fs.fed.us/crayfish/factsheets/FS0056.pdf. Accessed 23 Aug 2009
Beatty S, Morgan D, Gill H (2005) Role of life history strategy in the colonisation of Western Australian aquatic systems by the introduced crayfish Cherax destructor Clark, 1936. Hydrobiologia 549:219–237
Bitter F (ed) (2006) Wirbellose im Süßwasseraquarium—Garnelen, Krebse, Krabben & Co. Aquaristik Sonderheft 1, Dähne-Verlag, Ettlingen
Breiman L, Friedman J, Stone CJ, Olshen RA (1984) Classification and regression trees. CRC Press, Boca Raton
Chucholl C (2011) Population ecology of an alien “warm water” crayfish (Procambarus clarkii) in a new cold habitat. Knowl Manag Aquat Ecosyst 401:29. doi:10.1051/kmae/2011053
Chucholl C (2013) Invaders for sale: trade and determinants of introduction of ornamental freshwater crayfish. Biol Invasions 15:125–141. doi:10.1007/s10530-012-0273-2
Chucholl C (2015) Marbled crayfish gaining ground in Europe: the role of the pet trade as invasion pathway. In: Kawai T, Faulkes Z, Scholtz G (eds) Freshwater crayfish: a global overview. CRC Press, Boca Raton, pp 83–114
Chucholl C (2016) The bad and the super-bad: prioritising the threat of six invasive alien to three imperilled native crayfishes. Biol Invasions. doi:10.1007/s10530-016-1141-2
Chucholl C, Morawetz K, Groß H (2012) The clones are coming-strong increase in Marmorkrebs [Procambarus fallax (Hagen, 1870) f. virginalis] records from Europe. Aquat Invasions 7:511–519. doi:10.3391/ai.2012.7.4.008
Cohen J (1960) A coefficient of agreement for nominal scales. Educ Psychol Meas 20:37–46
Duggan IC, Rixon CAM, MacIsaac HJ (2006) Popularity and propagule pressure: determinants of introduction and establishment of aquarium fish. Biol Invasions 8:377–382. doi:10.1007/s10530-004-2310-2
European Commission (2016) Commission implementing regulation (EU) 2016/1141 of 13 July 2016 adopting a list of invasive alien species of union concern pursuant to regulation (EU) No 1143/2014 of the European parliament and of the council. Off J EU 59:4–8
Faulkes Z (2015a) Marmorkrebs (Procambarus fallax f. virginalis) are the most popular crayfish in the North American pet trade. Knowl Manag Aquat Ecosyst 416:20. doi:10.1051/kmae/2015016
Faulkes Z (2015b) The global trade in crayfish as pets. Crustac Res 44:75–92. doi:10.18353/crustacea.44.0_75
Gonella H (1999) Krebse, Krabben und Garnelen im Süßwasseraquarium. Bede-Verlag, Ruhmannsfelden
Gross H (2013) Blauer Floridakrebs (Procambarus alleni) im Rhein! Forum Flusskrebse 19:33–35
Heip CH, Herman PM, Soetaert K (1998) Indices of diversity and evenness. Océanis, Paris
Hobbs HH (1974) A checklist of the North and Middle American crayfishes (Decapoda: Astacidae and Cambaridae). Smithson Contrib Zool 166:161
Hobbs HH (1989) An illustrated checklist of the American crayfishes (Decapoda: Astacidae, Cambaridae, and Parastacidae). Smithson Contrib Zool 480:236
Hobbs HH, Jass JP, Huner JV (1989) A review of global crayfish introductions with particular emphasis on two North American species (Decapoda, Cambaridae). Crustaceana 56:299–316
Holdich DM (ed) (2002) Biology of freshwater crayfish. Blackwell Science, London
Holdich DM, Reynolds JD, Souty-Grosset C, Sibley PJ (2009) A review of the ever increasing threat to European crayfish from non-indigenous crayfish species. Knowl Manag Aquat Ecosyst 394–395:11. doi:10.1051/kmae/2009025
Holthuis LB (1986) The freshwater Crayfish of New Guinea. Freshw Crayfish 6:48–58
Jones JPG, Rasamy JR, Harvey A et al (2009) The perfect invader: a parthenogenic crayfish poses a new threat to Madagascar’s freshwater biodiversity. Biol Invasions 11:1475–1482. doi:10.1007/s10530-008-9334-y
Keller NS, Pfeiffer M, Roessink I et al (2014) First evidence of crayfish plague agent in populations of the marbled crayfish (Procambarus fallax forma virginalis). Knowl Manag Aquat Ecosyst 414:15. doi:10.1051/kmae/2014032
Kuhn M (2008) Building predictive models in R using the caret package. J Stat Softw 28:1–26
Laister G, Lehmann G, Martens A (2014) Exotic Odonata in Europe. Odonatologica 43:125–135
Lietsch P (1984) Zur Pflege und Zucht des südamerikanischen Sumpfkrebses (Procambarus troglodytes). AT 31:313
Lipták B, Vitázková B (2015) Beautiful, but also potentially invasive. Ekológia (Bratislava) 34:155–162
Lockwood JL, Cassey P, Blackburn T (2005) The role of propagule pressure in explaining species invasions. Trends Ecol Evol 20:223–228. doi:10.1016/j.tree.2005.02.004
Lukhaup C, Pekny R (2005) Krebse im Aquarium. Dähne Verlag, Ettlingen
Lukhaup C, Pekny R (2006) Cherax (Cherax) holthuisi, a new species of crayfish (Crustacea: Decapoda: Parastacidae) from the centre of the Vogelkop Peninsula in Irian Jaya (West New Guinea), Indonesia. Zool Med Leiden 80–1:101–107
Lukhaup C, Herbert B (2008) A new species of freshwater crayfish (Crustacea: Decapoda: Parastacidae) from the Fly River drainage, Western Province, Papua New Guinea. Mem Queensl Mus 52:213–219
Lukhaup C, Pekny R (2008) Süßwasserkrebse aus aller Welt. Dähne Verlag, Ettlingen
Lukhaup C, Panteleit J, Schrimpf A (2015) Cherax snowden, a new species of crayfish (Crustacea, Decapoda, Parastacidae) from the Kepala Burung (Vogelkop) Peninsula in Irian Jaya (West Papua), Indonesia. ZooKeys 518:1–14. doi:10.3897/zookeys.518.6127
Maceda-Veiga A, Domínguez-Domínguez O, Escribano-Alacid J, Lyons J (2016) The aquarium hobby: can sinners become saints in freshwater fish conservation? Fish Fish 17:860–874. doi:10.1111/faf.12097
Mengedoht O (2014) Hilft nur ein Selbstverzicht? DATZ 4:28–31
Mrugała A, Kozubíková-Balcarová E, Chucholl C et al (2014) Trade of ornamental crayfish in Europe as a possible introduction pathway for important crustacean diseases: crayfish plague and white spot syndrome. Biol Invasions. doi:10.1007/s10530-014-0795-x
Nyström P (1999) Ecological impact of introduced and native crayfish on freshwater communities: European perspectives. Crustac Issues 11:63–86
Padilla DK, Williams SL (2004) Beyond ballast water: aquarium and ornamental trades as sources of invasive species in aquatic ecosystems. Front Ecol Environ 2:131–138
Papavlasopoulou I, Perdikaris C, Vardakas L, Paschos I (2014) Enemy at the gates: introduction potential of non-indigenous freshwater crayfish in Greece via the aquarium trade. Cent Eur J Biol 9:11–18. doi:10.2478/s11535-013-0120-6
Patoka J, Kalous L, Kopecký O (2014a) Risk assessment of the crayfish pet trade based on data from the Czech Republic. Biol Invasions 16:2489–2494. doi:10.1007/s10530-014-0682-5
Patoka J, Petrtýl M, Kalous L (2014b) Garden ponds as potential introduction pathway of ornamental crayfish. Knowl Manag Aquat Ecosyst 414:13
Patoka J, Bláha M, Devetter M et al (2015) Aquarium hitchhikers: attached commensals imported with freshwater shrimps via the pet trade. Biol Invasions. doi:10.1007/s10530-015-1018-9
Peay S, Holdich D, Brickland J (2010) Risk assessments of non-indigenous crayfish in Great Britain. Freshw Crayfish 17:109–122
Pekny R, Lukhaup C (2005) Aquarienkrebse in Europa—eine rasante Entwicklung! 2. Int Flusskrebstagung Baden 78–94
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rodríguez C, Bécares E, Fernández-Aláez M, Fernández-Aláez C (2005) Loss of diversity and degradation of wetlands as a result of introducing exotic crayfish. Biol Invasions 7:75–85
Schlüter M (1989) Flußkrebse aus Australien. Ritter in farbenfroher Rüstung. DATZ 42:526–528
Seitz R, Vilpoux K, Hopp U, Harzsch S, Maier G (2005) Ontogeny of the Marmorkrebs (Marbled Crayfish): a parthenogenetic crayfish with unknown origin and phylogenetic position. J Exp Zool 303:393–405
Soes M, Koese B (2010) Invasive crayfish in the Netherlands: a preliminary risk analysis. Interim report, Bureau Waardenburg bv, Stichting EIS-Nederland, Invasive Alien Species Team, Waardenburg
Souty-Grosset C, Holdich DM, Noel PY, Reynolds JD, Haffner P (eds) (2006) Atlas of Crayfish in Europe. Museum national d`Histoire naturelle, Paris (Patrimoines naturels, 64)
Strobl C, Malley J, Tutz G (2009) An introduction to recursive partitioning: rationale, application, and characteristics of classification and regression trees, bagging, and random forests. Psychol Methods 14:323
Svoboda J, Mrugała A, Kozubíková-Balcarová E, Petrusek A (2016) Hosts and transmission of the crayfish plague pathogen Aphanomyces astaci: a review. J Fish Dis. doi:10.1111/jfd.12472
Taylor CA, Schuster GA (2004) Crayfishes of Kentucky. Illinois natural history survey, Special Publication 28, Carbondale
Therneau T, Atkinson B, Ripley B (2015) rpart: recursive Partitioning and Regression Trees. R package version 4.1–9.https://github.com/cran/rpart. Accessed 12 Aug 2015
Tricarico E, Vilizzi L, Gherardi F, Copp GH (2010) Calibration of FI-ISK, an Invasiveness Screening Tool for Nonnative Freshwater Invertebrates. Risk Anal 30:285–292. doi:10.1111/j.1539-6924.2009.01255.x
Vaeßen S, Hollert H (2015) Impacts of the North American signal crayfish (Pacifastacus leniusculus) on European ecosystems. Environ Sci Eur. doi:10.1186/s12302-015-0065-2
Venables WN, Ripley BD (2002) Modern Applied Statistics with S. Fourth, Springer, New York
Veselý L, Buřič M, Kouba A (2015) Hardy exotics species in temperate zone: can “warm water” crayfish invaders establish regardless of low temperatures? Sci Rep 5:16340. doi:10.1038/srep16340
Werner U (1993) Ausgefallene Aquarienpfleglinge. Landbuch-Verlag, Hannover
Werner U (1998) Garnelen, Krebse und Krabben im Süßwasseraquarium. Aqualog Special, Verlag A.C.S., Mörfelden-Walldorf
Zeng Y, Chong KY, Grey EK et al (2015) Disregarding human pre-introduction selection can confound invasive crayfish risk assessments. Biol Invasions. doi:10.1007/s10530-015-0881-8
Acknowledgments
We are grateful to A. Martens, R. Wegner (both PH Karlsruhe), and S. Nehring (BfN) for kindly providing records of free-living Marmorkrebs. The helpful comments and suggestions made by I. Duggan (Editor) and two anonymous reviewers are also gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Chucholl, C., Wendler, F. Positive selection of beautiful invaders: long-term persistence and bio-invasion risk of freshwater crayfish in the pet trade. Biol Invasions 19, 197–208 (2017). https://doi.org/10.1007/s10530-016-1272-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-016-1272-5