
Abstract
Human disturbances, such as anchoring and dredging, can cause physical removal of seagrass rhizomes and shoots, leading to the fragmentation of meadows. The introduced green alga, Caulerpa racemosa, is widely spread in the North-West Mediterranean and, although it can establish in both degraded and pristine environments, its ability to become a dominant component of macroalgal assemblages seems greater in the former. The aim of this study was to estimate whether the spread of C. racemosa depends on the intensity of disturbance to the canopy structure of Posidonia oceanica. A field experiment was started in July 2010 when habitat complexity of a P. oceanica meadow was manipulated to simulate mechanical disturbances of different intensity: rhizome damage (High disturbance intensity = H), leaf removal (Low disturbance intensity = L), and undisturbed (Control = C). Disturbance was applied within plots of different size (40 × 40 cm and 80 × 80 cm), both inside and at the edge of the P. oceanica meadow, according to an orthogonal multifactorial design. In November 2011 (16 months after the start of the experiment), no C. racemosa was found inside the seagrass meadow, while, at the edge, the cover of the seaweed was dependent on disturbance intensity, being greater where the rhizomes had been damaged (H) than in leaf removal (L) or undisturbed (C) plots. The results of this study indicate that physical disturbance at the margin of seagrass meadows can promote the spread of C. racemosa.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Several factors have been identified as pivotal in determining the establishment and spread of non-native invasive species (see reviews by Airoldi 1998; Richardson et al. 2000; Dietz and Edwards 2006; Theoharides and Dukes 2007). Because the diversity of extant native communities and structured habitats has been considered as an important barrier for invasion (Byers 2002; Predick and Turner 2008), anthropogenic disturbance is considered as an invasion promoter (Richardson et al. 2000; Bulleri et al. 2010). Removal of biomass due to disturbance can favour the establishment of nonindigenous species by reducing competition from natives and/or by enhancing resource availability (Davis et al. 2000; Mack et al. 2000; Johnston and Keough 2002; Shea and Chesson 2002). Although this disturbance-invasion paradigm is not always supported by empirical studies (e.g. Lohrer et al. 2008), correlative and manipulative studies have extensively shown a positive relationship between ecosystem invasibility and disturbance, both in terrestrial and marine ecosystems (Hobbs and Huenneke 1992; D’Antonio and Vitousek 1992; Clark and Johnston 2011; Tamburello et al. 2014).
In marine environments, habitat-forming macrophytes regulate ecosystem processes by dominating primary production (Paine 2002), increasing habitat complexity and modifying the environment for associated organisms (Dayton 1975; Bruno et al. 2003). The endemic seagrass Posidonia oceanica (L.) Delile is a key species in Mediterranean coastal ecosystems (Pergent 1992), experiencing a widespread decline throughout the basin (Pérès 1984; Boudouresque et al. 1990; Peirano and Bianchi 1997; Boudouresque 2003). The regression of seagrass beds and the consequent expansion of alternative habitats (e.g. algal turfs or dead seagrass rhizomes generally referred to as “dead matte”) have been documented adjacent to urban areas worldwide (Boudouresque et al. 2009; Waycott et al. 2009; Montefalcone et al. 2010; Tamburello et al. 2012). Among the various human activities responsible for seagrass regression, direct mechanical damage from various fishing activities (e.g. trawling, gillnetting), anchoring (Ardizzone et al. 2006; Ceccherelli et al. 2007; Montefalcone et al. 2008) and dredging can cause physical removal of seagrass rhizomes and shoots, reducing habitat complexity and, thus, species diversity. Introduced species have also often been implicated in seagrass declines, but the evidence for negative effects on seagrasses is largely correlative (Glasby 2013 for a review), and in general, invasions by macroalgae occur in seagrass meadows already exposed to human perturbations (Bulleri et al. 2011). Given the limited experimental evidence, it thus remains unclear if macroalgal invasions are “drivers” of seagrass declines or instead “passengers” (Bulleri et al. 2010) that are responding opportunistically to degradation of seagrass beds caused by other disturbances (Williams 2007; Glasby 2013).
Species of the genus Caulerpa are the best known macroalgal invaders of seagrass beds (Williams and Smith 2007). Caulerpa racemosa is widely spread in the northwestern Mediterranean, where it has established in different habitats, at depths ranging from 0 to 40 m (Ceccherelli et al. 2002; Piazzi et al. 2005; Ruitton et al. 2005; Piazzi and Balata 2009). Correlative evidence suggests that P. oceanica meadows fragmented or characterized by low shoot densities are more heavily colonised by C. racemosa (Katsanevakis et al. 2010; Bulleri et al. 2011). In particular, Katsanevakis et al. (2010) found a low occurrence of C. racemosa inside intact meadows, with the macroalga mostly restricted to the margins of seagrass meadows or to gaps within disturbed meadows. In addition, experimental evidence indicates that the growth of C. racemosa at the edge of seagrass meadows is regulated by P. oceanica: stolon elongation and percent cover of C. racemosa were significantly higher where P. oceanica shoot density was reduced to 10 % of the natural density (Ceccherelli et al. 2000).
Thus, disturbance to seagrass shoots due to anchoring or fishing activities (trawling, gillnetting) could promote the establishment and/or spread of C. racemosa into seagrass beds (Francour et al. 1999; Milazzo et al. 2004; Ceccherelli et al. 2007; Montefalcone et al. 2007). Gaps in P. oceanica beds are likely to create favorable microhabitats for the settlement and spread of C. racemosa (Kiparissis et al. 2011). C. racemosa is, in fact, able to colonize space through both vegetative growth of prostrate stolons and settlement of current-borne fragments (Piazzi and Cinelli 1999). However, while several descriptive studies have documented the presence of C. racemosa in degraded P. oceanica habitats (Katsanevakis et al. 2010; Bulleri et al. 2011), except for the study of Ceccherelli et al. (2000), there has been no experimental investigation of how mechanical disturbance can promote the spread of this macroalga into P. oceanica meadows.
The aim of this study was to estimate whether the spread of C. racemosa depends on the intensity of disturbance on the canopy structure of the seagrass and the location of the disturbance within the seagrass bed. We mimicked the damage provoked by mechanical human disturbance on the seagrass canopy structure by inflicting different disturbance intensities (rhizome damage, leaf removal and undisturbed) to areas of different size and thus simulating damage due to human activities, such as anchoring or gillnet operation, of variable extent. C. racemosa can colonize space at the edge of meadows through both vegetative propagation from invaded area at the margins and supply of fragments from the water column. Differently, only the latter mechanism can promote space colonization by C. racemosa inside meadows. Thus, the different combinations of intensity and extent of disturbance were applied inside and at the edge of the meadow. Specifically, we tested the hypotheses that the spread of C. racemosa would be greater (1) under high intensity of disturbance, (2) in larger disturbed patches and (3) at the edge rather than inside the meadow.
Materials and methods
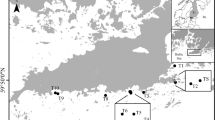
The study was done in the Asinara Gulf (northwestern tip of Sardinia, 40° 51.878N and 8° 20.290E), in the area between the town of Porto Torres and the Asinara Island MPA (Fig. 1). At our study site, continuous P. oceanica meadows occurred from shallow waters to about 40 m of depth. Canopy structure of the seagrass in this area seemed relatively well preserved, with shoot densities of 176.4 ± 14.2 and 237.0 ± 20.0 shoots.m−2 (mean ± SE; n = 48) in the inner meadow and at the edge, respectively (the difference in structure is due the presence of orthotropic and plagiotropic rhizomes at the edge of the meadow; Hemminga and Duarte 2000). Leaf lengths (see Results) corresponded generally with those reported from areas not exposed to major human disturbance (Pergent et al. 1995). Rhizomes of the plants are mostly orthotropic, and areas of dead matte were not common. At this site, C. racemosa has been spreading for about 15 years, and at the time of the study, it had become common on rocky reefs, on dead matte of P. oceanica and at the margin of meadows (Bulleri et al. 2011). The presence of the macroalga in discontinuities of the meadow, probably caused by mechanical damage from illegal trawling, was quite common (authors’ personal observation).

Study site in the Asinara Gulf (NW Sardinia)
A field experiment was started in July 2010, when habitat complexity of a P. oceanica bed, about 5 m deep, was manipulated to simulate the effects of mechanical disturbances of different intensity: rhizome damage (High disturbance intensity = H), leaf removal (Low disturbance intensity = L), and undisturbed (Control = C). The high disturbance intensity consisted in the damaging of rhizomes through the cutting of all the shoots under the basal meristem (not allowing leaf re-growth), while the low level was obtained by cutting shoots over the basal meristem (allowing leaf re-growth). Disturbance was applied at two positions of the seagrass meadow: inside and at the edge of the meadow. For the latter position, one side of each plot was open to the adjacent rocky platform, which had already been invaded by C. racemosa (site-wide cover in July 2010 was 58.7 ± 9.1 % (mean ± SE), based on 25 haphazardly-placed 400 cm2 quadrats). Colonization of plots at the edge could, thus, occur directly by clonal spread from the adjacent area. We did not, however, measure the abundance or distribution of C. racemosa in the area adjacent to each individual plot, and unfortunately, due to the sampling frequency of our experiment, we could not determine if the colonization of any individual plot was due to clonal growth or the settlement of fragments. Disturbance treatments were applied to plots (i.e., the experimental units) of different size (small 40 × 40 cm and large 80 × 80 cm), according to an orthogonal multifactorial design. Plots were interspersed within the same P. oceanica bed: edge plots followed the contour of the bed, while plots inside the bed were established about 5 m from the edge. Three replicate plots were established for each combination of position, plot size and intensity of disturbance. There was no C. racemosa in any of the plots at the beginning of the study.
Leaf length in the low disturbance plots was quantified at three different times (July 2010, about 3 weeks after the start of the experiment, November 2010 and May 2011), by measuring the longest leaf of five shoots chosen at random within each experimental unit (=plot). Data taken on the last time were analysed by four-way ANOVA including the factors ‘position’ (edge and inside), ‘size’ of the plots (small and large), ‘intensity’ of disturbance (H, L, and C), and ‘experimental unit’ (nested in the interaction ‘position × size × disturbance’). No regrowth of the leaves was observed where the meristem of the rhizome was damaged (H).
In order to obtain a comprehensive assessment of the response of C. racemosa to experimental conditions, its cover was quantified both on the bottom of plots and on P. oceanica rhizomes. The cover of C. racemosa on the bottom of each plot was quantified at four different times (August, October 2010, May and November 2011), by photographing the sedimentary substrate (25 × 25 cm surface). The cover of the C. racemosa on rhizomes was estimated at three different times (November 2010, May and December 2011) by photographing five randomly-selected rhizomes for each experimental plot. Photographs were taken using a digital camera (Canon S70) equipped with a waterproof case (Canon WPDC40) and a metal spacer and framer. The percentage cover of C. racemosa on the substrate was estimated visually on a computer screen: a grid of twenty-five sub-quadrats was superimposed onto each image and a score from 0 to 4 % was given in each sub-quadrat, and the percentage cover estimated by summing over the entire set of sub-quadrats (Dethier et al. 1993). Similarly, the percentage cover of C. racemosa on the rhizomes was estimated by using a vertical rectangular grid of twenty sub-quadrats (2 × 10) giving a score from 0 to 5 % to each sub-quadrat, adapting the grid to the rhizome size. Rhizomes shorter than 3 cm were not included in the sampling.
C. racemosa did not colonize plots inside the meadows. Analyses on the abundance of the seaweed were, therefore, restricted to data from the edge. The cover of C. racemosa on the substrate and on rhizomes was analysed by means of a two-way ANOVA including the factors ‘size of the plot’ (small and large) and ‘intensity’ of disturbance (H, L, and C), treated as fixed and orthogonal. Cochran’s test was performed to check for homogeneity of variances, while SNK test was utilized to make a posteriori comparisons of mean values (Underwood 1997).
Results
The structure of P. oceanica canopy was affected by disturbance treatments, although there was large variability among plots. In plots exposed to high intensity of disturbance, no re-growth was observed during the study period after the initial removal of all leaves in July 2010 (Fig. 2). Although the recovery of the plants exposed to a low intensity of disturbance was appreciable, the height of the canopy was lower than in control plots, independent of the size of the plot (significant Size × Intensity and SNK test in Table 1; Fig. 2).

Mean (+SE) P. oceanica canopy height (cm) at the edge and inside the meadow at the three sampling times (July–November 2010 and May 2011) for the three disturbance levels: High intensity (H), Low (L) and Controls (C) in the small (40 × 40 cm) and large (80 × 80 cm) plots. The last sampling time has been used for the analysis
C. racemosa was never found in plots inside the meadow, either on the substrate or on P. oceanica rhizomes. In contrast, at the edge of the meadow, C. racemosa colonized all the plots, although its abundance on the substrate and rhizomes depended on the intensity of disturbance (Table 2; Fig. 3): at the end of the study, the cover of C. racemosa on the substrate in plots exposed to high disturbance intensity was greater than in those exposed to low disturbance or un-disturbed controls (SNK tests in Table 2): graphical inspection suggests that, through time, the re-growth of the canopy reduced the abundance of C. racemosa in low disturbance intensity plots. The size of the plot did not affect the cover of C. racemosa on the substrate, but it did influence the occurrence on P. oceanica rhizomes, although cover values were generally low (Table 2). C. racemosa cover on rhizomes was, in fact, appreciable only in large, highly disturbed plots at the edge of the meadow (significant Size × Intensity and SNK test in Table 2).

Mean (±SE) C. racemosa percent cover through the study period on the substrate (left) and on the rhizomes (right) at the edge of the meadow for the High intensity (H), Low intensity (L), and Controls (C) in the small (40 × 40 cm) and large (80 × 80 cm) plots. The last sampling time has been used for the analysis. C. racemosa cover inside the meadow was zero and not plotted
Discussion
In NW Sardinia, patterns of occurrence of C. racemosa among areas subjected to different human influences have been found to match variations in the relative proportion of favourable and unfavourable habitats (Bulleri et al. 2011). Degraded habitats, such as dead P. oceanica rhizomes or macroalgal turfs, are more vulnerable to be colonized by C. racemosa, while structured habitats, such as seagrass beds and stands of canopy-forming macroalgae, can act as a barrier against the spread of this introduced seaweed (Bulleri et al. 2010). The experimental results of this study suggest that physical disturbance clearing the canopy promotes the spread of C. racemosa at the margin of seagrass meadows. Our prediction that invasion success of C. racemosa would have been greater under a higher intensity of disturbance was confirmed.
In particular, the marked re-growth of leaves in low-disturbance plots, both small and large in size, can thus reduce the spread and reattachment of fragments of C. racemosa. C. racemosa was able to spread rapidly into low-disturbance plots shortly after the beginning of the experiment, when the canopy had not re-grown yet, indicating a 3-month response of the macroalga to the availability of free space. However, over time, the cover of the macroalga in low-disturbance plots declined and was not different from controls by the end of the experiment. In contrast, in high-disturbance plots C. racemosa cover was initially much lower but increased steadily during the course of the experiment. Thus, the availability of free space is apparently not the only factor driving initial colonization rates since high and low disturbance intensity should have then supported similar covers of C. racemosa shortly after disturbance treatments were applied (August 2010). Nevertheless, a clear effect of the disturbance intensity emerged by the end of the experiment (November 2011), although the temporal trend among treatments seemed different especially until October 2010. In all treatments, the cover of C. racemosa decreased in winter and spring 2011 and then increased through the summer until November 2011, reflecting seasonal effects on its growth dynamics in this region of the Mediterranean (e.g. Ceccherelli et al. 2000; Piazzi et al. 2001).
Several mechanisms can be responsible for C. racemosa failing to spread into undisturbed and low disturbance intensity of P. oceanica canopy. The presence of the canopy can directly or indirectly affect the environment underneath it and influence the invasion success. For example, sweeping of the substrate by leaves, may prevent the establishment of understory species, as documented by Irving and Connell (2006) in kelp forests. Alternatively, reduced C. racemosa spread might be the result of a reduction in irradiance under the canopy. Whatever the mechanism, the results of this study provide support to previous correlative and manipulative evidence of a better ability of C. racemosa to colonize poorly structured seagrass architecture: vertical and horizontal C. racemosa growth is negatively influenced by dense and healthy P. oceanica meadows (Ceccherelli et al. 2000; Klein and Verlaque 2008; Katsanevakis et al. 2010). In contrast, in fragmented seagrass meadows, sand or matte patches are vulnerable to C. racemosa invasion (Katsanevakis et al. 2010; Bulleri et al. 2011; Kiparissis et al. 2011). In addition, this seaweed has often been found growing along the rhizomes inside sparse meadows (Klein and Verlaque 2008).
The physical and biological characteristics of a local patch of habitat and of the surrounding landscape can both be important determinants of habitat occupancy (Pearson 1993; Wiens et al. 1993; Ricketts 2001; Thomas et al. 2001; Driscoll 2007). Thus, the structure of P. oceanica canopy in the experimental plots may not be the only factor regulating establishment and spread of C. racemosa. The neighbour habitat and the seascape may change the probability for a seagrass patch of poor structure to be colonised by preventing species spread. However, the results of this study suggest that gaps of disturbed P. oceanica inside the meadow would not enhance the establishment of C. racemosa. Indeed, inside the meadow, C. racemosa was found neither in large nor in small size gaps created with high disturbance intensity suggesting that this type of localized and sparse clearances do not promote the establishment of the macroalga. Although ‘uprooted’ fragments of C. racemosa can easily re-attach to the substrate (Ceccherelli and Piazzi 2001; Piazzi et al. 2003), we believe that in such gaps the buffer effect of the surrounding canopy can contrast or delay the arrival of fragments that, being not-buoyant (Ceccherelli and Piazzi 2001), have reduced chances to reach such gaps. Thus, only a small proportion of drifting fragments transported by waves or currents reach these gaps, as they are likely to be retained by seagrass rhizomes at the margin of the meadow. However, at the edge of the meadow, we were not able to tease apart the relative contribution of the supply of fragments from the water column from that of clonal growth in determining colonization success of C. racemosa. However, the clonal growth habit of C. racemosa and its high abundance in areas adjacent to the meadow edge (>50 % cover) makes it likely that the greater colonization of disturbed edge plots was due to clonal growth of established individuals. Further experiments are required to isolate the relative contribution of the two mechanisms that potentially account for colonization of space at the edge of seagrass meadows. These experiments would imply the removal of C. racemosa in the area surrounding disturbed P. oceanica plots, so that recorded colonization could be reliably ascribed to the settlement of algal fragments from the water column.
Further, for the spread of the macroalga on rhizomes, the size of disturbance seems as important as the intensity of disturbance, highlighting that the larger the size the greater would be the invasion. This supports the hypothesis that the degree of the invasion of the seagrass might also change depending on the extension of disturbance and would be more rapid when disturbance is distributed over large areas. Although analyses of long-term invasion dynamics in disturbed seagrass habitats are lacking, our results suggest that C. racemosa is a passenger of changes in P. oceanica habitat rather than a driver, supporting previous experimental evidence from rocky reefs (Bulleri et al. 2010). However, this is not necessarily in opposition to evidences that the level of interaction between P. oceanica and C. racemosa significantly modifies the vegetative development of the seagrass (Dumay et al. 2002) and that the withdrawal of P. oceanica from the disturbed areas that have been colonized by C. racemosa may be permanent, due to high colonization potential of the macroalga and its phytotoxic properties (Kiparissis et al. 2011 and reference therein).
In conclusion, although the effect of seascape structure on invasion biology of the macroalga is still to be defined, disturbed P. oceanica patches enhance the occupancy and the spread of C. racemosa. Mechanical human disturbance of P. oceanica is weakly managed, as poaching occurs and measures of surveillance are inadequate. As a consequence, damage due to trawling is relatively common, while use of gillnets or anchoring is managed locally only for the specific objectives of marine protected areas. According to our results, management measures for protecting seagrass meadows and the biodiversity they host from mechanical disturbance would indirectly hinder the spread of C. racemosa. Determining which management scenario will best control invasive species will depend on their relative efficacy in protecting vulnerable habitats and raising of public awareness.
References
Airoldi L (1998) Roles of disturbance, sediment stress, and substratum retention on spatial dominance in algal turf. Ecology 79:2759–2770
Ardizzone G, Belluscio A, Maiorano L (2006) Long-term change in the structure of a Posidonia oceanica landscape and its reference for a monitoring plan. Mar Ecol 27:299–309
Boudouresque CF (2003) The erosion of Mediterranean biodiversity. In: Rodriguez-Prieto C, Pardini G (eds) The Mediterranean sea: an overview of its present state and plans for future protection. Servei de Publicacions de la Universitat de Girona, Girona, pp 53–112
Boudouresque CF, Ballesteros E, Ben Maiz N, Boisset F, Bouladier E, Cinelli F, Cirik S, Cormaci M, Jeudy De Grissac A, Laborel J, Lanfranco E, Lundberg B, Mayhoub H, Meisnez A, Panayotidis P, Semroud R, Sinnassamy JM, Span A, Vuignier G (1990) Livre rouge “Gérard Vuigner” des végétaux, peuplements et paysages marins menaces de Méditerranée. PNUE Publications, Athens 250
Boudouresque CF, Bernard G, Pergent G, Shili A, Verlaque M (2009) Regression of Mediterranean seagrasses caused by natural processes and anthropogenic disturbances and stress: a critical review. Bot Mar 52(5):395–418
Bruno JF, Stachowicz JJ, Bertness MD (2003) Inclusion of facilitation into ecological theory. Trends Ecol Evol 18:19–125
Bulleri F, Balata D, Bertocci I, Tamburello L, Benedetti-Cecchi L (2010) The seaweed Caulerpa racemosa on Mediterranean rocky reefs: from passenger to driver of ecological change. Ecology 91:2205–2212
Bulleri F, Alestra T, Ceccherelli G, Tamburello L, Pinna S, Sechi N, Benedetti-Cecchi L (2011) Determinants of Caulerpa racemosa distribution in the north-western Mediterranean. Mar Ecol Prog Ser 431:55–67
Byers JE (2002) Physical habitat attribute mediates biotic resistance to non-indigenous species invasion. Oecologia 130:146–156
Ceccherelli G, Piazzi L (2001) Dispersal of Caulerpa racemosa fragments in the Mediterranean: lack of detachment time effect on establishment. Bot Mar 44:209–213
Ceccherelli G, Piazzi L, Cinelli F (2000) Response of the non-indigenous Caulerpa racemosa (Forsskal) J-Agardh to the native seagrass Posidonia oceanica (L.) Delile: effect of density of shoots and orientation of edges of meadows. J Exp Mar Biol Ecol 243:227–240
Ceccherelli G, Piazzi L, Balata D (2002) Spread of introduced Caulerpa species in macroalgal habitats. J Exp Mar Biol Ecol 280:1–11
Ceccherelli G, Campo D, Milazzo M (2007) Short-term response of the slow growing seagrass Posidonia oceanica to simulated anchor impact. Mar Environ Res 63:341–349
Clark GF, Johnston EL (2011) Temporal change in the diversity–invasibility relationship in the presence of a disturbance regime. Ecol Lett 14:52–57
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass: fire cycle, and global change. Ann Rev Ecol Evol S 23:63–87
Davis MA, Grime JP, Thompson K (2000) Fluctuating resources in plant communities: a general theory of invasibility. J Ecol 88:524–534
Dayton PK (1975) Experimental evaluation of ecological dominance in a rocky intertidal community. Ecol Monogr 45:137–159
Dethier MN, Graham ES, Cohen S, Tear LM (1993) Visual versus random-point percent cover estimations: ‘objective’ is not always better. Mar Ecol Progr Ser 110:9–18
Dietz H, Edwards PJ (2006) Recognition that causal processes change during plant invasion helps explain conflicts in evidence. Ecology 87:1359–1367
Driscoll DA (2007) How to find a metapopulation. Can J Zool 85:1031–1048
Dumay O, Pergent G, Pergent-Martini C, Amade P (2002) Variations in caulerpenyne contents in Caulerpa taxifolia and Caulerpa racemosa. J Chem Ecol 28(2):343–352
Francour P, Ganteaume A, Poulain M (1999) Effect of boat anchoring in Posidonia oceanica seagrass beds in the Port-Cros national park (north-western Mediterranean sea). Aquat Conserv 9:391–400
Glasby TM (2013) Caulerpa taxifolia in seagrass meadows: killer or opportunistic weed? Biol Invas 15:1017–1035
Hemminga MA, Duarte CM (2000) Seagrass ecology. Cambridge University Press, Cambridge
Hobbs RT, Huenneke LF (1992) Disturbance, diversity, and invasion: implications for conservation. Conserv Biol 6:324–337
Irving AD, Connell SD (2006) Predicting understorey structure from the presence and composition of canopies: an assembly rule for marine algae. Oecologia 148:491–502
Johnston EL, Keough MJ (2002) Direct and indirect effects of repeated pollution events on marine hard-substrate assemblages. Ecol Appl 12:1212–1228
Katsanevakis S, Issaris Y, Poursanidis D, Thessalou-Legaki M (2010) Vulnerability of marine habitats to the invasive green alga Caulerpa racemosa var. cylindracea within a marine protected area. Mar Env Res 70:210–218
Kiparissis S, Fakiris E, Papatheodorou G, Geraga M, Kornaros M, Kapareliotis A, Ferentinos G (2011) Illegal trawling and induced invasive algal spread as collaborative factors in a Posidonia oceanica meadow degradation. Biol Invas 13:669–678
Klein J, Verlaque M (2008) The Caulerpa racemosa invasion: a critical review. Mar Poll Bull 56:645–650
Lohrer AM, Chiaroni LD, Hewitt JE, Thrush SF (2008) Biogenic disturbance determines invasion success in a subtidal soft-sediment system. Ecology 89:1299–1307
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M et al (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl 10:689–710
Milazzo M, Badalamenti F, Ceccherelli G, Chemello R (2004) Boat anchoring on Posidonia oceanica beds in a marine protected area (Italy, western Mediterranean): effect of anchor types in different anchoring stages. J Exp Mar Biol Ecol 299:51–62
Montefalcone M, Morri C, Peirano A, Albertelli G, Bianchi CN (2007) Substitution and phase shift within the Posidonia oceanica seagrass meadows of NW Mediterranean Sea. Estuar Coast Shelf Sci 75:63–71
Montefalcone M, Chiantore M, Lanzone A, Morri C, Albertelli G, Bianchi CN (2008) BACI design reveals the decline of the seagrass Posidonia oceanica induced by anchoring. Mar Poll Bull 56(9):1637–1645
Montefalcone M, Albertelli G, Morri C, Bianchi CN (2010) Patterns of wide-scale substitution within meadows of the seagrass Posidonia oceanica in NW Mediterranean Sea: invaders are stronger than natives. Aquat Conserv 20(5):507–515
Paine RT (2002) Trophic control of production in a rocky intertidal community. Science 296:736–739
Pearson SM (1993) The spatial extent and relative influence of landscape-level factors on wintering bird populations. Landsc Ecol 8:3–18
Peirano A, Bianchi CN (1997) Decline of the seagrass Posidonia oceanica in response to environmental disturbance: a simulation-like approach off Liguria (NW Mediterranean Sea). In: Hawkins LE, Hutchinson S (eds) The response of Marine organisms to their environments. University of Southampton, UK, pp 87–95
Pérès JM (1984) La regression des herbiers à Posidonia oceanica. In: Boudouresque CF, Jeudy De Grissac A, Olivier J (eds) First international workshop on Posidonia oceanica beds. GIS Posidonie Publ, France, pp 445–454
Pergent G (1992) Key species in the Mediterranean littoral. The importance of Posidonia oceanica meadows. The European coastline study. European Science Foundation and Commission of the European Communities, Galway, pp 32–36
Pergent G, Pergent-Martini C, Boudouresque CF (1995) Utilisation de l’herbier à Posidonia oceanica comme indicateur biologique de la qualité du milieu littoral en Méditerranée: Etat des connaissances. Mésogée 54:3–29
Piazzi L, Balata D (2009) Invasion of alien macroalgae in different Mediterranean habitats. Biol Inv 11:193–204
Piazzi L, Cinelli F (1999) Development and seasonal dynamics of a population of the tropical alga Caulerparacemosa (Forsskål) J Agardh in the Mediterranean. Cryptogam Algol 20:295–300
Piazzi L, Ceccherelli G, Cinelli F (2001) Threat to macroalgal diversity: effects of the introduced green alga Caulerpa racemosa in the Mediterranean. Mar Ecol Progr Ser 210:149–159
Piazzi L, Ceccherelli G, Balata D, Cinelli F (2003) Early patterns of Caulerpa racemosa in the Mediterranean Sea: the influence of algal turfs. J Mar Biol Assoc UK 83:27–29
Piazzi L, Meinesz A, Verlaque M, Akçali B, Antolić B, Argyrou M, Balata D, Ballesteros E, Calvo S, Cinelli F, Cirik S, Cossu A, D’Archino R, Djellouli AS, Javel J, Lanfranco E, Mifsud C, Pala D, Panayotidis P, Peirano A, Pergent G, Petrocelli A, Ruitton S, Žuljević A, Ceccherelli G (2005) Invasion of Caulerpa racemosa var. cylindracea (Caulerpales, Chlorophyta) in the Mediterranean Sea: an assessment of the spread. Cryptogam Algol 26:189–202
Predick KI, Turner MG (2008) Landscape configuration and flood frequency influence invasive shrubs in floodplain forests of the Wisconsin River (USA). J Ecol 96:91–102
Richardson DM, Holmes TM, Esler KJ, Galatowitsch SM et al (2000) Riparian vegetation degradation, alien plant invasions, and restoration prospects. Divers Distrib 13:126–139
Ricketts TH (2001) The matrix matters: effective isolation in fragmented landscapes. Am Nat 158:87–99
Ruitton S, Javel F, Culioli JM, Meinesz A, Pergent G, Verlaque M (2005) First assessment of the Caulerpa racemosa (Caulerpales, Chlorophyta) invasion along the French Mediterranean coast. Mar Pollut Bull 50:1061–1068
Shea K, Chesson P (2002) Community ecology theory as a framework for biological invasions. Trends Ecol Evol 17:170–176
Tamburello L, Benedetti-Cecchi L, Ghedini G, Alestra T, Bulleri F (2012) Variation in the structure of subtidal landscapes in the NW Mediterranean Sea. Mar Ecol Progr Ser 457:29–41
Tamburello L, Bulleri F, Balata D, Benedetti-Cecchi L (2014) The role of overgrazing and anthropogenic disturbance in shaping spatial patterns of distribution of an invasive seaweed. J Appl Ecol. doi:10.1111/1365-2664.12199
Theoharides KA, Dukes JS (2007) Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion. New Phytol 176:256–273
Thomas JA, Bourn NAD, Clarke RT, Stewart KE, Simcox DJ, Pearman GS, Curtis R, Goodger B (2001) The quality and isolation of habitat patches both determine where butterflies persist in fragmented landscapes. Proc R Soc Lond Ser B Biol Sci 268:1791–1796
Underwood AJ (1997) Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University Press, Cambridge
Waycott M, Duarte CM, Carruthers TJB, Orth RJ, Dennison WC, Olyarnik S, Calladine A, Forqurean JW, Heck KL, Hughes AR, Kendrick GA, Kenworthy WJ, Short FT, Williams SL (2009) Accelerating loss of seagrasses across the globe threatens coastal ecosystems. P Natl Acad Sci USA 106(30):12377–12381
Wiens JA, Stenseth NC, Van Horne B, Ims RA (1993) Ecological mechanisms and landscape ecology. Oikos 66:369–380
Williams SL (2007) Introduced species in seagrass ecosystems: status and concerns. J Exp Mar Biol Ecol 350:89–110
Williams SL, Smith JE (2007) A global review of the distribution, taxonomy and impacts of introduced seaweeds. Annu Rev Ecol Syst 38:327–359
Acknowledgments
We sincerely thank Francesco Mura for help with fieldwork and boating operations. S.P. was supported by funds from a “Regione Autonoma della Sardegna” (PO Sardegna FSE 2007-2013 LR7/2007) Grant. Two anonymous reviewers provided comments and constructive criticism on earlier draft.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ceccherelli, G., Pinna, S., Cusseddu, V. et al. The role of disturbance in promoting the spread of the invasive seaweed Caulerpa racemosa in seagrass meadows. Biol Invasions 16, 2737–2745 (2014). https://doi.org/10.1007/s10530-014-0700-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-014-0700-7