
Abstract
Non-native tree invasions occur not only in woodland or forest vegetation, but also into areas with little or no native tree presence. Limiting factors for tree establishment and survival include seasonal or annual drought, low nutrient availability, cold temperature extremes, fire, and other abiotic conditions to which trees are poorly adapted as well as biotic conditions such as herbivory and lack of soil mutualist inoculum. Tree invasions of grasslands and semi-arid riparian areas in particular are now widespread and frequently result in the rapid conversion of these habitats to woodlands or forests. In some cases, these invasions are the result of a change in extrinsic conditions such as climate, fire, and/or grazing that remove what have been previous barriers to tree establishment. However, in other cases, tree species with particular life-history and dispersal traits fill open niches or outcompete native species. Significant examples of tree invasion into treeless areas can be seen with invasions of Pinus species into temperate grasslands and fynbos shrublands, Melaleuca quinquenervia and Triadica sebifera into grassy wetlands, Prosopis and Tamarix species into semi-arid riparian zones, and Acacia and Morella invasions into nutrient-poor shrublands and barrens. The establishment of trees into treeless areas may have strong impacts on ecosystem processes, influencing biogeochemical cycling, carbon sequestration and cycling, and ecohydrology, as well possible edaphic legacies that persist even if trees are removed.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Native woody plant species are undergoing range expansions and increases in abundance in many parts of the world. At the same time, thousands of non-native woody plants have been moved outside their native ranges by humans and many species are spreading from sites of introduction in many regions. Both phenomena have profound consequences for community structure, biodiversity, and ecosystem functioning (Scholes and Archer 1997; Jackson et al. 2002; Browning et al. 2008; Richardson and Rejmánek 2011). Woody plants did not feature prominently on lists of the most important invasive alien species until fairly recently. However, dramatic invasions of many species of trees and shrubs in the last few decades around the world have ensured that woody plants are now among the most widespread and damaging of invasive organisms (Richardson and Rejmánek 2011).
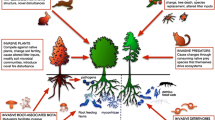
Many tree invasions occur in areas that were formerly treeless, or at least where native trees very seldom dominate vegetation formations. Extensive global areas of grasslands, shrublands, oligotrophic barrens, semi-arid lands, and alpine habitats possess limiting conditions that allow herbaceous or low woody vegetation to dominate and prevent the establishment/proliferation of trees (Fig. 1). Limiting factors for tree growth often include seasonal or annual drought, low nutrient availability, cold temperature extremes, fire, and/or other abiotic conditions to which trees are poorly adapted. Cycles of climate change, altered disturbance regimes or land-use histories frequently alter the conditions for tree recruitment, growth and survival, and in many cases change open treeless habitats to tree-dominated ecosystems (e.g. Richardson and Bond 1991; Pillar and Quadros 1997; Carilla and Grau 2010).

Conceptual model of limiting factors and releases from these limitations in the invasion of trees into grassland and shrubland ecosystems
Invasions of grasslands and the conversion of such systems to woodlands or forests represent some of the best documented examples of tree invasion into previously treeless areas. In many case studies around the world these invasions are the result of a change in extrinsic conditions such as climate, fire, and/or grazing that remove what have been previous barriers to tree establishment (Vale 1981; Richardson and Bond 1991; Scholes and Archer 1997; Rudel et al. 2005; Coop and Givnish 2008). However, while such intrinsic factors may promote colonization and seedling establishment, there are critical biotic interactions or positive feedbacks that act to maintain or accelerate tree recruitment and spread. These interactions occur because trees may modify their environments in ways that enhance further recruitment (Scholes and Archer 1997; Haugo and Halpern 2010). Nodes of colonization are often critical hotspots for invasion (Duarte et al. 2006). In particular, the colonization of grasslands by trees may have profound and irreversible impacts on primary production, plant allocation, rooting depth and soil faunal communities to depths of several meters. Increased abundance of trees may therefore have strong impacts on ecosystem processes such as biogeochemical cycling, carbon sequestration and cycling, and ecohydrology (Geesing et al. 2000; Guo and Gifford 2002; Jackson et al. 2002; Ehrenfeld 2003; Hibbard et al. 2003; Huxman et al. 2005; Dickie et al. 2011).
Native tree colonization and expansion in treeless areas
This successional process of tree establishment in treeless areas is best documented in native conifer invasions of mountain meadows in the western United States where species such as Pinus contorta have become increasingly established in high elevation meadows (Vale 1981; Miller and Halpern 1998; League and Veblen 2006). It is clear that extrinsic factors often trigger seedling establishment, and that positive interactions and density dependent feedbacks can regulate the pace and intensity of invasion even when climatic conditions are not optimal for establishment (Halpern et al. 2010; Rice et al. 2012). However, biotic interactions related to seed dispersal, phenology, and functional traits may strongly influence invasion success (Coop and Givnish 2008; Haugo and Halpern 2010; Halpern et al. 2010).
The success of the native pine colonization as indicated above suggests that tree growth forms are not precluded from mountain meadows if critical abiotic limitations are lifted and necessary biotic feedback takes place. It is not surprising therefore to imagine that non-native trees may also have the ability to colonize grassland habitats if they possess appropriate ecological, ecophysiological or phenological adaptations not present in the native tree flora and that obviate the critical abiotic limitations for seedling establishment (Richardson and Bond 1991). However, some caution is needed in such interpretations without a careful consideration of antecedent conditions that might have allowed grasslands to become dominant. For example, fire represents a potential abiotic limiting factor for tree survival that can be altered with changes in a fire regime (Mazia et al. 2001). Whatever these conditions, the colonizing trees can be seen to have an increased fitness over the grass species and can develop a shared dominance of microsites within a meadow.
Fitness advantage versus empty niches
Tree incursions into treeless areas can involve coexistence with existing herbaceous or shrub communities, as in the example of many tropical savannas. In other cases, tree invasions quickly lead to competitive exclusion of most native species as the stands of trees become established and shade out the existing vegetation. Such incursions/invasions open the question of whether invasive non-native trees have increased fitness over native species in treeless areas or instead are exploiting an open niche. Classic niche theory predicts that invaders with niches distinct from the native flora should coexist with little interaction with native species, yet successful invaders often have substantial impacts, suggesting that empty niches alone do not explain invasion success. MacDougall et al. (2009) argued that while successful invader establishment can be based on either a fitness advantage or niche difference from native species, only the former allows invaders to become dominant (Fig. 2). If this model is correct, it suggests that most examples of tree invasion into previously treeless areas involve elements of both superior fitness and existing empty niches.

The relative tradeoffs of fitness advantage versus niche differentiation in the invasion of tree species into treeless areas (adapted from MacDougall et al. 2009)
There are certainly examples of tree invasions into treeless areas that can be attributed solely to empty niches existing because of phylogenetic constraints within an existing flora (Mack 2003). Such phylogenetic constraints may leave arboreal niches open and available for successful colonization by pre-adapted non-native tree species. One of the often cited examples of such empty niches can be seen with oceanic islands where restricted dispersal limits the genetic diversity of colonizing lineages (Carlquist 1974). Trees are often poor dispersers and rarely good colonizers, leaving open niches in habitats otherwise favorable for tree growth. In evolutionary time such empty niches are filled to some extent by the evolution of tree growth forms within the colonizing lineages. This process can be seen well within the Hawaiian flora and in tropical alpine ecosystems. It is interesting to note that this process has not happened in some biodiversity hotspots, such as the Cape region of South Africa. There, despite remarkable radiations in many lineages, clades that have produced tree forms in other regions (e.g. in Ericaceae, Proteaceae, Rhamnaceae) have not done so in the Cape.
An example of an empty niche available for colonization by non-native arboreal species can be seen with mangroves in the Hawaiian Islands. Although many mangrove species are well adapted for ocean dispersal, the remote geographic position of the islands resulted in no natural colonization by mangroves. Rhizophora mangle was intentionally introduced to the Hawaiian archipelago to help stabilize shoreline erosion in quiet bays. This introduction was spectacularly successful in limiting erosion, but the accumulated sediments have altered the composition of the resident intertidal community and native birds that depend on shoreline invertebrates (Allen 1998). The invasion of quinine trees (Cinchona pubescens, Rubiaceae) into treeless upland areas of the Galapagos Islands presents another interesting example of the creation of a novel ecosystem (Jäger et al. 2007).
Tree invasions into temperate grasslands
With the exception of Pinus merkusii whose natural range barely extends south of the equator in Indonesia, pines (Pinus; Pinaceae) are restricted in their native range to the Northern Hemisphere. However, native grasslands and shrublands in many parts of the Southern Hemisphere are now invaded by pines introduced for forestry (Richardson and Higgins 1998; Simberloff et al. 2010). These invasions are widespread with Pinus taeda and P. elliottii in southern Brazil (Simberloff et al. 2010), Pinus halepensis and P. radiata in northern Argentina (Zalba and Villamil 2002), P. contorta in Chilean Patagonia, and P. contorta and P. nigra in montane grasslands of New Zealand. These invasions suggest that selected pines possess life history and ecophysiological traits that allow them to fill an empty niche in many Southern Hemisphere grasslands (Grotkopp et al. 2002).
In most cases, pines have been introduced to austral areas as a forestry crop or for erosion control, and then widely and often rapidly invaded surrounding grassland areas. Lodgepole pine (P. contorta) was introduced to New Zealand in about 1880 and has become the most vigorous non-native conifer, which has led to large areas of unwanted encroachment into montane grasslands (Ledgard 2001). Invasive trees compete with native species, and strongly impact the visual landscape, and affect land use values. Lodgepole pine spreads more vigorously than other introduced conifers as it cones earlier and regularly, and produces lightweight seeds capable of dispersal over long distances. Encroachment occurs most readily on ungrazed land with low vegetation density, and can spread long distances in windy areas. Its physiological tolerances allow lodgepole pine to grow successfully at sites as much as 200 m higher than the altitudinal limit for native mountain trees and persisted at even higher elevations as krummholz (Wardle 1985).
Similarly, P. contorta (lodgepole pine) has become an aggressive invader into many Patagonian grasslands (Peña et al. 2008; Richardson et al. 2008; Simberloff et al. 2010). This species was first planted in Chilean Patagonia for erosion control caused by historical fires and cattle farming. More recently planting has expanded over large areas for commercial purposes, with P. contorta now spreading from these plantations into natural areas. Invasion success is influenced by seed dispersal, with high seedling densities found close to the parent stand and these decreasing as distance from the seed source increases (Langdon et al. 2010).
The availability of compatible ectomycorrhizal symbionts can be an important limiting factor in pine invasion (e.g. Richardson et al. 2000; Nuñez et al. 2009) as well as pine expansion within its native range (Terwilliger and Pastor 1999; Ashkannejhad and Horton 2006; Wiemken and Boller 2006; Collier and Bidartondo 2009). Nonetheless, the rapid invasion of pines suggests that dispersal of mycorrhizal fungi is relatively rapid into grassland areas. Further, field studies have found that the presence of mycorrhizal inoculum in the soil at the edge of Patagonian pine plantations decreases linearly, with distances up as much as 1,000 m from the plantation edge (Salgado Salomón et al. 2011), rather than dropping off sharply as has been observed at the edge of Fagaceae forests (Dickie and Reich 2005; Dickie et al. 2012). It appears that pine invasions are facilitated by the co-invasion of compatible ectomycorrhizal fungi (Dickie et al. 2010). Two particular, closely-related fungal genera, Rhizopogon and Suillus, may play a critical role as pine-specific mycorrhizal fungi with long-distance dispersal via mycophagous animals (Ashkannejhad and Horton 2006; Wiemken and Boller 2006) and, in the case of Rhizopogon at least, high spore longevity in the soil spore bank (Bruns et al. 2009). Once these fungi arrive, dispersal by either introduced or native mammals may rapidly facilitate pine invasion (Dickie et al. 2010).
The need of ectomycorrhizal trees like pines for compatible fungi certainly contributed to the “lag-phase” of invasion (during which a species is present but does not spread; see Aikio et al. 2010) for pines in many parts of the Southern Hemisphere (Richardson et al. 1994). Initial plantings of pine in the Southern Hemisphere largely failed until mycorrhizal fungi were introduced (Marx 1991; Pringle et al. 2009). This raises the important question of whether tree species that are not currently invasive, such as Eucalyptus species in New Zealand, could become invasive if and when compatible ectomycorrhizal fungi arrive (Diez 2005).
Pines are not the only group of trees to invade dry temperate grasslands. Honey locust (Robinia pseudoacacia, Fabaceae) is one of the most problematic invasive tree species in Europe (Kleinbauer et al. 2010). Although originally introduced for erosion control, agroforestry, and ornamental use, it has spread rapidly into treeless grassland areas.
Tree invasions into grassy wetlands
Grassy wetlands form another herbaceous community that has been successfully invaded by non-native trees in many parts of the world. Native trees are generally not able to tolerate such habitats due to the perennially or seasonally saturated soils. Many of the seasonally saturated wetlands present extreme physiological drought in the dry season, as well as conditions that promote frequent fire, adding to limitations on tree establishment.
One of the most problematic wetland invaders in the southeastern United States is Melaleuca quinquenervia (Myrtaceae), the paper-bark tree. This tree is native to wetland areas along the eastern coast of Australia and was introduced into southern Florida in 1886, initially as an ornamental and later for erosion control and as a windrow tree (Bodel et al. 1994). It has now invaded and become dominant over extensive natural areas of sawgrass wetlands, pine flatwoods, pastures, and cypress forests. Its dominance in these areas results from its broad tolerance of variable water regimes, moderate salinity, and strong seasonal shifts in temperature. Once established, M. quinquenervia has a high growth rate and tremendous reproductive potential at an early stage of growth. Essential oils in the canopies of these trees are highly flammable, promoting fires that kill the native species while promoting the germination of soil seed pools. The result is a virtual monoculture, often in areas that had formerly been grassy wetlands (Serbesoff-King 2003). Once established, stands of M. quinquenervia significantly alter both above- and belowground ecosystem components of carbon allocation and soil nutrient availability, producing a novel ecosystem structure not formerly present and which promotes its continued stand dominance (Bodel et al. 1994; Martin et al. 2009).
Another dramatic example of wetland tree invasion is that of the Triadica sebifera (Euphorbiaceae, formerly Sapium sebiferum), Chinese tallow tree. This native of subtropical China is now naturalized in many areas of the world, apparently only limited by climate extremes (Pattison and Mack 2008). It was introduced into the southern United States in the early twentieth century, and originally largely confined to wetland prairies which have now been converted from grassy wetlands to a monoculture of woodland cover over extensive areas (Bruce et al. 1995). This non-native woodland is persistent as a novel vegetation type. In contrast to M. quinquenervia which promotes fire, T. sebifera shades out flammable grasses which reduces the frequency of fire where it invades.
Tree invasions into fynbos
Tree invasions into South African fynbos, and to a lesser extent other Mediterranean-climate shrublands, are a fascinating example of the filling of a vacant niche. The absence of trees in fynbos is very puzzling and has been debated for decades. In 1936 Charles Darwin wrote in his journal about a fynbos landscape near Cape Town as follows: “There was not even a tree to break the monotonous uniformity of the sandstone hills: I never saw a much less interesting country…”. The absence of trees in fynbos is particularly surprising when one considers the widespread presence of woodlands in each of the other four mediterranean-climate regions of the world. The paradigm of old landscapes with low-nutrient soils (Hopper 2009) does not explain this pattern as evidenced by vegetation structure in environmentally similar areas of South Africa and Southwestern Australia. The Australian formations are generally an order of magnitude taller than the Cape for matched sites (Richardson and Cowling 1992). In Southwestern Australia vegetation changes from heathland formations similar to fynbos at low rainfall to woodlands and forests up to 18 times taller than fynbos at higher rainfall (Richardson and Cowling 1992).
As an historical note, observations on the rapid invasion of South African fynbos shrublands by introduced trees, leading to the transformation of these species-rich shrublands into species-poor woodlands or forests over a few decades (Fig. 3) were pivotal for launching the international SCOPE (Scientific Committee on Problems of the Environment) programme on the ecology and management of biological invasions in the mid 1980s. Hal Mooney, the driving force behind this program relates in his book The globalization of ecological thought (Mooney 2008), how the idea for the SCOPE program was conceived in Stellenbosch at the MEDECOS conference in 1980. He tells of his discussions with Fred Kruger (then Director of the South African Forestry Research Institute) about invasions of alien trees and shrubs in the fynbos-clad mountains around Stellenbosch. They felt that these invasions into relatively undisturbed ecosystems went against the notions set out in Charles Elton’s (1958) seminal book that invasions occurred only in human-modified systems. Mooney wrote “… [we] decided that perhaps another look at this issue [the ecosystem consequences of invasions] was warranted—maybe things had changed since Elton’s time”. The SCOPE program was born and was a major stimulus for focusing scientific attention on the problem of biological invasions (Simberloff 2011).

Schematic representation of changes in the structure of South African fynbos shrublands: a in the absence of introduced Pinus species (note the cyclical replacement process); b invaded by pines. In the absence of invasive pines, recruitment of overstorey shrubs is highly variable; after some fires they proliferate and suppress understorey species (a1), but sometimes post-fire recruitment is poor, resulting in local extinction of overstorey shrubs. Restoration of either condition requires colonization by components from the regional species pool (a2). The superior resilience under any fire regime of the alien pines disrupts the prevailing non-equilibrium system. As the pines proliferate (b2), competition with fynbos plants intensifies, eventually leading to their local extinction as seed banks are depleted. This results in a depauperate steady state. Restoration of the cyclical replacement of fynbos elements can only be achieved through mechanical removal of the pines. Figure modified from Richardson and Cowling (1992); see also Richardson and Gaertner (2013)
The evergreen shrubs of fynbos have similar photosynthetic capacities to shrubs in other mediterranean-type ecosystems but lack the capacity either as individuals or as a community to optimize resource use later in stand development (see Fig. 9.3 in Richardson and Cowling 1992). Annual biomass increments in the first few years after fire in fynbos approximate or slightly exceed those in California chaparral, but slow a few years after fire. The result is that non-native trees in fynbos can support a biomass five times that of native communities (Richardson and Cowling 1992).
Several groups of alien trees possess life-history and ecophysiological traits that enable them to infiltrate and rapidly transform ecosystem processes in fynbos, but different groups achieve invasion success in different ways (Grotkopp et al. 2002). As mentioned above, the classic example has been pine species (P. halepensis, P. pinaster and P. radiata) which were widely introduced as a source of wood. These species share the trait of having serotinous cones, and colonize fynbos after fires. As such, they initially behave much like the native shrubs, but their short juvenile periods and large reserves of highly mobile seeds buffer them against fire-induced population crashes. The prevailing non-equilibrium system in fynbos is disrupted, and cyclical replacement of native shrubs is curtailed. As the invading pines proliferate after each fire, competition with fynbos elements is intensified, eventually leading to the local extinction of native shrubs as residual seed stores are depleted. Cyclical replacement cannot be reinstated without human intervention (such as felling of pines) and a depauperate steady-state results (Fig. 3). This process has transformed vast tracts of fynbos to pine woodlands or forests, locking these communities into a tree-dominated state.
Hakea species (Proteaceae), despite having a very different evolutionary history to pines, also have serotinous follicles and wind-dispersed seeds. Together with other ecophysiological adaptations that allow them to persist and proliferate in the nutrient-poor and fire-prone fynbos environment, serotinous follicles allow them to perform very similarly to pines, achieving the same result—high levels of efficiency at occupying new sites and even greater efficiency at persisting at these sites under diverse fynbos fire regimes.
Another group of alien trees that has been spectacularly successful in invading and transforming fynbos are Australian acacias. Unlike pines and hakeas, the acacias do not retain seeds in the canopy, but accumulate massive seed stores in the soil. Acacias also differ from hakeas and pines in not showing adaptations for dispersal by wind, but rather are adapted for dispersal by animals (birds and ants). They also have the trait of symbiotic nitrogen fixation providing them an advantage for growth in nutrient-poor soils. Despite such fundamental differences in life-history strategy from hakeas and pines, acacias are able to replicate the effects shown in Fig. 3—transforming the prevailing non-equilibrium, cyclical replacement sequence that maintains and generates species richness and ecosystem function in fynbos to a depauperate species assemblage with radically changed ecosystem functions (Le Maitre et al. 2011).
Tree invasions into edaphic barrens
Invasions of tree species into edaphic barrens with oligotrophic soils often occurs with trees possessing nitrogen fixation associated with symbiotic root nodules. Such a trait is associated, for example, with the ability of alders to successfully become established in soils otherwise inhospitable for native tree species. Such an invasion by a non-native species can be seen in Morella faya, a tree native to Macaronesia, which has escaped from cultivation on Hawai’i and become widely established. Its colonization of relatively nutrient-poor volcanic soils is aided by the presence of symbiotic nitrogen fixation and a broad ability to grow under a variety of soils ranging from recent thin ash over lava to deep well-developed silty clay loam soil. Once established, soil nitrogen supply beneath stands of Morella increases by as much as 90 times over that of native sites, facilitating the invasion of non-native perennial grasses which promote fire (Vitousek and Walker 1989). These conditions produce a novel ecosystem structure dominated by non-native species where fire-sensitive native species compete poorly.
Tree invasions into arid and semi-arid riparian ecosystems
Tree spread or invasions into arid and semi-arid riparian ecosystems provide well-documented examples of how tree invasions can greatly alter existing conditions and have created novel ecosystems with new conditions of nutrient availability, water availability, soil salinity, and/or altered fire regime changes. These can be notably seen in the Southwestern United States where trees and woody shrubs, largely but not exclusively native, have invaded semi-arid regions and greatly expanded their density and biomass over the past two centuries (Hastings and Turner 1966; Archer et al. 1988; Archer 1989, 1994; Bahre 1991; Van Auken 2000; McClaran 2003). Although these invasions have been recognized for many years, there remains a poor understanding of mechanisms controlling the rates, dynamics and extent of this vegetation change. Similar patterns of woody plant encroachment into arid and semi-arid shrublands and savannas have been documented in other parts of the world. Traditional explanations have focused on changes in climate, livestock grazing and fire regimes as causal factors.
Among the important invasive trees in arid and semi-arid regions of the Southwestern United States are species of Prosopis (Fabaceae, mesquite), with the native P. glandulosa and P. velutina as major players in the encroachment of trees into arid and semi-arid grasslands and desert scrub. These two taxa plus the tropical taxa P. juliflora and P. palida have been widely introduced and become invasive in formerly treeless or open savanna areas in other semi-arid regions of the world (Kaur et al. 2012).
Multiple species of Prosopis were introduced into South Africa in the late 1880s to provide shade, fodder, and fuel wood in arid regions, and rapidly expanded their range through both intentional plantings and natural seed dispersal. Today, large areas of the arid Nama Karoo desert region have been invaded by Prosopis (Richardson and van Wilgen 2004). The invasions have been particularly successful in areas with deep alluvial soils and associated groundwater which the phreatophytic roots systems of Prosopis can tap. However, there is an impact of this water use as these are important aquifers for groundwater supply to farmers, livestock and rural settlements (Le Maitre et al. 2000). Some economic balance to this water use occurs, however, as Prosopis is an active nitrogen-fixer with high productivity (Rundel et al. 1982), making it valuable for fodder. Similar examples of extensive Prosopis encroachment into rangelands, with positive and negative effects, are documented for extensive areas of East Africa, the Middle East and India (Hailu et al. 2004; Kaur et al. 2012).
Species of Tamarix (saltcedar, tamarisk) were widely introduced to the Southwestern United States in the latter part of the nineteenth century for erosion control and as ornamental plants. These saltcedar rapidly expanded their range and by the 1960s were present along most rivers in the semiarid and arid parts of the region. The period of tamarisk invasion coincided with changing physical conditions along western rivers associated with the construction and operation of dams, and the associated changes in hydrological flow appear to have favored saltcedar over native riparian competitors like cottonwoods and willows (Glenn and Nagler 2005, Merritt and Poff 2010; Merritt and Shafroth 2012). Ecosystem impacts of riparian stand dominance by saltcedar include significant losses of water through evapotranspiration, salinization of upper soil levels, increased fire hazard, and loss of important wildlife habitat (Di Tomaso 1998; Shafroth et al. 2005). However, effective biological control of saltcedar has led a controversy over the costs versus benefits of this control (Stromberg et al. 2009). Elaeagnus angustifolia (Oleaceae, Russian olive) is a Eurasian tree introduced to the western United States and other semiarid regions for erosion control in the late nineteenth century. Once established, Russian olive outcompetes native vegetation, alters natural plant succession and nutrient cycling, and taxes soil water reserves. Today it has vastly expanded its range to form extensive monotypic stands along the watercourses in aridland regions. One of the key elements of its success in colonizing nutrient-poor sites is its capability for symbiotic nitrogen fixation. This trait, combined with a remarkable plasticity in environmental tolerance that allows it to establish, grow, and displace native species in disturbed areas and old fields. It thrives on poor soil, has low seedling mortality rates, and reaches reproductive maturity in a few years (Shafroth et al. 1995). Although Russian olive provides a plentiful source of edible fruits for birds, which aid in the long distance dispersal of its seeds, bird species richness is actually higher in riparian areas dominated by native vegetation (Brock 1998).
Acacia nilotica (Fabaceae, prickly acacia), a species native across semi-arid Africa through the Middle East to India, provides an interesting case study. Following its introduction more than a century ago, it has had extensive invasion into subtropical grasslands of Queensland and Indonesia with a major impact on the vegetation structure of these areas (Brown and Carter 1998). Extraordinarily, although Australia has more than 1,000 species of Acacia, no native taxon has filled the niche taken over by A. nilotica.
Tree invasions into tropical alpine ecosystems
The eucalyptus forests that currently dominate many high Andean puna and altiplano areas of Peru and Bolivia represent a massive transformation of a high mountain landscape that had for many centuries previously been largely devoid of trees (Dickinson 1969; Gade 1975; Farley 2007; Luzar 2007). Eucalyptus globulus (Myrtaceae) was widely planted in the hope that it would both provide an alternate fuel source and reduce pressure upon existing forests at lower elevations. However, eucalyptus has been associated with environmental degradation as well. Eucalyptus depletes water supplies and enhances soil acidity, although the degree of nutrient depletion is still debated (see Mitchell 1991; Doughty 2000 for opposing perspectives).
Pines have more recently been extensively introduced in high Andean páramo habitats, and have escaped from these plantations. Mexican pine species have been favored in these plantations, with Pinus patula the most commonly planted species. The impacts of these plantations on soil nutrient pools and water supply have proven to be complex and site specific, but there are clear negative effects on the diversity and composition of the páramo grasslands (Hofstede et al. 2002; van Wesenbeeck et al. 2003).
Global change and tree invasions
While traditional explanations of the range expansion tree and shrub species into grassland areas have focused on changes in climate, grazing history, and fire regime as causal factors, it has been hypothesized that anthropogenic increases in atmospheric CO-2 may favor this process (Idso 1992; Polley et al. 1994; Bond and Midgley 2000). This argument is based on the fact that photosynthesis in woody plants with C3 metabolism in the Southwestern United States and tropical savannas are favored over that in the C4 grasses they are replacing under conditions of higher ambient CO2. While this hypothesis has been subject to criticism (Archer et al. 1995), recent modeling studies suggest that increasing atmospheric concentrations of CO2 will force transitions to vegetation states characterized by higher biomass and/or woody-plant dominance (Higgins and Scheiter 2012).
Conclusions
There is strong evidence that the suite of life-history traits possessed by certain woody plants associated with dispersal and seedling survival enables them to rapidly transform ecosystems that previously had a low or no representation of trees. However it is not always clear whether these treeless areas are naturally treeless, or instead represent reversions from induced stable ecosystems resulting from historical legacies of burning or grazing.
Many of the major case studies of tree invasions into treeless areas come from examples in the Southern Hemisphere. It may well be that phylogenetic constraints and the absence of native tree species with specialized suites of adaptive life history and ecophysiological traits may explain some of this pattern, particularly as seen in the spectacular success of many pines in the Southern Hemisphere. More difficult to explain are the large number of North American examples compared to Europe and Asia for tree invasions into grassy wetlands and arid and semi-arid riparian habitats. This may be in part explained by a greater focus of study in North America.
Several groups of trees and woody shrubs have emerged as “winners” in a wide range of novel environments normally dominated by herbaceous or low shrubland vegetation. The major players in these invasions are pines and woody legumes capable of fixing nitrogen such as species of Acacia and Prosopis that can establish, grow, proliferate, spread and persist in a very wide range of conditions. In many cases it is clear that key life-history traits are the fundamental element in such successful invasions. For example serotinous follicles in Hakea, and nitrogen fixation in E. angustifolia and M. faya appear to provide some of the same success seen in pines and woody legumes respectively.
Successful tree invasion often brings significant changes in ecosystem processes and services such as carbon sequestration, water use, biogeochemical cycling, and wildlife habitat. There are also potential changes in belowground processes such as increased ectomycorrhizal inoculums, shifts in ratios of soil bacteria to fungi, loss of soil carbon, and shifts in soil chemistry that may favor subsequent invasion. Such changes provide important lessons for ecosystem ecology and priorities for future research.
Finally, recent hypotheses that elevated CO2 is driving a general increase in woody plant cover across large parts of the world need to be rigorously tested. If this enhancement of tree invasion is correct it has huge implications for understanding and managing future invasions of non-native trees and shrubs into treeless areas.
References
Aikio S, Duncan RP, Hulme PE (2010) Lag-phases in alien plant invasions: separating the facts from the artefacts. Oikos 119:370–378
Allen JA (1998) Mangroves as alien species: the case of Hawaii. Global Ecol Biogeogr Lett 7:61–67
Archer S (1989) Have southern Texas savannas been converted to woodlands in recent history. Am Nat 134:545–561
Archer S (1994) Woody plant encroachment into Southwestern grasslands and savannas: rates, patterns and proximate causes. In: Vavra M, Laycock W, Piepe R (eds) Ecological implications of livestock herbivory in the West. Soc Range Manage, Denver, pp 13–68
Archer S, Scifres C, Bassham C, Maggio R (1988) Autogenic succession in a subtropical savanna: rates, dynamics and processes in the conversion of grassland to thorn woodland. Ecol Monogr 58:111–127
Archer S, Schimel DS, Holland EA (1995) Mechanisms of shrubland expansion: land use, climate or CO2? Climatic Change 29:91–99
Ashkannejhad S, Horton TR (2006) Ectomycorrhizal ecology under primary succession on coastal sand dunes: interactions involving Pinus contorta, suilloid fungi and deer. New Phytol 169:345–354
Bahre CJ (1991) A legacy of change: historic human impact on vegetation of the Arizona borderlands. University of Arizona Press, Tucson 231 pp
Bodel MJ, Ferriter A, Thayer DD (1994) The biology, distribution, and ecological consequences of Melaleuca quinquenervia in the Everglades. In: Davis SM, Ogden JC (eds) Everglades, the ecosystem and its restoration. St. Lucie Press, Delray Beach, pp 341–355
Bond WJ, Midgley GF (2000) A proposed CO2-controlled mechanism of woody plant invasion in grasslands and savannas. Global Change Biol 6:865–870
Brock JH (1998) Invasion, ecology and management of Elaeagnus angustifolia (Russian olive) in the southwestern United States of America. In: Starfinger U, Edwards K, Kowarik I et al (eds) Plant invasions: ecological mechanisms and human responses. Backhuys, Leiden, pp 123–136
Brown JR, Carter J (1998) Spatial and temporal patterns of exotic shrub invasion in Australian tropical grassland. Landsc Ecol 13:93–102
Browning DM, Archer SR, Asner GP, McClaran MP, Wessman CA (2008) Woody plants in grasslands: post-encroachment stand dynamics. Ecol Appl 18:928–944
Bruce KA, Cameron GN, Harcombe PA (1995) Initiation of a new woodland type on the Texas coastal prairie by the Chinese tallow tree (Sapium sebiferum (L) Roxb). Bull Torrey Bot Club 122:215–225
Bruns TD, Peay KG, Boynton PJ, Grubisha LC, Hynson NA, Nguyen NH, Rosenstock NP (2009) Inoculum potential of Rhizopogon spores increases with time over the first 4 yr of a 99-yr spore burial experiment. New Phytol 181:463–470
Carilla J, Grau HR (2010) 150 Years of tree establishment, land use and climate change in montane grasslands, northwest Argentina. Biotropica 42:49
Carlquist S (1974) Island biology. Columbia University Press, New York
Collier FA, Bidartondo MI (2009) Waiting for fungi: the ectomycorrhizal invasion of lowland heathlands. J Ecol 97:950–963
Coop JD, Givnish TJ (2008) Constraints on tree seedling establishment in montane grasslands of the Valles Caldera, New Mexico. Ecology 89:1101–1111
Di Tomaso JM (1998) Impact, biology, and ecology of saltcedar (Tamarix spp.) in the Southwestern United States. Weed Techn 12:326–336
Dickie IA, Reich PB (2005) Ectomycorrhizal fungal communities at forest edges. J Ecol 93:244–255
Dickie IA, Bolstridge N, Cooper JA, Peltzer DA (2010) Co-invasion by Pinus and its mycorrhizal fungi. New Phytol 187:475–484
Dickie IA, Yeates GW, St J, Mark G, Stevenson BA, Scott JT, Rillig MC, Peltzer DA, Orwin KH, Kirschbaum MUF, Hunt JE, Burrows LE, Barbour MM, Aislabie J (2011) Ecosystem service and biodiversity trade-offs in two woody successions. J Appl Ecol 48:926–934
Dickie IA, Davis M, Carswell FE (2012) Quantification of mycorrhizal limitation in beech spread. NZ J Ecol 36:210–215
Dickinson JC (1969) The eucalypt in the Sierra of southern Peru. Ann Assoc Am Geogr 59:294–307
Diez J (2005) Invasion biology of Australian ectomycorrhizal fungi introduced with eucalypt plantations into the Iberian Peninsula. Biol Invasions 7:3–15
Doughty R (2000) The eucalyptus: a natural and commercial history of the gum tree. Johns Hopkins University Press, Baltimore
Duarte LS, Machado RE, Hartz SM, Pillar VD (2006) What saplings can tell us about forest expansion over natural grasslands. J Veg Sci 17:799–808
Ehrenfeld JG (2003) Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 6:503–523
Elton CS (1958) The ecology of invasions by animals and plants. Metheun, London
Farley KA (2007) Grasslands to tree plantations: forest transition in the Andes of Ecuador. Ann Assoc Am Geogr 97:755–771
Gade D (1975) Plants, man and the land in the Vilcanota Valley of Peru. Junk Publishers, The Hague
Geesing D, Felker P, Bingham RL (2000) Influences of mesquite (Prosopis glandulosa) on soil nitrogen and carbon development: implications for global carbon sequestration. J Arid Environ 46:157–180
Glenn EP, Nagler PL (2005) Comparative ecophysiology of Tamarix ramosissima and native trees in western U.S. riparian zones. J Arid Environ 61:419–446
Grotkopp E, Rejmánek M, Rost TL (2002) Toward a causal explanation of plant invasiveness: seedling growth and life-history strategies of 29 pine (Pinus) species. Am Nat 159:396–419
Guo LB, Gifford R (2002) Soil carbon stocks and land use change: a meta analysis. Global Change Biol 8:345–360
Hailu S, Demel T, Sileshi N, Fassil A (2004) Some biological characteristics that foster the invasion of Prosopis juliflora (Sw.) DC. at middle awash rift valley area, north- eastern Ethiopia. J Arid Environ 58:135–154
Halpern CB, Antos JA, Rice JM et al (2010) Tree invasion of a montane meadow complex: temporal trends, spatial patterns, and biotic interactions. J Veg Sci 21:717–732
Hastings JR, Turner RM (1966) The changing mile. University of Arizona Press, Tucson
Haugo RD, Halpern CB (2010) Tree age and tree species shape positive and negative interactions in a montane meadow. Botany 88:488–499
Hibbard KA, Schimel DS, Archer S, Ojima DS, Parton W (2003) Grassland to woodland transitions: integrating changes in landscape structure and biogeochemistry. Ecol Appl 13:911–926
Higgins SI, Scheiter S (2012) Atmospheric CO2 forces abrupt vegetation shifts locally, but not globally. Nature. doi:10.1038/nature11238
Hofstede RGM, Groenendijk JP, Coppus R, Fehse JC, Sevink J (2002) Impact of pine plantations on soils and vegetation in the Ecuadorian high Andes. Mt Res Develop 22:159–167
Hopper SD (2009) OCBIL theory: towards an integrated understanding of the evolution, ecology and conservation of biodiversity on old, climatically buffered, infertile landscapes. Plant Soil 322:49–86
Huxman TE, Wilcox BP, Breshears D, Scott RL, Snyder KA, Small EE, Hultine K, Pockman WT, Jackson RB (2005) Ecohydrological implications of woody plant encroachment. Ecology 86:308–319
Idso SB (1992) Shrubland expansion in the American Southwest. Clim Change 22:85–86
Jackson RB, Banner JL, Jobbágy EG, Pockman WT, Wall DH (2002) Ecosystem carbon loss with woody plant invasion of grasslands. Nature 418:623–626
Jäger H, Tye A, Kowarik I (2007) Tree invasion in naturally treeless environments: impacts of quinine (Cinchona pubescens) trees on native vegetation in Galapagos. Biol Conserv 140:297–307
Kaur R, Gonzáles WL, Llambi LD, Soriano PJ, Callaway RM et al (2012) Community impacts of Prosopis juliflora invasion: biogeographic and congeneric comparisons. PLoS ONE 7(9):e44966. doi:10.1371/journal.pone.0044966
Kleinbauer I, Dullinger S, Peterseil J, Essl F (2010) Climate change might drive the invasive tree Robinia pseudacacia into nature reserves and endangered habitats. Biol Conserv 143:382–390
Langdon B, Pauchard A, Aguayo M (2010) Pinus contorta invasion in the Chilean Patagonia: local patterns in a global context. Biol Invasions 12:3961–3971
Le Maitre DC, Versfeld DB, Chapman RA (2000) The impact of invading alien plants on surface water resources in South Africa: a preliminary assessment. Water SA 26:397–407
Le Maitre DC, Gaertner M, Marchante E, Ens EJ, Holmes PM, Pauchard A, O’Farrell PJ, Rogers AM, Blanchard R, Blignaut J, Richardson DM (2011) Impacts of invasive Australian acacias: implications for management and restoration. Divers Distrib 17:1015–1029
League K, Veblen T (2006) Climatic variability and episodic Pinus ponderosa establishment along the forest-grassland ecotones of Colorado. For Ecol Manag 228:98–107
Ledgard N (2001) The spread of lodgepole pine (Pinus contorta, Dougl.) in New Zealand. For Ecol Manag 141:43–57
Luzar J (2007) The political ecology of a “forest transition”: eucalyptus forestry in the southern Peruvian Andes. Ethnobot Res Appl 5:85–93
MacDougall AS, Gilbert B, Levine JM (2009) Plant invasions and the niche. J Ecol 97:609–615
Mack RN (2003) Phylogenetic constraint, absent life forms, and preadapted alien plants: a prescription for biological invasions. Intern J Plant Sci 164:S185–S196
Martin MR, Tipping PW, Sickman JO (2009) Invasion by an exotic tree alters above and belowground ecosystem components. Biol Invasions 11:1883–1894
Marx DH (1991) The practical significance of ectomycorrhizae in forest establishment. Ecophysiology of ectomycorrhizae of forest trees. Marcus Wallenberg Foundation, Falun, Sweden, pp 54–90
Mazia CN, Chaneton EJ, Ghersa CM, León JC (2001) Limits to tree species invasion in Pampean grassland and forest plant communities. Oecologia 128:594–602
McClaran MP (2003) A century of vegetation change on the Santa Rita experimental range. Pages 16–33 in Santa Rita experimental range: one-hundred years (1903–2003) of accomplishments and contributions. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Tucson, Arizona
Merritt DM, Poff NL (2010) Shifting dominance of riparian Populus and Tamarix along gradients of flow alteration in western North American rivers. Ecol Appl 20:135–152
Merritt DM, Shafroth PB (2012) Edaphic, salinity, and stand structural trends in chronosequences of native and non-native dominated riparian forests along the Colorado River, USA. Biol Invasions 14:2665–2685
Miller EA, Halpern CB (1998) Effects of environment and grazing disturbance on tree establishment in meadows of the central Cascade Range, Oregon, USA. J Veg Sci 9:265–282
Mitchell B (1991) Peasants on the edge: crop, culture, and crisis in the Andes. University of Texas Press, Austin
Mooney HA (2008) The globalization of ecological thought. Ecology Institute, Oldendorf/Luhe, Germany
Nuñez MA, Horton TR, Simberloff D (2009) Lack of belowground mutualisms hinders Pinaceae invasions. Ecology 90:2352–2359
Pattison RR, Mack RN (2008) Potential distribution of the invasive tree Triadica sebifera (Euphorbiaceae) in the United States: evaluating CLIMEX predictions with field trials. Glob Change Biol 14:813–826
Peña E, Hidalgo M, Langdon B, Pauchard A (2008) Patterns of spread of Pinus contorta Dougl. ex Loud in a natural reserve in southern South America. For Ecol Manag 256:1049–1054
Pillar VD, Quadros FLF (1997) Grassland–forest boundaries in southern Brazil. Coenoses 12:119–126
Polley HW, Johnson HB, Mayeaux HS (1994) Increasing CO2: competitive responses of the C4 grass Schizachyrium and grassland Prosopis. Ecology 75:976–988
Pringle A, Bever JD, Gardes M, Parrent JL, Rillig MC, Klironomos JN (2009) Mycorrhizal symbioses and plant invasions. Ann Rev Ecol Evol Syst 40:699–715
Rice JM, Halpern CB, Antos JA et al (2012) Spatio-temporal patterns of tree establishment are indicative of biotic interactions during early invasion of a montane meadow. Plant Ecol 213:555–568
Richardson DM, Bond WJ (1991) Determinants of plant distribution: evidence from pine invasions. Am Nat 137:639–668
Richardson DM, Cowling RM (1992) Why is mountain fynbos invasible and which species invade? In: Van Wilgen BW, Richardson DM, Kruger FJ, van Hensbergen HJ (eds) Fire in South African mountain fynbos. Springer, Berlin, pp 161–181
Richardson DM, Gaertner M (2013) Plant invasions as builders and shapers of novel ecosystems. In: Hobbs RJ, Higgs EC, Hall CM (eds) Novel ecosystems: intervening in the new ecological world order. Wiley-Blackwell, Oxford, pp 102–114
Richardson DM, Higgins SI (1998) Pines as invaders in the southern hemisphere. In: Richardson DM (ed) Ecology and biogeography of Pinus. Cambridge University Press, Cambridge, pp 450–473
Richardson DM, Rejmánek M (2011) Trees and shrubs as invasive alien species—a global review. Divers Distrib 17:788–809
Richardson DM, van Wilgen BW (2004) Invasive alien plants in South Africa; how well do we understand the ecological impacts? S Afr J Sci 100:45–52
Richardson DM, Williams PA, Hobbs RJ (1994) Pine invasions in the Southern Hemisphere: determinants of spread and invadability. J Biogeogr 21:511–527
Richardson DM, Allsopp N, D’Antonio CM, Milton SJ, Rejmánek M (2000) Plant invasions: the role of mutualisms. Biol Rev 75:65–93
Richardson DM, van Wilgen BW, Nuñez MA (2008) Alien conifer invasions in South America: short fuse burning? Biol Invasions 10:573–577
Rudel TK, Coomes OT, Moran E, Achard F, Angelsen A, Xu J, Lambin E (2005) Forest transition: towards a global understanding of land use change. Global Environ Change 15:23–31
Rundel PW, Nilsen ET, Sharifi M, Virginia R, Jarrell W, Kohl D, Shearer G (1982) Seasonal dynamics of nitrogen cycling for a Prosopis woodland in the Sonoran Desert. Plant Soil 67:343–353
Salgado Salomón ME, Barroetaveña C, Rajchenberg M (2011) Do pine plantations provide mycorrhizal inocula for seedlings establishment in grasslands from Patagonia, Argentina? New For 41:191–205
Scholes RJ, Archer SR (1997) Tree–grass interactions in savannas. Ann Rev Ecol Syst 28:517544
Serbesoff-King K (2003) Melaleuca in Florida: a literature review on the taxonomy, distribution, biology, ecology, economic importance and control measures. J Aquatic Plant Manag 41:98–112
Shafroth PB, Auble GT, Scott ML (1995) Germination and establishment of native plains cottonwood (Populus deltoides Marshall subsp. monilifera) and the exotic Russian-olive (Elaeagnus angustifolia L.). Conserv Biol 9:1169–1175
Shafroth PB, Cleverly JR, Dudley L, Stuart J, Taylor JP, van Riper C, Weeks EP (2005) Control of Tamarix in the western U.S.—implications for water salvage, wildlife use, and riparian restoration. Environ Manag 35:231–246
Simberloff D (2011) Charles Elton. Neither founder nor siren, but prophet. In: Richardson DM (ed) Fifty years of invasion ecology. The legacy of Charles Elton, Wiley-Blackwell, Oxford, pp 11–24
Simberloff D, Nuñez MA, Ledgard NJ et al (2010) Spread and impact of introduced conifers in South America: lessons from other southern hemisphere regions. Austral Ecol 35:489–504
Stromberg JC, Chew MK, Nagler PL, Glenn EP (2009) Changing perceptions of change: the role of scientists in Tamarix and river management. Restor Ecol 17:177–186
Terwilliger J, Pastor J (1999) Small mammals, ectomycorrhizae, and conifer succession in beaver meadows. Oikos 85:83–94
Vale TR (1981) Tree invasion of montane meadows in Oregon. Am Midl Nat 105:61–69
Van Auken OW (2000) Shrub invasions of North American semiarid grasslands. Ann Rev Ecol Syst 31:197–215
van Wesenbeeck B, van Mourik T, Duivenvoorden JF, Cleef AM (2003) Strong effects of a plantation with Pinus patula on Andean subpáramo vegetation: a case study from Colombia. Biol Conserv 114:207–218
Vitousek PM, Walker LR (1989) Biological Invasion by Myrica faya in Hawaii—plant demography, nitrogen-fixation, ecosystem effects. Ecol Monogr 59:247–265
Wardle P (1985) New Zealand timberlines. 3. A synthesis. NZ J Bot 23:263–271
Wiemken V, Boller T (2006) Delayed succession from alpine grassland to savannah with upright pine: limitation by ectomycorrhiza formation? For Ecol Manag 237:492–502
Zalba SM, Villamil CB (2002) Woody plant invasion in relictual grasslands. Biol Invas 4:55–72
Acknowledgments
PWR is grateful for support from the University of California Los Angeles and the Stunt Ranch Santa Monica Mountains Reserve. IAD acknowledges Core funding for Crown Research Institutes from the New Zealand Ministry of Business, Innovation and Employment’s Science and Innovation Group. DMR acknowledges support from the DST-NRF Centre of Excellence for Invasion Biology and the National Research Foundation (grant 85417). The Oppenheimer Memorial Trust funded DMR’s participation in the workshop at Bariloche, Argentina, in September 2012 at which an early version of this paper was tabled. We thank all delegates at the workshop for their contributions to the ideas that are presented here.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rundel, P.W., Dickie, I.A. & Richardson, D.M. Tree invasions into treeless areas: mechanisms and ecosystem processes. Biol Invasions 16, 663–675 (2014). https://doi.org/10.1007/s10530-013-0614-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-013-0614-9