
Abstract
The emerald ash borer (EAB) Agrilus planipennis, first detected in 2002 in the vicinity of Detroit, Michigan, USA, is one of the most recent in a long list of introduced insect pests that have caused serious damage to North American forest trees, in this case ash trees in the genus Fraxinus. We used data from Project FeederWatch, a citizen science program focused on winter bird populations, to quantify the effects of EAB invasion on four species of resident, insectivorous birds known or likely to be EAB predators: three woodpecker species and the white-breasted nuthatch (Sitta canadensis). We compared relative numbers of birds within 50 km of the epicenter of the region where EAB was first detected, an area known to have suffered high ash tree mortality by 2008, to numbers 50–100 km from the epicenter and to control sites within 50 km of five comparable Midwestern cities where damage due to EAB has yet to be severe. We found evidence for significant effects on all four of the species in response to the EAB invasion in the highly impacted region, with red-bellied woodpeckers (Melanerpes carolinus) and white-breasted nuthatches showing numerical increases while downy woodpeckers (Picoides pubescens) and hairy woodpeckers (Picoides villosus) initially declined but exhibited at least temporary increases several years later. Temporal correlation analyses failed to provide support for immigration being a major cause of the elevated numbers in the highly impacted area, and thus these results are consistent with the hypothesis that increases were due to enhanced survival and/or reproduction associated with the EAB invasion within the highly impacted area. Results suggest that the continuing invasion of EAB into new areas is likely to significantly alter avian communities, although not always in ways that will be easy to predict.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Biological invasions can have profound ecological effects, including alteration of food-web structure (Baxter et al. 2004), modification of nutrient cycles (Ehrenfeld 2003), and changes to ecosystem processes such as fire regimes (D’Antonio and Vitousek 1992). The most commonly reported trophic impacts, however, are adverse effects on prey availability either directly or via interspecific competition. For example, invasions by both generalist and specialist predators often result in greatly diminished numbers of prey species (Beisner et al. 2003; Blackburn et al. 2004). Similarly, considerable attention has been given to the adverse effects of herbivore invasions on host plant species (Liebhold and Elkinton 1991; Husheer et al. 2003), and for some species of introduced herbivores, negative population-level effects have been demonstrated on native species competing with invasives for the same resource (Strayer and Malcom 2007).
Despite the focus given to invasions by herbivore consumers, relatively little is known about the ecological consequences of such invasions on higher trophic levels. Invasions may benefit some predators if invasive prey represent a new and abundant food supply. Alternatively, because the increased food supply provided by an invasive species may negatively affect native prey species via apparent competition, invasives have the potential to adversely influence species at higher trophic levels (Bonsall and Hassell 1997; Settle and Wilson 1990; Meiners 2007).
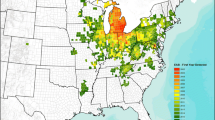
The recent invasion of the emerald ash borer (EAB) Agrilus planipennis, an herbivore species that has exhibited explosive growth in North America, provides an opportunity to document the effects of an invasive consumer on populations of vertebrate predators. EAB is a phloem-feeding beetle native to northern China, Korea, Japan, and eastern Russia, that was determined to be the cause of widespread mortality of ash trees (Fraxinus sp.) in the Detroit, Michigan and nearby Windsor, Ontario areas in 2002, although it is believed that the species probably arrived 10–20 years previously (Poland and McCullough 2006). In its native range, populations of EAB are generally sparse and damage to hosts is uncommon. In North America, however, native ash species do not possess effective defenses against EAB and feeding damage is extensive, causing crown dieback and eventually tree death. Since its discovery in 2002, populations of EAB have increased and spread to nine additional states and to Quebec (Poland and McCullough 2006; Kovacs et al. 2010) (Fig. 1a).

Location of sample data relative to the EAB invasion. a Extent of EAB in the United States and Canada as of April 2011. Concentric black circles designate regions <50 km and 50–100 km from the EAB epicenter. b Location of Project FeederWatch sample locations used in the analyses
In areas where the species has been established for several years, populations typically become very abundant and kill most ash trees present. Pugh et al. (2011) analyzed regional forest inventory data from Michigan and found that between 2004 and 2009, the density of ash live wood volume decreased from 12.7 to 3.2 m3 ha−1 in forest stands within 50 km of the EAB invasion epicenter, a decrease of 75 %. Based on an aerial survey conducted in 2007, Smith et al. (2009) estimated that EAB had killed 54 million trees in Michigan, Ohio, and Indiana. While many of the trees attacked by EAB are in naturally regenerating forests, many are also in urban settings where ash has been widely planted as a street tree (MacFarlane and Meyer 2005; Kovacs et al. 2010).
As many as 60,000 or more EAB individuals may develop from a single tree (McCullough and Siegert 2007). Thus, the massive numbers of EAB-infested trees represents a large resource for predators. Indeed, one of the most common sources of mortality for EAB larvae and pupae are avian generalist predators, including several species of woodpeckers (Picidae), that have been found to depredate up to 95 % (mean = 44 %) of emerging EAB from sites in southeastern Michigan (Cappaert et al. 2005b; Lindell et al. 2008; Duan et al. 2010). This finding led these authors to conclude that although the EAB invasion is a catastrophe for ash trees, it may be providing a ‘feast for woodpeckers’ (Cappaert et al. 2005a). Thus, in addition to the massive impact of EAB on lower trophic levels (i.e., millions of dead trees), it may be affecting species of native birds in a variety of ways either negatively (by adverse effects on habitat or native prey populations) or positively (by providing a ‘trophic subsidy’ positively influencing their reproduction and survivorship; Branch and Steffani 2004; Rodriguez 2006). In addition, the large pulse of resources provided by EAB could potentially attract immigrants from surrounding regions. Thus, impacts of EAB on birds may extend beyond the areas directly infested, and the resulting population increases of subsidized bird species in the affected areas could have cascading effects on the other species competing for cavities and food supplies. Testing for a trophic subsidy is a first step toward understanding the potential widespread impacts of the EAB on avian populations.
We investigated the effects of the recent EAB invasion on abundance of four species of resident, insectivorous birds. Species considered were the downy woodpecker (Picoides pubescens), hairy woodpecker (Picoides villosus), and red-bellied woodpecker (Melanerpes carolinus), three species that are known to forage on ash trees infested by EAB (Lindell et al. 2008), and the white-breasted nuthatch (Sitta canadensis), a common bark-gleaning species that is also a potential predator of EAB. All are species that might be expected to reap a substantial nutritional benefit and experience relatively high reproductive success in areas where EAB are common. Several species of woodpeckers have been shown to respond to local increases in prey abundance (e.g., Murphy and Lehnhausen 1998), making larger-scale responses likely. Furthermore, all four species are cavity-nesters, and thus might be expected to benefit from an increase in nest substrate as trees are killed by EAB (Cockle et al. 2011). This would be particularly true for downy woodpeckers and red-bellied woodpeckers, which regularly nest in dead limbs or snags (Shackelford et al. 2000; Jackson and Ouellet 2002), and for white-breasted nuthatches, which often use old woodpecker cavities as nest sites (Grubb and Pravosudov 2008).
Our primary goal was to quantify the numerical changes in populations of these avian predators since the EAB invasion in 2002, the prediction being that all four species were likely to exhibit increases either due the increased food supply offered by EAB or the increased availability of nesting substrate as a consequence of elevated tree mortality in highly-impacted sites. Secondarily, we attempted to detect whether birds were being attracted to areas where tree mortality due to EAB was particularly severe or whether increases were likely to be due to increased reproduction and survival within the impacted areas. We approached this problem using data collected as part of a citizen science program (FeederWatch.org) uniquely suited to measuring year-to-year population changes over the geographic and temporal scale of interest.
Methods
Siegert et al. (2009) estimated the location of the presumed initial EAB introduction using dendrochronological methods to reconstruct EAB’s historical spread. Based on their estimates of tree mortality across a five-county region, they concluded that EAB was first established in the early to mid-1990s in the vicinity of Westland-Garden City, Michigan, approximately 25 km west of downtown Detroit. By 2008, EAB had killed >50 % of ash trees within 50 km of this epicenter (Pugh et al. 2011). Thus, we considered sites within 50 km of the epicenter, basically the city of Detroit and its suburbs, to have been most highly impacted by EAB, and refer to them as ‘highly-impacted’ (or ‘HIMP’) sites (Fig. 1b). For comparison, we determined numerical abundance of the bird species in two sets of sites where the impact of EAB has not yet been severe. First, we used sites 50–100 km from the EAB epicenter where tree mortality due to the insects, although detectable, was considerably less than nearer the epicenter (Pugh et al. 2011). We refer to these as ‘lesser impacted’ (or ‘LIMP’) sites. Second, we used sites <50 km from five large Midwestern cities of more or less comparable size: Cincinnati, Cleveland, and Columbus (all in Ohio), Indianapolis (Indiana), and Pittsburgh (Pennsylvania). All five cities were sites where EAB had not yet caused major damage, although EAB had in some cases been reported (Fig. 1a). We refer to these five cities as ‘control’ sites. A summary of the major characteristics of sites used in the analyses is provided in Table 1. Although there are differences, the three sets of sites (using the average of the five control cities) contained reasonably comparable proportions of developed (25–42 %) and forested/wetland (12–27 %) habitat.
A primary difference between the LIMP and control sites was that the former, being adjacent to the highly impacted area, was potentially a source for immigrants attracted to the highly-impacted region or, alternatively, a sink for birds emigrating from the highly-impacted region. Otherwise, because of habitat differences (Table 1), we did not make any a priori predictions about how the LIMP and control sites might differ.
All four bird species considered commonly visit backyard bird feeders. This made it possible for us to use data from Project FeederWatch (PFW), a program in which volunteer observers record the maximum observed number of individuals of each species at backyard feeders over a 2-day period at weekly intervals during the winter (Bonter and Hochachka 2009). FeederWatch data have been found in prior studies to both match patterns derived from the more commonly used Audubon Christmas Bird Counts (Cooper et al. 2007) and reflect real variation in resident bird populations (Wells et al. 1998; Lepage and Francis 2002). Prior studies have indicated that the availability of wild foods can reduce the reliance of birds on feeders (Chamberlain et al. 2007); to the extent that this might be occurring in our study due to the availability of overwintering EAB larvae (Wang et al. 2010), it might potentially reduce numbers of birds at feeders in the highly impacted area and render results conservative.
Observations from the public are reviewed for accuracy using automated filters (Bonter and Cooper 2012). All observations reporting more individuals of a species than were reported by 99 % of all checklists were eliminated, since prior work has indicated that a large proportion of such extraordinarily large counts are erroneous (States et al. 2009). This limited the data we considered to sites with a maximum of five downy woodpeckers, three hairy woodpeckers, two red-bellied woodpeckers, and four white-breasted nuthatches. Each winter spanning two calendar years is referred to as a ‘season’ (i.e., winter 2001–2002 is referred to as the 2001 season). Analyses were conducted through the 2011 season (winter 2011–2012).
We analyzed data for each of the four bird species using Generalized Linear Mixed Models (GLMMs) with Poisson errors. The maximum number of birds observed per site was the dependent factor, ‘site’ was a random effect, and the fixed effects were (1) the number of half-days of effort expended (1–4 as a categorical variable); (2) date (both linear and quadratic terms as days since 1 Oct.); (3) season (as a categorical variable with 2001–2003 averaged together and used as an estimate of pre-EAB invasion densities, along with the 2004–2011 seasons treated as separate units); (4) dataset (HIMP sites, LIMP sites, or control sites); and (5) the ‘season × dataset’ interaction. Statistical significance of these interactions (one for each season post-EAB invasion) indicated differences in numbers of birds in the highly impacted area compared to the non-highly impacted sites. The number of different sites used in these analyses was 167 of which 29 were in the HIMP area, 21 were in the LIMP area, and 117 were in the five control cities. The number of total observations (i.e., site × date combinations) ranged from 22,737 to 22,963 per season. On average, individual participants reported data in 3.1 ± 2.7 (mean ± standard deviation) of the nine seasons and maintained an average of 6.2 ± 3.1 feeders per site.
In order to gain insight into the mechanism behind the changes observed in the analyses, we performed temporal correlations across years between the mean number of birds in the HIMP sites compared to those in the adjacent LIMP sites. Correlations were done for each species and for each of three temporal combinations, which involved correlating the mean number of birds in the HIMP sites in year t n versus (1) LIMP sites in year t n−1 (lag−1), (2) LIMP sites in year t n (lag−0), and (3) LIMP sites in year t n+1 (lag + 1). Our predictions were as follows: if high numbers of young birds produced in surrounding areas were attracted to and tended to immigrate into the highly-impacted areas the following year, there should be strong positive correlations between LIMP sites in year t n−1 and HIMP sites the following year (year t n ). If second-year and older birds were attracted to HIMP sites from nearby surrounding areas due to the relatively high availability of food, there should be negative correlations between LIMP sites in year t n and HIMP sites the same year. Finally, if high productivity due to resource availability in highly-impacted areas resulted in subsequent dispersal of birds to surrounding areas, there should be strong positive correlations between HIMP sites in year t n and LIMP sites the following year (year t n+1). As controls for these tests, we performed the same cross-correlations between HIMP and the control sites, which were sufficiently far away (>200 km) that we would not predict the above relationships to hold given that these species are unlikely to commonly disperse such large distances.
All analyses were performed in R 2.10.1 (R Development Core Team 2009); the GLMMs were performed using the ‘lme4’ library (procedure ‘lmer’).
Results
There were significant changes in abundance of all four species in the HIMP sites relative to LIMP and/or control sites based on the GLMMs (Fig. 2). Increases were particularly evident for red-bellied woodpeckers starting in the 2006 season and white-breasted nuthatches starting in the 2005 season and continued through the end of the study in winter 2011–2012. In contrast, both downy woodpeckers and hairy woodpeckers exhibited significant declines in the highly impacted area one to three years following the initial discovery of EAB. Numbers then generally recovered (with the exception of hairy woodpeckers in 2010), with significantly elevated numbers relative to the control sites during one of the latter years of the study (2009 for downy woodpeckers; 2011 for hairy woodpeckers).

Estimates of the relative numbers of the four species of birds in sites highly impacted by EAB (sites <50 km from EAB epicenter; N = 29 sites) compared to the sites in lesser impacted areas (sites 50–100 km from the epicenter; N = 21) and sites <50 km from five Midwestern control cities (N = 117) based on the GLMMs, 2004–2011 seasons. Initial population sizes are indicated by the straight dotted line. Values and significance (as indicated by the different colors) are based on the ‘year × site’ interaction terms
We tested for evidence of immigration into or emigration out of the highly-impacted region by means of temporal correlations (Table 2). lag−1 correlations between LIMP sites and HIMP sites were generally low or negative for all four species (mean r = −0.05), failing to support the hypothesis that young birds in surrounding areas were attracted to and immigrated into HIMP sites the following year. In contrast, lag−0 correlations between LIMP sites and HIMP sites were generally positive and high (mean r = 0.46) and were in all cases greater than the lag−1 correlations, failing to support the pattern predicted if older birds from surrounding areas were attracted to and immigrated into HIMP sites from LIMP sites. Finally, the evidence suggesting that birds from HIMP sites were dispersing out into surrounding LIMP sites was equivocal at best. The lag+1 correlations between HIMP sites and LIMP sites were fairly high (mean r = 0.39), but this was primarily due to high correlations for downy and hairy woodpeckers, the two species that did not exhibit clear increases in response to the EAB invasion (Fig. 2). More compellingly, the mean lag+1 correlations between the HIMP and LIMP sites were generally much greater than the corresponding lag+1 correlations between HIMP sites and the considerably more distant control sites (mean r = 0.04). However, this was not true of the white-breasted nuthatch, one of the two species exhibiting a clear increase due to EAB in the GLMMs.
Discussion
These results support the hypothesis that the invasion and subsequent spread of EAB has had significant impacts on populations of the four insectivorous, resident bird species considered in this study. Relative abundance of red-bellied woodpeckers and white-breasted nuthatches clearly increased in the highly impacted area following the detection of EAB around 2002 and have not returned to baseline levels as of the 2011 season. Populations of downy and hairy woodpeckers initially declined significantly and have subsequently switched back and forth but were at significantly elevated levels relative to control sites during the 2009 (downy woodpecker) and 2011 (hairy woodpecker) season. Thus, the response of these four species to the EAB invasion was surprising to the extent that only two of the species exhibited clear increases while the other two apparently suffered decreases, at least initially. Such variability is perhaps to be expected given the many routes, both direct and indirect, that an invasive such as EAB can influence predator populations. Similarly variable and unexpected population responses were detected in a recent analysis of the effects of gypsy moth (Lymantria dispar) outbreaks on populations of seven species of North American woodpeckers (Koenig et al. 2011).
Secondarily, we attempted to detect evidence for immigration into and out of the highly-impacted sites using temporal cross-correlation analysis. Results failed to support the hypothesis that increases in HIMP sites were due to birds from surrounding regions, either young produced the prior year or adults the same year, immigrating into the area. We found some evidence to support the hypothesis that birds in HIMP sites were emigrating out into nearby, lesser-impacted surrounding areas, but the evidence was equivocal in being most influenced by the two species for which we observed no clear increases in response to the EAB invasion. We conclude that the increase in numbers of birds in the HIMP sites is primarily due to increased reproductive success and survivorship resulting from the increased abundance of resources—either food and/or nesting substrate—resulting from the EAB invasion. Demographic data, preferrably on marked individuals, would be necessary to confirm this conclusion.
It remains to be seen how EAB invasion will affect long-term trends in abundance of these bird species. It is possible that the observed elevation in abundance in red-bellied woodpeckers and white-breasted nuthatches may be short-lived, since EAB populations are likely to eventually decline, followed by declines in the birds, once most of the ash trees in the region have died. Such a result would be consistent with population cycles of the black-backed woodpecker (Picoides arcticus), which tracks short-term bursts of food following forest fires (Murphy and Lehnhausen 1998; Nappi et al. 2010). In contrast, the effects of availability of dead trees providing nesting sites may persist longer, particularly to the extent that this resource limits avian population growth.
Predation by birds has been reported as the largest source of EAB mortality in outbreak populations (Duan et al. 2010). As such, the numerical response documented here involving species known to depredate EAB may contribute to regulatory pressure on the EAB populations. Such pressure is apparently not strong enough to prevent the explosive growth of invading EAB populations but it does suggest that EAB spread might be faster were it not for the numerical increase of insectivorous birds resulting from the invasion. It is also possible that elevated predator densities associated with the EAB invasion may act negatively on native prey insects via apparent competition (Holt 1977; Bonsall and Hassell 1997), an effect that could then negatively influence other predators. Thus, it is perhaps not surprising that an invasive species such as EAB effects predators in different ways, some positive (such as was observed in red-bellied woodpeckers and white-breasted nuthatches) and some negative (as seen in downy and hairy woodpeckers, at least initially). Careful monitoring of abundance and demography of other prey species, as well as on avian predators themselves, in EAB invaded areas would clearly be of interest.
The concept of predators responding numerically to host densities is not new. As mentioned above, several species of birds, including woodpeckers and others, are known to respond positively to increases in food abundance, most dramatically after fires (Murphy and Lehnhausen 1998; Covert-Bratland et al. 2006; Nappi et al. 2010), but also in response to regional insect outbreaks of gypsy moths (Barber et al. 2008; Koenig et al. 2011) and periodical cicadas Magicicada sp. (Koenig and Liebhold 2005). The results presented here contribute new information, however, about how insectivorous bird populations can respond to large-scale eruptions of insect populations, and is unique in allowing us to falsify, at least circumstantially, the hypothesis that these responses are being driven by immigration of birds into regions of abundant food. Hence, our data indicate that numerical responses of at least some avian predators will be observed across the expanding area impacted by EAB. Similar effects have been suggested for red-headed woodpeckers (Melanerpes erythrocephalus), whose densities increased in the mid-twentieth century apparently in response to widespread mortality of American chestnuts (Castanea dentata) and American elms (Ulmus americana) resulting from introduced pathogenic fungi (Smith et al. 2000).
Our use of citizen science data to document regional ecological changes associated with a pest invasion also illustrates the potential that these data have as tools for investigating landscape-level ecological changes (Dickinson et al. 2010). Such changes can be detected in ‘real time’ using appropriate citizen science data, even when the geographic range of the expected effects is far greater than can otherwise be covered by one or even a coordinated group of researchers.
References
Barber NA, Marquis RJ, Tori WP (2008) Invasive prey impacts the abundance and distribution of native predators. Ecology 89:2678–2683
Baxter CV, Fausch KD, Murakami M, Chapman PL (2004) Fish invasion restructures stream and forest food webs by interrupting reciprocal prey subsidies. Ecology 85:2656–2663
Beisner BE, Ives AR, Carpenter SR (2003) The effects of an exotic fish invasion on the prey communities of two lakes. J Anim Ecol 72:331–342
Blackburn TM, Cassey P, Duncan RP, Evans KL, Gaston KJ (2004) Avian extinction and mammalian introductions on oceanic islands. Science 305:1955–1958
Bonsall MB, Hassell MP (1997) Apparent competition structures ecological assemblages. Nature 388:371–373
Bonter DN, Cooper CB (2012) Data validation in citizen science: a case study from Project FeederWatch. Front Ecol Environ 10:305–307
Bonter DN, Hochachka WM (2009) A citizen science approach to ornithological research: twenty years of watching backyard birds. In: Rich TD, Arizmendi C, Demarest E, Thompson C (eds) Tundra to tropics: connecting birds, habitats and people. McAllen, TX: Proceedings of 4th International Partners in Flight Conference, pp 453–458
Branch GM, Steffani CN (2004) Can we predict the effects of alien species? A case-history of the invasion of South Africa by Mytilus galloprovincialis (Lamarck). J Exp Mar Biol Ecol 300:189–215
Cappaert D, McCullough DG, Poland T (2005a) The upside of the emerald ash borer catastrophe: a feast for woodpeckers. In: Mastro V, Reardon R (eds) Emerald ash borer research and technology development meeting. USDA Forest Service Pub FHTET-2004-15, Romulus, pp 69–70
Cappaert D, McCullough DG, Poland TM, Siegert NW (2005b) Emerald ash borer in North America: a research and regulatory challenge. Am Entomol 51:152–165
Chamberlain DE, Gosler AG, Glue DE (2007) Effects of the winter beechmast crop on bird occurrence in British gardens: capsule woodland birds were significantly less likely to occur in gardens in years of high beechmast crop. Bird Study 54:120–126
Cockle KL, Martin K, Wesołowski T (2011) Woodpeckers, decay, and the future of cavity-nesting vertebrate communities worldwide. Front Ecol Environ 9:377–382
Cooper CB, Hochachka WM, Dhondt AA (2007) Contrasting natural experiments confirm competition between house finches and house sparrows. Ecology 88:864–870
Covert-Bratland KA, Block WM, Theimer TC (2006) Hairy woodpecker winter ecology in ponderosa pine forests representing different ages since wildfire. J Wildl Manag 70:1379–1392
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass–fire cycle, and global change. Annu Rev Ecol Syst 23:63–87
Dickinson JL, Zuckerberg B, Bonter DN (2010) Citizen science as an ecological research tool: challenges and benefits. Annu Rev Ecol Evol Syst 41:149–172
Duan JJ, Ulyshen MD, Bauer LS, Gould J, Van Driesche R (2010) Measuring the impact of biotic factors on populations of immature emerald ash borers (Coleoptera: Buprestidae). Environ Entomol 39:1513–1522
Ehrenfeld JG (2003) Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 6:503–523
Fry JA, Xian G, Jin S, Dewitz JA, Homer CG, Yang L, Barnes CA, Herold ND, Wickham JD (2011) Completion of the 2006 National Land Cover database for the conterminous United States. Photogramm Eng Remote Sensing 77:858–864
Grubb TC Jr, Pravosudov VV (2008) White-breasted nuthatch (Sitta carolinensis). In: Poole A (ed) The Birds of North America Online. Cornell Lab of Ornithology Ithaca. Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/054
Holt RD (1977) Predation, apparent competition, and the structure of prey communities. Theor Pop Biol 12:197–229
Husheer SW, Coomes DA, Robertson AW (2003) Long-term influences of introduced deer on the composition and structure of New Zealand Nothofagus forests. For Ecol Manag 181:99–117
Jackson JA, Ouellet HR (2002) Downy woodpecker (Picoides pubescens) In: Poole A (ed) The Birds of North America Online. Cornell Lab of Ornithology, Ithaca. Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/613
Koenig WD, Liebhold AM (2005) Effects of periodical cicada emergences on abundance and synchrony of avian populations. Ecology 86:1873–1882
Koenig WD, Walters EL, Liebhold AM (2011) Effects of gypsy moth outbreaks on North American woodpeckers. Condor 113:352–361
Kovacs KF, Haight RG, McCullough DG, Mercader RJ, Siegert NW, Leibhold AM (2010) Cost of potential emerald ash borer damage in U.S. communities, 2009–2019. Ecol Econ 69:569–578
Lepage D, Francis CM (2002) Do feeder counts reliably indicate bird population changes? 21 years of winter bird counts in Ontario, Canada. Condor 104:255–270
Liebhold AM, Elkinton JS (1991) Gypsy moth dynamics. Trends Ecol Evol 6:263–264
Lindell CA, McCullough DG, Cappaert D, Apostolou NM, Roth MB (2008) Factors influencing woodpecker predation on emerald ash borer. Am Midl Nat 159:434–444
MacFarlane DW, Meyer SP (2005) Characteristics and distribution of potential ash tree hosts for emerald ash borer. For Ecol Manag 213:15–24
McCullough DG, Siegert NW (2007) Estimating potential emerald ash borer (Coleoptera: Buprestidae) populations using ash inventory data. J Econ Entomol 100:1577–1586
Meiners SJ (2007) Apparent competition: an impact of exotic shrub invasion on tree regeneration. Biol Invasions 9:849–855
Murphy EC, Lehnhausen WA (1998) Density and foraging ecology of woodpeckers following a stand-replacement fire. J Wild Manag 62:1359–1372
Nappi A, Drapeau P, Saint-Germain M, Angers VA (2010) Effect of fire severity on long-term occupancy of burned boreal conifer forests by saproxylic insects and wood-foraging birds. Int J Wildland Fire 19:500–511
Poland TM, McCullough DG (2006) Emerald ash borer: invasion of the urban forest and the threat to North America’s ash resource. J For 104:118–124
Pugh SA, Liebhold AM, Morin RS (2011) Changes in ash tree demography associated with emerald ash borer invasion, indicated by regional forest inventory data from the Great Lakes states. Can J For Res 41:2165–2175
R Development Core Team (2009) R: a language and environment for statistical computing: R Foundation for Statistical Computing, Vienna, Austria. ISBN3-900051-07-0, URL http://www.R-project.org
Rodriguez LF (2006) Can invasive species facilitate native species? Evidence of how, when, and why these impacts occur. Biol Invasions 8:927–939
Settle WH, Wilson LT (1990) Invasion by the variegated leafhopper and biotic interactions: parasitism, competition, and apparent competition. Ecology 71:1461–1470
Shackelford CE, Brown RE, Conner RN (2000) Red-bellied woodpecker (Melanerpes carolinus) In: Poole A (ed) The Birds of North America Online. Cornell Lab of Ornithology, Ithaca. Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/650
Siegert NW, McCullough DG, Liebhold AM, Telewski FW (2009) Reconstruction of the establishment and spread of emerald ash borer through dendrochronological analysis. In: McManus K, Gottschalk KW (eds) Proceedings of 19th USDA interagency research forum on invasive species, Gen Tech Rep NRS-P-36 USDA Forest Service, Annapolis, MD, Northern Research Station, Newtown Square, PA, 70
Smith KG, Withgott JN, Rodewald PG (2000) Red-headed woodpecker (Melanerpes erythrocephalus). In: Poole A (ed) The Birds of North America Online. Cornell Lab of Ornithology, Ithaca. Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/518
Smith EL, Storer AJ, Roosien BK (2009) Emerald ash borer infestation rates in Michigan, Ohio, and Indiana. In: McManus K, Gottschalk KW (eds) Proceedings of 20th interagency research forum on invasive species, Gen Tech Rep NRS-P-51, Annapolis, MD, USDA Forest Service, Northern Research Station, Newtown Square, PA, p 96
States SL, Hochachka WM, Dhondt AA (2009) Spatial variation in an avian host community: implications for disease dynamics. EcoHealth 6:540–545
Strayer DL, Malcom HM (2007) Effects of zebra mussels (Dreissena polymorpha) on native bivalves: the beginning of the end or the end of the beginning? J North Am Benthol Soc 26:111–122
Wang X-Y, Yang Z-Q, Gould JR, Zhang Y-N, Liu G-J, Liu E (2010) The biology and ecology of the emerald ash borer, Agrilus planipennis, in China. J Insect Sci 10(128):1–23
Wells JV, Rosenberg KV, Dunn EH, Tessaglia-Hymes DL, Dhondt AA (1998) Feeder counts as indicators of spatial and temporal variation in winter abundance of resident birds. J Field Ornithol 69:577–586
Acknowledgments
We thank Project FeederWatch participants and donors for their continuing support and the reviewers for their comments. Incidental support came from NSF grant IOS-0918944 to WDK. Special thanks to Ben Zuckerberg for extracting the landcover data.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Koenig, W.D., Liebhold, A.M., Bonter, D.N. et al. Effects of the emerald ash borer invasion on four species of birds. Biol Invasions 15, 2095–2103 (2013). https://doi.org/10.1007/s10530-013-0435-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-013-0435-x