
Abstract
Observational studies of patterns of species invasions provide important insights into invasion processes at large spatial scales, leading to improved and focused efforts to prevent invasions. Nonnative species richness (NNSR), a common measure of ecosystem invasibility, varies widely among systems and is regulated by human activity and ecosystem characteristics such as habitat heterogeneity and climate. Patterns of NNSR may be confounded by variation in research effort, defined as the effort made to identify and catalogue invasions, and colonization pressure, defined as the number of species introduced. To explain spatial variation in ecosystem invasibility by fishes in the Mid-Atlantic region of the United States, we explored relationships among watershed characteristics, colonization pressure, research effort, and NNSR. Colonization pressure, research effort, and range in elevation were most predictive of differences in NNSR among river basins. Montane watersheds had greater NNSR, probably because of greater habitat heterogeneity due in part to human activity; widespread impoundments here created novel lentic habitats, while lotic environments were maintained elsewhere in these watersheds. Our results illustrate the importance of examining invasion patterns of specific taxa, within regions delineated by ecologically relevant criteria. Fish invasions in the Mid-Atlantic region seem to be regulated by different factors than in other regions of North America, demonstrating that invasive species management plans need to account for geographic variation among ecosystems. In the Mid-Atlantic region, reduction of novel, artificial environmental conditions (e.g., impoundments) in montane watersheds may reduce the invasibility of these ecosystems by generalist fish invaders.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Better understanding of the factors regulating the number of established nonnative species in an ecosystem (nonnative species richness [NNSR]; Blanchet et al. 2009) is needed to help prevent invasions, conserve biodiversity (Marchetti et al. 2004; Whittaker et al. 2005), and reduce social costs of biological invasions. Landscape-scale studies of nonnative species distributions can improve understanding of community assembly, because introductions provide large-scale observational evidence germane to assembly processes (Sax et al. 2007). Several observational studies have explored biotic, abiotic, and anthropogenic processes regulating ecosystem invasibility at landscape scales (e.g., Marchetti et al. 2004; Chiron et al. 2009). Observational studies of patterns in NNSR are important tools for understanding invasion processes at landscape scales (Fridley et al. 2007; Light and Marchetti 2007); however, confounding effects of colonization pressure (the number of species introduced to an ecosystem; Chiron et al. 2009; Lockwood et al. 2009) and data biases are not always adequately considered (Pyšek et al. 2008; Richardson and Pyšek 2008; Sol et al. 2008). To understand the mechanisms of species invasions at landscape scales, such confounding effects must be accounted for before ecological determinants of ecosystem invasibility are examined.
Biotic factors may have limited effect on invasibility at landscape scales. Native species richness is often positively related to NNSR across regions (Marchetti et al. 2004; Leprieur et al. 2008; Chiron et al. 2009; Davis 2009), suggesting that biotic acceptance is more common than biotic resistance at this scale. The biotic acceptance hypothesis posits that ecosystems with heterogeneous habitats meet the niche requirements of many species, thus supporting speciose native and nonnative faunas simultaneously (Fridley et al. 2007); whereas, the biotic resistance hypothesis suggests that ecosystems with greater native species richness will be less invasible because of fewer empty niches for introduced species to occupy (Elton 1958). At landscape scales, native communities are often subject to disturbance and do not necessarily occupy all available niches, leading to numerous opportunities for introduced species to establish (Fridley et al. 2007; Davis 2009). This scenario implies that biotic mechanisms have limited effect on invasibility at landscape scales.
Abiotic factors can contribute to ecosystem invasibility by providing conditions that generally support high species diversity. Ecosystems with greater habitat heterogeneity support higher NNSR because a wider range of species are more likely to encounter conditions that fulfill their ecological niche requirements (Davies et al. 2005). Environmental favorableness (lack of physicochemical extremes) can also contribute to invasibility (Fridley et al. 2007), and particular conditions at the time of an introduction can affect success (Moyle and Light 1996). The theory of fluctuating resource availability (Davis 2009) proposes that natural disturbances can free resources, providing opportunities for the establishment of nonnative species; however, frequent or extreme disturbance may inhibit establishment of most species (i.e., the intermediate disturbance hypothesis; Connell 1978; Davis 2009). Modification of natural disturbance regimes may play a stronger role in regulating invasibility than disturbance itself (Chytrý et al. 2008). In comparative studies of NNSR across landscapes, the role of measured abiotic factors is often secondary to factors representing human activity such as propagule pressure and anthropogenic disturbance (Marchetti et al. 2004; Leprieur et al. 2008; Chiron et al. 2009; Gassó et al. 2009); however, Blanchet et al. (2009) showed that abiotic characteristics were more strongly associated with fish NNSR than human activity in biogeographic realms with little economic development.
Ecosystem invasibility is expected to increase with anthropogenic disturbance (Lodge 1993; Moyle and Light 1996; Facon et al. 2006). Factors such as human population size, urban and agricultural land use, and access are associated with invasibility in terrestrial ecosystems (Chiron et al. 2009; Gassó et al. 2009). Many of these factors are also correlated with anthropogenic propagule pressure, which suggests that general human activity facilitates or accelerates invasions (Leprieur et al. 2008). In aquatic systems, alteration of hydrological regimes, land development, and human population density are correlated with NNSR at landscape scales (Gido and Brown 1999; Whittier and Kincaid 1999; Marchetti et al. 2004; Leprieur et al. 2008). Impoundment can greatly increase the invasibility of lotic systems by altering flow regimes and forming reservoirs, which are routinely stocked with nonnative fishes adapted to the novel, lentic conditions (Olden et al. 2006).
Observed patterns in NNSR across ecosystems depend in part on colonization pressure and research effort. Colonization pressure can be estimated from records of nonnative species that established and failed to establish reproducing populations (Chiron et al. 2009); however, available data on colonization pressure may have significant biases. Failed invasions are more difficult to detect than successful ones, especially in aquatic systems; thus, observed patterns of failed invasions may be particularly distorted by research effort. Reliable NNSR data are difficult to obtain at landscape scales (Pyšek et al. 2008) because research effort can vary spatially (Gassó et al. 2009). Detailed study of one ecosystem or area (e.g., near a university or museum of natural history) may result in better historic and contemporary collection records, and thorough faunal surveys by trained experts can identify additional nonnative species (Courtenay 2007). The effect of research effort on observed values of NNSR may be particularly great in areas of high endemism, where species are transplanted to nearby ecosystems and records of historic species distributions are patchy and incomplete. We are not aware of any landscape-scale study examining the contribution of research effort to observed patterns of NNSR.
Freshwater ecosystems offer ideal landscapes for the study of invasibility, acting as biogeographical islands because natural dispersal among systems is restricted (Gido and Brown 1999; Olden et al. 2010). Though large drainage basins may be separated, natural dispersal may be possible among sub-watersheds within a given basin, leading to similarities in NNSR. Additionally, nearby watersheds in separate drainage basins may have been subjected to similar historical management policies (e.g., game-fish stocking programs) or vectors of introduction associated with trade and commerce (e.g., pet releases). Such spatial correlation of NNSR can confound landscape-scale analyses (Sol et al. 2008; Chiron et al. 2009; Gassó et al. 2009).
Here, we examine the relative influences of abiotic, biotic, and human factors, as well as confounding variables representing colonization pressure and data quality, on patterns of nonnative fish species richness in watersheds in the Mid-Atlantic region of the United States, one of the most heavily invaded regions in the US (Nico and Fuller 1999). We explored relationships among natural watershed characteristics, anthropogenic disturbance, colonization pressure, research effort, and NNSR to understand the relative contribution of these factors to watershed invasibility. Based on previous studies of ecosystem invasibility, we predict that: (1) that descriptors of habitat heterogeneity and anthropogenic disturbance will have strong positive relationships with NNSR, and (2) biotic acceptance (i.e., positive relationships with native species richness) will be more evident than biotic resistance. We additionally hypothesize that, while controlling for colonization pressure and research effort, our proxies for these effects will have a positive association with NNSR. Our results will inform efforts to identify ecosystems that are most susceptible to invasion and thereby allow focus of prevention and management efforts.
Methods
Study area
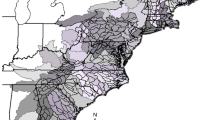
The study area included all eight-digit hydrologic units (HUC8s) delineated by the US Geological Survey (Seaber et al. 1987) in the Chesapeake Bay subregion (henceforth called the Mid-Atlantic region) of the Atlantic ichthyogeographic region (Fig. 1). Regions and subregions were defined by Edwards et al. (1998) based on similarity in native fish assemblages. The study area includes 11 HUC6 Atlantic-slope drainages, from the Susquehanna River in Pennsylvania and New York, south to the Neuse River in North Carolina. Nested within these are 80 HUC8s, two of which (Upper Chesapeake Bay, Lower Chesapeake Bay) were removed prior to analysis because they were almost entirely marine.

Distribution of nonnative species richness (NNSR) among HUC8s in the Mid-Atlantic region
Data collection
Nonnative fish distribution data were obtained from the USGS’s Nonindigenous Aquatic Species Database (NASD; nas.er.usgs.gov, accessed 17 September 2009). To reduce bias associated with lag times, we included only species recorded as established by 1995. A decline in the number of records per year after 1995 was observed for the Mid-Atlantic region in the NASD, and only four species were reported as established after this date (Lapointe 2010). The number of records peaked from the 1970s to 1990s; however, these records likely represent introductions that occurred in earlier decades, when sport and forage fishes were more widely stocked (Jackson et al. 2004). Lag times likely exist for more recent introductions because of delays in detecting, reporting, and incorporating records of nonnative species in large-scale databases (Keller et al. 2009). We calculated NNSR as the total number of nonnative species established in each HUC8. Freshwater and diadromous species were included, while marine species were excluded. One hybrid (Morone chrysops x saxatilis) was included as an established species because of the potential to interbreed with native species in some drainages.
For a proximate measure of colonization pressure, we calculated the number of nonnative species that failed to establish in each HUC8. This included species listed in the NASD as collected (but not known to have reproduced), stocked [persist because of repeated stockings) or failed (not collected after an unspecified amount of time after introduction; definitions provided in Shafland et al. (2008)]. Lockwood et al. (2009) pointed out that patterns in NNSR cannot be analyzed as a function of colonization pressure because NNSR must be included in the calculation of colonization pressure itself. They recommend accounting for colonization pressure by using the proportion of species established, rather than the actual number of species established, as response variable; however, many watersheds in the Mid-Atlantic region had very few established species and failed species. Thus, many watersheds had extreme values of either 0 or 100 % of species established with values based on few records of introductions. Instead, we used the number of failed invaders as a proximate measure of colonization pressure, because this measure was independent of the number of established species.
We used ArcGIS 9.2 (ESRI Inc. Redlands, CA, USA) to compile data on ecosystem characteristics for each HUC8. Variables were related to land cover (proportion of land forested, developed, or in agriculture), habitat availability (proportion of land cover as lakes, reservoirs, and wetlands, density of small, large, and artificial streams), climate (minimum January temperature, mean annual precipitation), physical characteristics (watershed area, range in elevation), and other anthropogenic characteristics (human population density, number of dams; Table 1). Native species richness was defined as the total number of native species in each watershed (excluding marine species), as listed in NatureServe Explorer (http://www.natureserve.org). We obtained the Universal Transverse Mercator coordinates for the centroid of each watershed, and calculated Euclidean distance (km) between all pairs of HUC8 centroids.
Detailed examination of NASD records and the NatureServe Explorer list of native species distributions showed that the nonnative status of several species was in dispute. The NASD provides information on the distribution of nonnative aquatic species in the United States, based on personal communications and published reports (nas.er.usgs.gov/about/default.aspx). NatureServe Explorer provides similar information on the distribution of native species, based on published records and consultation with scientists in their natural heritage program (http://www.natureserve.org/explorer/aboutd.htm). Several species were listed as both native and nonnative in a given watershed by these two databases, and these occurrences appeared to be related to research effort. Thirty-eight species were listed as native to some parts of the region but transplanted to at least one other HUC8 in the region. For 25 % (55 records) of the NASD records by HUC8 for these species, NatureServe also listed the species as native to that HUC8. We found the citations for each of these records in the NASD, and reviewed the rationale in each citation for considering these species nonnative. Most of the shared records represented disputes about historic distributions, but were included in the NASD because researchers [primarily Jenkins and Burkhead (1994) but also Menhinick (1991); Starnes (2002)] provided detailed examinations of historic and contemporary collection records for particular watersheds. These examinations provided evidence that one or more species were not native to a given watershed even though they were generally considered native there; however such detailed reviews were not available for all watersheds in the region. As such, we interpreted the number of shared records (i.e., species listed as both native and nonnative in a HUC8) as a proximate measure of the amount of effort put forth by researchers to document and compare historic and contemporary collection records. We include this variable to control for the confounding effect of research effort on other results. If this variable is in fact unrelated to research effort, it will decrease the statistical efficiency of our analysis; on the other hand, if it is related to research effort it will serve to control for difference in research effort on other results.
Analytic methods
To explore relationships between ecosystem characteristics and NNSR, we used hierarchical partitioning (HP; Chevan and Sutherland 1991) and a Poisson error distribution to identify which explanatory variables had a significant, independent influence on nonnative species richness. Hierarchical partitioning assesses variable importance by partitioning variance explained into independent and joint contributions for all possible combinations of explanatory variables, and is useful for identifying variable importance rather than constructing a parsimonious predictive model (Pont et al. 2005). A Poisson distribution was used (instead of for instance a normal distribution in a conventional linear model) because the response was discrete-valued and bounded at zero; the Poisson distribution also models a process composed of infrequent but independent events (Zuur et al. 2009) such as is likely for species invasions. Hierarchical partitioning requires computation of N! models (for N variables); thus, we first needed to reduce the set of explanatory variables to allow a computationally tractable problem. We did this using Akaike information criterion (AIC) stepwise model selection, as available in the package MASS (Venables and Ripley 2002), and a Poisson-distributed generalized linear model (GLM). We then implemented HP using the HIER.PART package (Mac Nally and Walsh 2004; Walsh and Mac Nally 2008) within R (R Development Core Team 2009) on the selected variables. We tested statistical significance for variable importance with randomization techniques (Mac Nally 2002) at 95 % confidence.
To describe the direction of correlations and explore the sensitivity of HP to non-linear relationships, we applied generalized additive models (GAMs; Hastie and Tibshirani 1986). We implemented GAMs (using thin-plate regression splines) by including both Poisson and negative-binomial error distributions and all explanatory variables in the MGCV package (Wood 2008) within R. The negative binomial model was fitted to explore the potential impact of overdispersion, which can be caused by missing explanatory variables (Lindén and Mäntyniemi 2011). We configured GAMs to automatically estimate a parsimonious degree of nonlinearity from generalized cross-validation (Craven and Wahba 1979), and allowed automatic model building to penalize a variable out of the model (i.e., estimate an effect that was zero for all values of the independent variable). We then applied the AIC to final, fitted GAM models to select between Poisson and negative-binomial GAMs.
To explore the sensitivity of plausible spatial effects on variable selection and interpretation we also applied a generalized linear mixed model (GLMM) using a Poisson error distribution. This model included random effects for each HUC6 and a spherical semivariogram by latitude and longitude. The random-effects structure accounted for dispersal of nonnative species to nearby watersheds, which could lead to similarities in NNSR among HUC8s within a given HUC6; the semivariogram estimated correlations between HUC8s based on linear distance, such as would be caused by changes in NNSR in HUC8s due to similar propagule pressure from shared management histories or nearby human activities. We estimated random effects using restricted maximum likelihood, which precluded further use of deviance-based model selection criteria such as AIC or Bayesian information criterion, in the NLME package (Pinheiro et al. 2009). Instead, based on a Wald test, we performed backwards model selection for all fixed effects (starting at the full model), with a significance threshold of 0.15 for all retained variables.
Different analytical methods capture different types of effect so that, predictably, each method will attribute observed differences among HUC8s to different explanatory variables. In particular, HP assumes a linear effect for quantitative variables, while GAMs allow for nonlinear effects, and GLMMs allow for spatial correlation. No analytical approach is ideal for describing landscape patterns of invasibility; however, several variables were routinely important, suggesting that our principal findings are robust to limitations of analytical approaches and effects of spatial correlation. Variables selected in only one model were interpreted to likely represent spurious associations (particularly when effects were weak). Thus, we focus our interpretation on variables that showed strong effects in at least two models.
Results
There were 69 nonnative species documented in the study region, with a maximum of 21 nonnative species in a HUC8. The least-invaded HUC8s were in the southern coastal plain, east of Chesapeake Bay, and in central Pennsylvania; five had no nonnative species. HUC8s with the greatest number of invaders included the Upper Roanoke, Middle Roanoke, Upper James, and Upper Dan in central and southern Virginia and North Carolina, and the Middle Potomac-Catoctin, Middle Potomac-Anacostia-Occoquan, and Rapidan-Upper Rappahanock in northern Virginia and Maryland. The most widely-distributed nonnative species included bluegill (Lepomis macrochirus; 66 % of HUC8s), common carp (Cyprinus carpio; 62 %), largemouth bass (Micropterus salmoides; 59 %), smallmouth bass (Micropterus dolomieu; 58 %), and rock bass (Ambloplites rupestris; 47 %).
Stepwise model selection filtered our original fourteen variables to nine for the HP analysis. Of these, six were statistically significant based on randomization methods, and explained the majority (91 %) of independent effects on NNSR (Table 2). Variables describing research effort and colonization pressure were most important, followed by range in elevation and proportion of land forested. Of the variables related to anthropogenic disturbance, only human population density and proportion of land in agriculture had a significant independent effect on NNSR, though these were minor (~5 % each). Other variables related to habitat availability and native species richness were identified by stepwise model selections, but did not explain a significant portion of the independent effects identified in HP.
Between Poisson and negative-binomial GAMs, the Poisson distribution was selected by AIC, implying a lack of overdispersion. The GAM explained 70.4 % of the deviance in NNSR and confirmed strong effects of range in elevation and proportion of land forested but the effect of the number of failed invasions as a measure of colonization pressure was weaker and the measure of research effort was not selected (Table 2). Human population density, proportion of land developed, native species richness, and variables describing habitat availability also had strong effects. Most variables were positively correlated with NNSR at low values but demonstrated thresholds beyond which the relationship weakened or became negative (Fig. 2). The density of small and large streams was negatively correlated with NNSR.

Partial dependence plots of relationships between explanatory variables and NNSR (n = 78) from GAM analysis, showing the predicted relationship between NNSR and each explanatory variable, while accounting for the effects of all other variables. Explanatory variables (explained in Table 1) are shown on the x-axes. Y-axes represent the difference between actual and predicted (i.e., residual) NNSR. Solid lines represent the estimated relationship, and dashed lines represent 95 % confidence intervals. Residual values for each HUC8 are marked by ‘x’s, and hash marks above the x-axes represent the location of each observation
Several factors were associated with NNSR across HUC8s in the GLMM models. The standard deviation for random effects among HUC6s was ~1, suggesting that HUC8s nested within a HUC6 are more likely to have similar NNSR, and that nearby HUC8s have similar NNSR. The effects of geographic distance on NNSR similarity decreased until HUC8 centroids were >185 km apart, after which no spatial correlation was detected. A nine-variable model was identified by AIC for the linear model after accounting for spatial effects. The lower bound on the percent of deviance explained by the GLM was 86.6 %, based on calculations for a Poisson mixed-effect model without the semivariogram (this lower bound is presented due to difficulties in defining a deviance explained for a GLMM with a semivariogram).
Measures of colonization pressure, research effort, and range in elevation remained important but the association of NNSR with proportion of land forested was negative and not significant. Native species richness was positively associated with NNSR and the density of small and large streams was negatively correlated with NNSR. The proportion of land cover as wetlands and density of artificial streams were associated with NNSR, though neither was selected by other analytical approaches.
Discussion
In this study, we demonstrate the importance of natural abiotic characteristics and anthropogenic disturbance in structuring patterns of NNSR across landscapes, after accounting for the confounding effects of research effort and colonization pressure. Previous studies have stressed that colonization pressure must be considered when examining large-scale invasion patterns (Chiron et al. 2009; Lockwood et al. 2009), though few studies quantitatively explore the effect of research effort (but see Moerman and Estabrook 2006; Pautasso and McKinney 2007). In contrast to other large-scale studies where human activity such as hydrologic alteration of aquatic systems appears to be the main determinant of ecosystem invasibility (e.g., Marchetti et al. 2004; Leprieur et al. 2008), we found that invasion patterns were driven by a combination of natural abiotic factors and anthropogenic disturbance in the Mid-Atlantic region.
Effects of research effort and colonization pressure on observed NNSR patterns
Understanding and accounting for biases in large-scale invasion datasets is challenging yet necessary. Varying levels of research effort can introduce errors into NNSR datasets, confounding relationships between NNSR and ecosystem characteristics. The presence of institutions of higher learning in a county has been used as a surrogate for research effort (Pautasso and McKinney 2007); however, research often occurs beyond the immediate environs of a particular institution. Here, close examination of the NASD database, its sources, and the specific details surrounding certain questionable records revealed a likely ‘ichthyologist effect,’ similar to the ‘botanist effect’ suggested by Moerman and Estabrook (2006), whereby the number of nonnative species documented as established in a watershed depends in part on the degree of historic and contemporary ichthyological knowledge there. Contemporary research effort and the availability of historical records are not uniform across the Mid-Atlantic region (Starnes 2002), leading to uncertainties in the ranges of some nonnative species whose native ranges are poorly understood. Other large-scale databases and checklists of nonnative species (e.g., DAISIE in Europe; Vilà et al. 2010) likely suffer from similar biases, which should be considered when analyzing regional patterns of invasibility (Pyšek et al. 2008; Sol et al. 2008). The effect on analyses of not compensating for research effort could range from innocuous to severe, depending upon whether research effort is correlated with other explanatory variables.
To complement the colonization pressure hypothesis (Chiron et al. 2009; Lockwood et al. 2009), we suggest a research effort hypothesis: large-scale patterns of NNSR are biased by geographic variation in research effort to document species distributions, especially historic efforts prior to widespread introductions. This hypothesis is relevant at both regional and global scales, and may be particularly important in regions with high endemism. In such regions, species transplanted between adjacent drainage basins may be identified as nonnative only if detailed knowledge of their historical native range exists. Previous studies demonstrating positive relationships between measures of human activity such as gross domestic product (GDP) or international trade (e.g., Leprieur et al. 2008; Westphal et al. 2008) are particularly confounded by research effort. At global scales, wealthier countries or regions have greater resources to survey native biota and to monitor and record invasions (Westphal et al. 2008), making it unclear whether the higher NNSR observed in nations with high GDP is a function of increased colonization pressure or invasibility or simply an effect of measurement error. Within regions such as the Mid-Atlantic, wealth and trade may not vary enough to be important drivers of invasions patterns, yet research effort can still vary considerably. At all spatial scales, failure to account for biases or errors in invasion records can lead to erroneous conclusions about ecosystem invasibility (Sol et al. 2008).
Colonization pressure was also a strong determinant of NNSR, even though a proximate measure (i.e., the number of failed invasions) was used. Failed invasions are difficult to detect (Sol et al. 2008), especially when species are introduced accidentally (Diez et al. 2009), leading to considerable uncertainty in records of failed invasions. Thus, the veracity of records of failed invaders is particularly sensitive to research and monitoring efforts. We acknowledge that the number of failed invasions could be viewed as a measure of ecosystem resistance to invasion (Lockwood et al. 2009); however, the strong association with the number of established non-native species provides evidence that, in this dataset, the number of failed invasions is probably associated with colonization pressure. Sufficient information does not exist to evaluate the relative contribution of research effort, colonization pressure, and invasibility to observed patterns of failed invasions; however, controlling for failed invasions accounted for one confounding effect while the roles of anthropogenic disturbance and biotic/abiotic factors in regulating NNSR were assessed.
Biotic and abiotic factors as determinants of NNSR
After accounting for measures of research effort and colonization pressure, abiotic factors explained much of the variation in NNSR among watersheds. Notably, range in elevation and proportion of land forested were most important, while variables describing anthropogenic disturbance (e.g., human population density) had minor or no effects. Range in elevation and proportion of land forested were correlated (Spearman r = 0.65), and were greatest in montane watersheds in the western part of the region. Downstream watersheds in the coastal plain had low range in elevation, little forested land, and lower NNSR. In contrast with other studies of landscape-scale patterns of fish NNSR (Gido and Brown 1999; Marchetti et al. 2004), watershed area and native species richness were not major factors. Watershed area probably had little effect because HUC8s had limited variance in area. Conversely, the limited effect of native species richness on NNSR could not be explained by lack of contrast among HUC8s, because native species richness varied considerably among watersheds (20–76 species/HUC8).
Patterns of NNSR in the Mid-Atlantic region likely result from an interaction between anthropogenic and natural abiotic factors. The limited importance of anthropogenic factors in analysis results, particularly for HP, was unexpected because invasibility of freshwater systems is widely considered to increase with anthropogenic disturbance (Gido and Brown 1999; Whittier and Kincaid 1999; Marchetti et al. 2004; Leprieur et al. 2008), especially in wealthy regions (Blanchet et al. 2009). Instead, range in elevation was found to be the most important ecosystem characteristic in explaining NNSR. In the Mid-Atlantic region, low-elevation lotic systems provide warm, often turbid, nutrient-rich conditions and support generalist species, whereas montane streams support specialist species adapted to cool, clear, nutrient-poor waters (Scott and Helfman 2001). Watersheds with large range in elevation naturally contain a wide variety of habitats suitable for colonization by a diversity of invaders (Davies et al. 2005; Leprieur et al. 2008). Yet even here, anthropogenic disturbance may contribute to increasing ecosystem invasibility. Moderate anthropogenic disturbance to montane streams can increase temperature, sediment, and nutrient loads (e.g., through deforestation) and create lentic habitats (e.g., through impoundment), all providing novel conditions suitable for generalist nonnative species (Angermeier and Winston 1998). Many species native to downstream sections were stocked in montane reservoirs after upstream sections were impounded (Jenkins and Burkhead 1994).
Effects of anthropogenic disturbance on ecosystem invasibility seem to depend on the ecological context, especially the extent to which human alterations make available habitat more suitable for a wide array of potential invaders. Anthropogenic disturbance is not unique to watersheds with large range in elevation but relations between specific types of disturbance and natural conditions can vary spatially. For example, although the number of dams and range in elevation were not correlated (Spearman r = 0.13), damming appear to have a greater effect on the invasibility of montane watersheds compared to coastal plain watersheds because the resulting lentic habitats are novel and widely suitable. Conversely, the habitat diversity of coastal plain watersheds has not been equally increased by human alteration. Many coastal plain streams and swamps are naturally harsh because of acidity, low dissolved oxygen, and dystrophic conditions which limit their suitability for many potential invaders (Smock and Gilinsky 1992).
The strong positive relationship between range in elevation and NNSR remained even after accounting for significant spatial correlation in NNSR among watersheds in the region, both within and across larger drainage basins. Such correlation likely reflects similarities in both abiotic characteristics and colonization pressure. Nearby watersheds are generally found in the same physiographic province, with comparable ranges in elevation and corresponding habitat heterogeneity. Colonization pressure is probably related to management practices, angling habits, and regulatory frameworks controlling vectors of introduction (e.g., baitfish, aquaculture, pet trade), most of which follow political rather than watershed boundaries. In addition, spatial variation in colonization pressure occurs at a coarser, drainage-basin scale when nonnative species disperse (or are transplanted) to connected watersheds. In the Mid-Atlantic region, numerous impoundments restrict upstream movement among watersheds; however, individuals can disperse downstream through turbines and spillways (Schmetterling and McFee 2006; Williams 2008). Thus, downstream watersheds may be subjected to particularly high colonization pressure because species introduced to montane areas may disperse downstream. That coastal plain watersheds generally had lower NNSR despite this likely increase in colonization pressure provides further evidence that abiotic factors (e.g., harsh conditions including low pH and dissolved oxygen in coastal plain watersheds) play a significant role in determining invasibility.
Non-linear effects and negative relationships with particular habitat types provided further evidence that habitat heterogeneity and suitability drive ecosystem invasibility. Range in elevation and proportion of land forested both exhibited thresholds beyond which NNSR did not increase. At a certain point, large range in elevation and high forest cover may reflect steep channel gradients, little disturbance and, therefore, little warm, lentic, nutrient-rich habitat and low habitat diversity. Similarly, NNSR increased with human population density but decreased at high density, supporting the intermediate disturbance hypothesis. Habitat quality and diversity can decrease with high human population densities (Nelson et al. 2009), limiting species establishment even if colonization pressure is high. NNSR also decreased as the density of either small or large streams increased. Increases of one such habitat likely reflect decreases in other habitats, i.e., reduced habitat heterogeneity. Finally, native species richness had a weak positive correlation with NNSR, probably reflecting similar responses between native and nonnative species along gradients of habitat heterogeneity (Davies et al. 2005).
Biotic resistance and acceptance
The weak relationship between native species richness and NNSR provided little support for either the biotic resistance or biotic acceptance hypotheses (Leprieur et al. 2008), and biotic characteristics appeared to have little effect on ecosystem invasibility at watershed scales. Native species richness probably has little effect on landscape-scale invasibility, and positive relationships between native and nonnative species richness likely reflect similarities in response to habitat heterogeneity (Fridley et al. 2007). Analyses of functional diversity and complementarity may be more relevant to assessments of biotic resistance, given that competition with native species may be low for nonnative species with novel traits (Olden et al. 2006). However, analyses of biotic effects are complicated by differences in the communities encountered by each invader. Early invaders primarily compete with native species, whereas successive invaders also compete with established nonnative species. Similarly, extirpations may alter the communities encountered by successive invaders. Our results show that native species richness is a weak predictor of nonnative species richness in the Mid-Atlantic region and suggest that the recent focus on such relationships (e.g., Leprieur et al. 2008; Blanchet et al. 2009) may be unwarranted in some ecosystems.
Relevance to other regions
Our results differ considerably from similar studies in other regions, suggesting that drivers of ecosystem invasibility vary among regions. Blanchet et al. (2009) found that drivers of ecosystem invasibility varied globally among biogeographical realms. Several studies now show that drivers of ecosystem invasibility by fishes differ considerably among regions within a continent: drainage area in Mississippi basin watersheds (Gido and Brown 1999), altered hydrology and watershed disturbance in California watersheds (Marchetti et al. 2004), and range in elevation in Mid-Atlantic watersheds (present study). Such differences may reflect regional variation in habitat conditions, species composition, and historical patterns of biogeography and human activity. For example, Mid-Atlantic watersheds have a unique evolutionary history, which has led to relatively depauperate native assemblages with a high degree of endemism. This creates the possibility for a large number of species to be transplanted among nearby watersheds, leading to unique patterns of invasions. Furthermore, the differences in habitats between coastal plain and montane watersheds in the Mid-Atlantic region do not exist in the Mississippi Basin watersheds examined by Gido and Brown (1999). Each regional-scale study (Gido and Brown 1999; Marchetti et al. 2004), including this study, used different variables and analytical approaches. It is unclear whether observed differences in patterns of invasibility among these regions are artifacts of different analytical approaches or reflect real variation in natural or anthropogenic factors. We suspect that both explanations play a role. Each of these studies contributes to a general conceptual model of invasibility, but there is a need for caution in generalizing the drivers of invasibility across scales, regions, and taxa; results from one study are unlikely to transfer directly to other systems. Future studies could directly assess the differences among regions by analyzing similar data from multiple regions. We suggest that efforts to identify especially invasible ecosystems and prevent future invasions need to be regionally focused to appropriately inform conservation strategies.
NNSR as a measure of ecosystem invasibility
Though NNSR is commonly used as the primary measure of ecosystem invasibility (e.g., Gido and Brown 1999; Chiron et al. 2009), there are several limitations to this measure beyond biases associated with colonization and research effort. The actual number of nonnative species may be a less relevant measure of ecosystem invasibility than their relative abundance or biomass; however, such measures can be difficult to obtain at landscape scales. The identity of nonnative species is at least as important as species richness in understanding ecosystem invasibility, as two ecosystems with equal NNSR may contain completely different species. No ecosystem is equally invasible by all nonnative species, as establishment depends fundamentally on a match between environment and the ecological niche requirements of a given species (Moyle and Light 1996), and consideration of both nonnative species richness and identity can improve understanding of ecosystem invasibility. In the Mid-Atlantic region, nonnative communities can be sorted along gradients in temperature and range in elevation (Lapointe and Light 2012). Range in elevation largely affected NNSR with fewer established species in coastal plain watersheds; however, the type of species established here also differed from montane regions. Additionally, temperature affected the type of species that established, and northern watersheds contained different nonnative communities from southern watersheds, even when NNSR values were similar (Lapointe and Light 2012). Thus, improved understanding of ecosystem invasibility can be obtained by considering both nonnative species richness and identity, and multi-scale analyses that incorporate data on relative abundance may provide further insight.
Conclusions
Patterns of NNSR appear to be driven by natural ecosystem characteristics, human disturbance, and colonization pressure; however, biases in data quality must be accounted for before underlying mechanisms are understood. Our results primarily support hypotheses relating to colonization pressure and habitat heterogeneity, with human activity leading to invasions by increasing both factors. Our results also illustrate the importance of examining invasion patterns of specific taxa, within regions delineated by ecologically relevant criteria (Moyle and Marchetti 2006; Hayes and Barry 2008). Different factors likely regulate terrestrial invasions across regions, and fish invasions in the Mid-Atlantic region appear to be regulated differently than in previously studied regions (e.g., Marchetti et al. 2004).
In particular, we demonstrated how underlying abiotic characteristics mediated differential effects of anthropogenic disturbance on ecosystem invasibility. Montane watersheds can support a wider range of nonnative species (and thus higher NNSR) because anthropogenic disturbance has increased habitat heterogeneity there. Anthopogenic disturbance of coastal plain watersheds did not lead to high invasibility; however, invasibility of these watersheds may still be high for species with appropriate environmental tolerances (Moyle and Light 1996). Species niche requirements are important considerations in understanding invasibility and predicting future invasions.
These findings can aid in managing invasions in the Mid-Atlantic region, and suggest that management plans should account for abiotic conditions. Coastal plain systems can be more effectively managed by focusing prevention efforts on species most likely to tolerate warm, turbid conditions. Targeted prevention efforts in montane watersheds are more difficult, given the range of available habitats. Instead, reduction of novel conditions created by anthropogenic activities (e.g., impoundments) may reduce the invasibility of montane ecosystems by generalist invaders.
References
Angermeier PL, Winston MR (1998) Local vs. regional influences on local diversity in stream fish communities of Virginia. Ecology 79:911–927
Blanchet S, Leprieur F, Beauchard O, Staes J, Oberdorff T, Brosse S (2009) Broad-scale determinants of non-native fish species richness are context-dependent. Proc R Soc B Biol Sci 276:2385–2394
Chevan A, Sutherland M (1991) Hierarchical partitioning. Am Stat 45:90–96
Chiron F, Shirley S, Kark S (2009) Human-related processes drive the richness of exotic birds in Europe. Proc R Soc B Biol Sci 276:47–53
Chytrý M, Jarošik V, Pyšek P, Hájek O, Knollová I, Tichý L, Danihelka J (2008) Separating habitat invasibility by alien plants from the actual level of invasion. Ecology 89:1541–1553
Connell JH (1978) Diversity in tropical rain forests and coral reefs. Science 199:1302–1310
Courtenay WR Jr (2007) Introduced species: what do you have and how do you know? Trans Am Fish Soc 136:1160–1164
Craven P, Wahba G (1979) Smoothing noisy data with spline functions: estimating the correct degree of smoothing by the method of generalized cross-validation. Numer Math 31:377–403
Davies KF, Chesson P, Harrison S, Inouye BD, Melbourne BA, Rice KJ (2005) Spatial heterogeneity explains the scale dependence of the native-exotic diversity relationship. Ecology 86:1602–1610
Davis MA (2009) Invasion biology. Oxford Univeristy Press, Inc., New York
Diez JM, Williams PA, Randall RP, Sullivan JJ, Hulme PE, Duncan RP (2009) Learning from failures: testing broad taxonomic hypotheses about plant naturalization. Ecol Lett 12:1174–1183
Edwards C, Hill D, Maxwell J (1998) Aquatic zoogeography of North America (nearctic zone). U.S.D.A. Forest Service, Rhinelander
Elton CS (1958) The ecology of invasions by animals and plants. Chapman & Hall, London
Facon B, Genton BJ, Shykoff J, Jarne P, Estoup A, David P (2006) A general eco-evolutionary framework for understanding bioinvasions. Trends Ecol Evol 21:130–135
Fridley JD, Stachowicz JJ, Naeem S, Sax DF, Seabloom EW, Smith MD, Stohlgren TJ, Tilman D, Von Holle B (2007) The invasion paradox: reconciling pattern and process in species invasions. Ecology 88:3–17
Gassó N, Sol D, Pino J, Dana ED, Lloret F, Sanz-Elorza M, Sobrino E, Vilà M (2009) Exploring species attributes and site characteristics to assess plant invasions in Spain. Divers Distrib 15:50–58
Gido KB, Brown JH (1999) Invasion of North American drainages by alien fish species. Freshw Biol 42:387–399
Hastie T, Tibshirani R (1986) Generalized additive models. Stat Sci 1:297–318
Hayes KR, Barry SC (2008) Are there any consistent predictors of invasion success? Biol Invasions 10:483–506
Jackson JR, Boxrucker JC, Willis DW (2004) Trends in agency use of propagated fishes as a management tool in inland fisheries. In: Nickum MJ, Mazik PM, Nickum JG, MacKinlay DD (eds) Propagated fish in resource management. American Fisheries Society, Symposium 44, Bethesda, pp 121–138
Jenkins RE, Burkhead NM (1994) Freshwater fishes of Virginia. American Fisheries Society Bethesda, Maryland
Keller RP, Ermgassen P, Aldridge DC (2009) Vectors and timing of freshwater invasions in great Britain. Conserv Biol 23:1526–1534
Lapointe NWR (2010) Fish invasions in the Mid-Atlantic region of the United States. Doctoral dissertation, Virginia Polytechnic Institute, Blacksburg
Lapointe NWR, Light T (2012) Landscape-scale determinants of nonnative fish communities. Divers Distrib 18:182–293
Leprieur F, Beauchard O, Blanchet S, Oberdorff T, Brosse S (2008) Fish invasions in the world’s river systems: when natural processes are blurred by human activities. PLoS Biol 6:404–410
Light T, Marchetti MP (2007) Distinguishing between invasions and habitat changes as drivers of diversity loss among California’s freshwater fishes. Conserv Biol 21:434–446
Lindén A, Mäntyniemi S (2011) Using the negative binomial distribution to model overdispersion in ecological count data. Ecology 92:1414–1421
Lockwood JL, Cassey P, Blackburn TM (2009) The more you introduce the more you get: the role of colonization pressure and propagule pressure in invasion ecology. Divers Distrib 15:904–910
Lodge DM (1993) Biological invasions—lessons for ecology. Trends Ecol Evol 8:133–137
Mac Nally R (2002) Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables. Biodivers Conserv 11:1397–1401
Mac Nally R, Walsh CJ (2004) Hierarchical partitioning public-domain software. Biodivers Conserv 13:659–660
Marchetti MP, Light T, Moyle PB, Viers JH (2004) Fish invasions in California watersheds: testing hypotheses using landscape patterns. Ecol Appl 14:1507–1525
Menhinick EF (1991) The freshwater fishes of North Carolina. North Carolina Wildlife Resources Commission, Raleigh
Moerman DE, Estabrook GF (2006) The botanist effect: counties with maximal species richness tend to be home to universities and botanists. J Biogeogr 33:1969–1974
Moyle PB, Light T (1996) Biological invasions of fresh water: empirical rules and assembly theory. Biol Conserv 78:149–161
Moyle PB, Marchetti MP (2006) Predicting invasion success: freshwater fishes in California as a model. Bioscience 56:515–524
Nelson KC, Palmer MA, Pizzuto JE, Moglen GE, Angermeier PL, Hilderbrand RH, Dettinger M, Hayhoe K (2009) Forecasting the combined effects of urbanization and climate change on stream ecosystems: from impacts to management options. J Appl Ecol 46:154–163
Nico LG, Fuller PL (1999) Spatial and temporal patterns of Nonindigenous fish introductions in the United States. Fisheries 24:16–27
Olden JD, Poff NL, Bestgen KR (2006) Life-history strategies predict fish invasions and extirpations in the Colorado River basin. Ecol Monogr 76:25–40
Olden JD, Kennard MJ, Leprieur F, Tedesco PA, Winemiller KO, García-Berthou E (2010) Conservation biogeography of freshwater fishes: recent progress and future challenges. Divers Distrib 16:496–513
Pautasso M, McKinney ML (2007) The botanist effect revisited: plant species richness, county area, and human population size in the United States. Conserv Biol 21:1333–1340
Pinheiro J, Bates D, DebRoy S, Sarkar D, The R Core Team (2009) nlme: linear and nonlinear mixed effects models. R package version 3.1–96
Pont D, Hugueny B, Oberdorff T (2005) Modelling habitat requirement of European fishes: do species have similar responses to local and regional environmental constraints? Can J Fish Aquat Sci 62:163–173
Pyšek P, Richardson DM, Pergl J, Jarošik V, Sixtová Z, Weber E (2008) Geographical and taxonomic biases in invasion ecology. Trends Ecol Evol 23:237–244
R Development Core Team (2009) R: a language and environment for statistical computing. Austria, Vienna
Richardson DM, Pyšek P (2008) Fifty years of invasion ecology—the legacy of Charles Elton. Divers Distrib 14:161–168
Sax DF, Stachowicz JJ, Brown JH, Bruno JF, Dawson MN, Gaines SD, Grosberg RK, Hastings A, Holt RD, Mayfield MM, O’Connor MI, Rice WR (2007) Ecological and evolutionary insights from species invasions. Trends Ecol Evol 22:465–471
Schmetterling DA, McFee JA (2006) Migrations by fluvial largescale suckers (Catostomus macrocheilus) after transport upstream of Milltown Dam, Montana. Northwest Sci 80:18–25
Scott MC, Helfman GS (2001) Native invasions, homogenization, and the mismeasure of integrity of fish assemblages. Fisheries 26:6–15
Seaber PR, Kapinos FP, Knapp GL (1987) Hydrologic unit maps
Shafland PL, Gestring KB, Stanford MS (2008) Categorizing introduced fishes collected from public waters. Southeast Nat 7:627–636
Smock LA, Gilinsky E (1992) Coastal plain blackwater streams. In: Hackney CT, Adams SM, Martin WH (eds) Biodiversity of the southeastern United States: aquatic communities. Wiley, New York, pp 271–313
Sol D, Vilà M, Kuhn I (2008) The comparative analysis of historical alien introductions. Biol Invasions 10:1119–1129
Starnes WC (2002) Current diversity, historical analysis, and biotic integrity of fishes in the lower Potomac basin in the vicinity of Plummers Island, Maryland-Contribution to the natural history of Plummers Island, Maryland XXVIIx. Proc Biol Soc Wash 115:273–320
Venables WN, Ripley BD (2002) Modern applied statistics with S, 4th edn. Springer, New York
Vilà M, Basnou C, Pyšek P, Josefsson M, Genovesi P, Gollasch S, Nentwig W, Olenin S, Roques A, Roy D, Hulme PE (2010) How well do we understand the impacts of alien species on ecosystem services? a pan-European, cross-taxa assessment. Front Ecol Environ 8:135–144
Walsh C, Mac Nally R (2008) heir.part: hierarchical partitioning. R package version 1.0-3
Westphal MI, Browne M, MacKinnon K, Noble I (2008) The link between international trade and the global distribution of invasive alien species. Biol Invasions 10:391–398
Whittaker RJ, Araujo MB, Paul J, Ladle RJ, Watson JEM, Willis KJ (2005) Conservation biogeography: assessment and prospect. Divers Distrib 11:3–23
Whittier TR, Kincaid TM (1999) Introduced fish in northeastern USA lakes: regional extent, dominance, and effect on native species richness. Trans Am Fish Soc 128:769–783
Williams JG (2008) Mitigating the effects of high-head dams on the Columbia River, USA: experience from the trenches. Hydrobiologia 609:241–251
Wood SN (2008) Fast stable direct fitting and smoothness selection for generalized additive models. J R Stat Soc Series B Stat Methodol 70:495–518
Zuur AF, Leno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer Science+Business Media, LLC., New York
Acknowledgments
We thank Rich Pendleton and Tye Deweber for help with data compilation. Many others provided help and advice, including Scott Klopfer and Michael Marchetti. We thank Amy Benson (USGS) for providing NASD data. Theo Light inspired us to explore the importance of research effort as a determinant of landscape-scale patterns of NNSR. The manuscript was greatly improved by comments from Walter Courtenay, Jr., Andrew Dolloff, Emili García-Berthou, Theo Light, Brian Murphy, André Punt, and anonymous reviewers. This research was supported by the U.S. Geological Survey Cooperative Fish and Wildlife Research Unit at Virginia Tech, a grant from the U.S. Fish and Wildlife Service Mid-Atlantic Panel on Aquatic Invasive Species, and a Natural Science and Engineering Research Council of Canada post-graduate fellowship. Partial funding for JTT came from the University of Washington, a NMFS groundfish project grant to the University of Washington, and a National Marine Fisheries Service Sea Grant Population Dynamics Fellowship (NA09OAR4170120). Any use of trade, product, or firm names does not imply endorsement by the U.S. government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Virginia Cooperative Fish and Wildlife Research Unit is jointly supported by US Geological Survey, Virginia Polytechnic Institute and State University, Virginia Department of Game and Inland Fisheries, and Wildlife Management Institute.
Rights and permissions
About this article
Cite this article
Lapointe, N.W.R., Thorson, J.T. & Angermeier, P.L. Relative roles of natural and anthropogenic drivers of watershed invasibility in riverine ecosystems. Biol Invasions 14, 1931–1945 (2012). https://doi.org/10.1007/s10530-012-0204-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-012-0204-2