
Abstract
The invasive Asian tiger mosquito, Aedes albopictus, has been established on the French overseas island of Mayotte in the Indian Ocean since 2007. Despite the presence of a resident population of Aedes aegypti, Ae. albopictus has expanded its presence in urban areas, in contrast to the pattern of invasion by this mosquito elsewhere in the world. We undertook a comparative study of the distribution and abundance of Ae. albopictus and Ae. aegypti populations between 2007 and 2010 in different types of urban landscapes to document the invasion process. Urban and suburban areas at five localities on Mayotte were selected and ten houses with mosquito-infested larval habitats on the property were located in each area. To calculate variables that might explain mosquito infestation, the areas around the houses selected were overlaid on maps with grid cells (each 25 × 25 m) and the areas inside grid cells were analyzed to define landscape characteristics (percentage of built on land and human density) that might be predictive of Ae. albopictus presence and abundance. The proportion of sites occupied by only Ae. albopictus and the relative abundance of this species relative to total Aedes larvae and pupae significantly increased between 2007 and 2010, in both urban and in rural areas. In the 2010 survey, the number of larval habitats occupied by Ae. aegypti in the sampled area decreased sharply compared to 2007. The proportion of land with paved or hard surfaces (roads, buildings, hereafter “built-up area”), the increase of urbanized areas between 2003 and 2008, and the density of human residents were all correlated with higher relative abundance of Ae. albopictus. The urban areas of Mayotte have significant amounts of vegetation even in the more densely populated areas, and this factor may have facilitated the invasion of Ae. albopictus by providing abundant adult resting sites. These findings are relevant for vector control strategies and forecasting the success of invasions of Ae. albopictus in urban areas in other countries.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Aedes albopictus (Skuse) (Diptera: Culicidae), also called the Asian tiger mosquito, is a container-breeding mosquito that has become invasive in many parts of the world since the 1980s (Benedict et al. 2007). This mosquito is native to Asia and has extended its range into Africa, Europe, the Americas, and some Pacific and Indian Ocean islands via intercontinental shipments of tires (Hawley 1988). This range expansion of Ae. albopictus has been driven by its strong physiological and ecological plasticity, which has allowed it to thrive in a wide range of artificial and natural larval sites, climates and habitats (Paupy et al. 2009).
Aedes aegypti (L.), first described in 1862 from Africa, is a pantropical species, with the widest known geographical range for any single mosquito species (between 45° N and 35° S) (Christophers 1960). Its global expansion occurred before that of Ae. albopictus (Tabachnick 1991). Aedes aegypti is considered a polytypic species, composed of at least two forms, a domestic (urban) form and a relatively more rural one (Tabachnick 1991). Due to frequent invasion of new areas, the current worldwide distributions of Ae. albopictus and Ae. aegypti now overlap.
Both mosquitoes have similar larval niches and are often found in the same larval habitats, but their distributions can be modified by local environmental conditions. For example in Cameroon, where both species occur, Ae. aegypti is dominant in both more arid and more urban areas (Kamgang et al. 2010). Differences in dominance of mosquito vectors along urban–rural gradients have been extensively studied owing to the ecological and epidemiological importance of even small shifts in vector communities (Chan et al. 1971; Braks et al. 2003; Tsuda et al. 2006; Cox et al. 2007; Bagny et al. 2009a). Aedes aegypti is well adapted to the domestic environment, and therefore its abundance in a region is positively correlated with increasing urbanization. In contrast, the distribution of Ae. albopictus is associated with vegetation in both rural and urban areas, and its abundance is usually reduced by urbanization (Hawley 1988; Braks et al. 2003; Maciel-De-Freitas et al. 2006; Tsuda et al. 2006). Despite this association, Ae. albopictus has been found in some very densely populated urban areas such as Rome in the presence of Culex (L.) pipiens (Carrieri et al. 2003).
Following Ae. albopictus invasion, populations of Ae. aegypti have been observed to decline in several locations, including the southeastern United States (Lounibos 2007), Brazil (Benedict et al. 2007), and Reunion Island in the Indian Ocean (Bagny et al. 2009b). One widespread hypothesis to explain the decline of Ae. aegypti following Ae. albopictus invasion is interspecific larval competition, given that both species often share the same larval habitats (Juliano 2009).
Both mosquitoes are of medical concern because they are known to be efficient vectors of arboviruses such as dengue, chikungunya, and yellow fever (Gratz 2004). Since 2000, chikungunya has re-emerged in many countries and become a threat in temperate countries partly due to the spread of Ae. albopictus (Paupy et al. 2009). Mosquito community structure (the relative abundance and spatial distribution of mosquito species in an area) may be a critical factor in assessing human risk from mosquito-borne diseases (Juliano and Lounibos 2005).
In the southwestern Indian Ocean, both Ae. albopictus and Ae. aegypti are present on various islands, with different patterns of distribution among the islands (Delatte et al. 2011). On the island of Mayotte, Ae. albopictus was introduced during the period 2000–2007. The first record of the distribution of Ae. albopictus and Ae. aegypti on these islands (in 2007) showed that Ae. albopictus was well established, principally in urban areas, whereas Ae. aegypti was more common in suburban and rural areas (Bagny et al. 2009a). There was a large variation in the prevalence of Aedes species among the various localities examined: villages showing a predominance of Ae. aegypti and urban centers being dominated by Ae. albopictus (Bagny et al. 2009a). Adaptation of Aedes species to urban habitats implies that these species can find resting sites, hosts for feeding, and oviposition sites in these areas (Vezzani et al. 2001). Aedes albopictus was likely responsible for the chikungunya epidemics observed in Mayotte in 2005–2006, at least for cases in urban and suburban areas (Bagny et al. 2009a).
The pattern of invasion and dominance observed in Mayotte differed from that in other newly invaded countries. Therefore, in this study, we investigated the spatio-temporal distribution and abundance of the invasive Ae. albopictus and the resident Ae. aegypti in different urban and suburban environments to assess the ongoing expansion of Ae. albopictus on Mayotte. We used GIS to follow the Ae. albopictus spread and to identify landscape characteristics and other factors that may have facilitated its spread on this island.
Materials and methods
Study areas and collection sites
Mayotte (12°45′43 S; 45°12′25 E) is a pair of French islands (376 km²) in the Indian Ocean, belonging to the Comoros archipelago, situated in the Mozambique Channel. In 2011, it became an overseas French department, which may stimulate more rapid development of the islands (Taglioni and Dehecq 2009), increasing the urbanized area. Mayotte is comprised of two islands 1 km apart, Grande Terre and Petite Terre. Surveys for this study were conducted in five localities, four on Grande Terre (Mtsapere, Mtsamoudou, Chiconi, Dzoumogne) and one on Petite Terre (Labattoir) (Fig. 1) (Bagny et al. 2009a). The sample sites selected were characterized as to their population density, total land area, and proportion of area covered by buildings or roads (=% built-up land) (Table 1). Communities where sampling was done fell into three size classes: two cities (ca 11–15,000) (Mtsapere and Labbatoir), one town (ca 5,000) (Chiconi) and two villages (ca 1,500–3,300) (Mtsamoudou and Dzoumogne) (Table 1). The same five locations were sampled in 2007 and 2010. At each locality, two different “areas” (in some cases, the sampled area was not fully contiguous, see example in Fig. 1) were selected, less than 1 km apart, based on their level of urbanization, choosing one urban and one suburban area per locality. An urban area was characterized by a higher (30–60%) percentage of built-up land as compared to a suburban area (<30% built-up land) (Bagny et al. 2009a). In each urban and suburban area, ten houses were sampled that first were determined to have larval habitats occupied by some Aedes mosquitos. Both surveys (2007, 2010) were conducted during the rainy season (March, and mid-January to mid-February, respectively). We chose this season because during this period, larval habitats were plentiful and would not be a limiting factor influencing species distribution. Under rainy season conditions (i.e., unlimited larval habitats), mosquito larvae were considered to be a good representation of adult mosquito populations because the abundance and distribution of larvae would reflect the density of adult mosquitoes mediated by their choice of developmental sites. It is further assumed that adults select larval sites that are optimal for the development of their offspring (Ellis 2008).

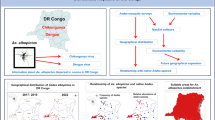
Map of Mayotte indicating the locations of sample sites. Urban areas are shown in gray on maps. Aerial pictures show two of the five sites (Dzoumogne and Mtsapere), and –the surveyed urban and the sub-urban areas are the areas overlaid with black grids
Sampling of immature stages
Two methods were used to count the number of larvae and pupae. In 2007 all larvae and pupae from each mosquito-positive container were collected, counted, and brought back to the regional vector control laboratory (ARS-OI Mayotte) (Bagny et al. 2009a). In 2010, all potential larval habitats were sampled using a standard mosquito dipper (350 ml). Ten dips were randomly taken from each habitat. In small habitats, where this method was not practical, larvae were collected individually using plastic pipettes. All the larvae and pupae were counted and brought back to the regional vector control laboratory for identification (Bagny et al. 2009a).
Landscape analysis of sample areas
To define sample locations and measure the physical variables (% built-up land and human density) characterizing the landscape, we used 1:25,000 topographic maps of Mayotte from National Institute of Geography 2008, complemented with aerial photos at a resolution of 0.5 m pixel geometrically referenced (obtained from the “Préfecture de Mayotte”). We also used an older topographic map of Mayotte (2003) that enabled us to determine the % increase in built-up area between 2003 and 2008 in the surveyed areas (Table 1). This % increase was calculated as following: (constructed surface area in 2003—constructed surface area in 2008)/constructed surface area in 2003 (ArcGis®9.2, ESRI).
We decided to divide the prospected localities into a grid of cells (25 × 25 m), representing, on average, the surface of one or two houses and their yards. The number of cells per grid in an area depended on the size of the area and the number of houses that had to be surveyed to find the 10 required positive houses. Even though the survey was conducted house by house (because permission was required for entry), the data obtained at the house scale were summarized at the cell scale (ArcGis®9.2, ESRI). If there were more than one house per cell, the data on Aedes population abundance at the cell scale corresponded to the sum of the collected data for every house among which more than 50% of the surface was present in the cell. Two types of data are reported here from these surveys. In some analyses we report the proportion of sites that were occupied by each species alone, or together. In other analyses we report the relative abundance of each species in counts of larvae + pupae. For each grid cell, we also computed the total built-up area from the GIS layers showing buildings and roads. This parameter, “% built-up area”, was an index value that ranged from 0 (no hard surfaces, 100% vegetated) to 100 (the entire grid cell covered by buildings or roads), and was divided into four levels: <21, 21–40, 41–60, >60%. In 2010, a sociological survey was conducted along with the entomological survey. Thanks to a questionnaire, we obtained the number of people living in each house. Then we made a calculation of the count of inhabitants per grid cell using the same approach as describe above for Aedes population abundance. This variable (inhabitants per grid cell) had four levels: 1–3, 4–6, 7–9, and 10 or more.
Statistical analyses
Logistic regression was used to evaluate associations between the presence or absence of each species (Ae. albopictus alone (0, 1), Ae. aegypti alone (0, 1)) or the simultaneous presence of both in “positive” sites (grid cells with at least one mosquito larvae or pupae detected in surveys) and several independent variables: (1) survey year, (2) locality, and (3) percentage of built-up area in each grid cell. We also investigated the interactions between sample year and study variables. We performed an analysis of deviance on the most complete model, including additional parameters and interaction effects. The deviance analysis was then used in a step by step approach to identify the significant factors. Significant differences between the proportions of sites with both species, with only Ae. aegypti, or with only Ae. albopictus for the different years, localities and levels of built-up areas in each grid cell were investigated using the binomial test.
To examine species interactions at grid cells where both Aedes species occurred, data on relative species abundance (i.e., numbers of larvae plus pupae of each species in a given grid cell divided by the sum of larvae plus pupae of both species) was analyzed using a generalized linear regression model (based on a quasi-binomial distribution to take into account the over-dispersion in Aedes population abundance data) to evaluate the effects of the study parameters on the relative abundance of Ae. albopictus. The relative abundance of Ae. albopictus was calculated as the number of Ae. albopictus larvae and pupae collected, divided by the total number of Ae. albopictus and Ae. aegypti larvae plus pupae collected in a grid cell. For the data collected in 2010, we also examined the effect of the number of inhabitants per grid cell on the relative abundance of Ae. albopictus with a Kruskal–Wallis rank sum test. Statistical analyses were performed using R software (R Development Core Team 2011).
Results
Out of 274 larval habitats surveyed in 2007, 175 (64%) were positive among which, 145 (53%) contained Ae. albopictus and 156 (57%) contained Ae. aegypti; whereas in 2010, out of 290 larval habitats surveyed, 157 (54%) were positive among which, 138 (47%) contained Ae. albopictus and seventy-seven (26%) contained Ae. aegypti. The proportion of larval habitats occupied by Ae. albopictus did not differ statistically between 2007 and in 2010 (Binomial test, P = 0.1); however, a lower proportion of larval habitats were occupied by Ae. aegypti in 2010 than 2007 (Binomial test, P < 0.001).
For a better understanding of these differences, we performed generalized (logistic) linear regression of the effects of year, locality, and index of the % built-up area on the relative occupancy rates of Ae. aegypti and Ae. albopictus and on the relative abundance of each species (Table 2). The parameters year and locality both significantly affected the relative proportion of sites with both species and with Ae. albopictus only (Table 2). The interaction between year and locality was the only significant parameter for the proportion of sites with only Ae. aegypti. The proportion of sites occupied by both species decreased significantly between 2007 and 2010 in three of the five sampled localities (Fig. 2, Binomial test P < 0.05) (all except Mtsamoudou and Mtsapere). However, Mtsapere had the lowest proportion of occupied habitat containers among all sample sites in both years. In addition, the proportion of sites with only Ae. albopictus increased significantly between 2007 and 2010 in all sample localities (Fig. 2, Binomial test <0.05) except one (Mtsapere). In Mtsamoudou, Dzoumogne, and Labbatoir, there were no sites with only Ae. albopictus in 2007, but by 2010, on average half of the sampled grid cell locations contained only this species. The proportion of sites with Ae. aegypti only decreased significantly between the two surveys at two sites (Mtsamoudou and Dzoumogne) (Binomial test, P < 0.05). At two other sites (Chiconi and Labbatoir), the proportion of sites occupied by Ae. aegypti increased because a few such sites were found in 2010, whereas none had been located in 2007. In Mtsapere, the proportion did not change because there were no sites with only A. aegypti in either 2007 or in 2010 (Fig. 2).

Change between 2007and 2010 in the relative proportion of sites with both species, with Ae. aegypti only and with Ae. albopictus only in each survey locality. *** Refers to significant differences between the proportion of sites between these 2 years in each locality. (Binomial test)
At sites where larvae of both mosquitos were present, both locality and the index of % built-up area had significant effects on the relative abundance of Ae. albopictus (Table 2). In Mtsamoudou and Dzoumogne, the smallest localities of this survey (Table 1), the relative abundance of Ae. albopictus larvae and pupae increased significantly between sample years (Fig. 3a). In 2007, Ae. aegypti was the dominant species in these localities (relative abundance of Ae. albopictus <0.5, Fig. 3a). By 2010, the relative density Ae. albopictus in Dzoumogne and Mtsamoudou had increased to the point that it was similar to levels at the other larger localities dominated by Ae. albopictus. These two localities (Dzoumogne and Mtsamoudou) also showed the largest increase in their index score of the percentage of land covered by buildings and roads (from 2003 to 2008) (Table 1). In the other three localities (Chiconi, Labattoir, and Mtsapere), the relative abundance of Ae. albopictus did not change between the 2 years, being tilted in favor of Ae. albopictus in both years (Fig. 3a).

The relative abundance of Ae. albopictus immature stages in co-occurrence sampled areas (a) for the different survey localities, arranged from smallest (left) to largest (right) communities; (b) and the relative abundance of Ae. albopictus in relation to the percentage of built-up area in the set of grid cells that defined the sampled portion of a community
When the effect of the % of built-up area was considered, a trend in favor of Ae. albopictus emerged. At Mtsapere, which had the highest proportion of area covered with buildings and roads and the lowest vegetation coverage (Table 1), sites occupied by both mosquito species were highly dominated by Ae. albopictus (Fig. 3a). In general, Ae. albopictus was the most abundant species in the areas with a high level of built-up land (>40%) (Fig. 3b), particularly in 2010. In the less built-up areas (<40%), the relative abundances of Ae. aegypti and Ae. albopictus were almost the same (ca 50% each) (Fig. 3b) in both 2007 and 2010.
Similarly, an increase in the number of people living in a sample grid cell favored dominance by Ae. albopictus (Fig. 4). In 2010, sites (sample grid cells) with Ae. albopictus only had an average of seven inhabitants, whereas grid cells with Ae. aegypti only had an average of just three inhabitants. At sites with both mosquito species, the relative species abundance was significantly influenced by the numbers of inhabitants per grid cell (Kruskal–Wallis rank sum test, P = 0.002) (Fig. 4). At lower human densities (1–3, 4–6, or 7–9 per grid cell), the relative abundance index was equal between mosquito species (ca 0.5), but in the highest human density class (10 or more per grid cell), there was a clear dominance of Ae. albopictus (Fig. 4).

Correlation of the relative abundance of Ae. albopictus immature stages and the number of inhabitants in 2010 summed over grid cells
Discussion
On Mayotte, the relative dominance of Ae. albopictus immature stages in sites (grid cells) increased between 2007 and 2010. While the proportion of sites occupied by Ae. albopictus did not increase between sample years, the proportion of sites occupied by Ae. aegypti decreased significantly. As a result, there was a decrease in both the number of sites where both species co-occur and an increase (by default) in the number of sites with Ae. albopictus alone. The relative dominance of Ae. albopictus over Ae. aegypti in 2010 was observed in all localities, even in the smallest towns, where Ae. aegypti had been the dominant species in 2007. Our results also suggest a decline in the Ae. aegypti population on the island in all localities, making Mayotte another place where the arrival of Ae. albopictus has caused the decline in the population of Ae. aegypti.
The increase in dominance of Ae. albopictus immature stages from 2007 to 2010 was affected both by the locality and the percentage of area in a grid cell covered by roads or buildings. Chiconi and Labattoir shared a similar pattern of mosquito occupancy and dominance, with little change between sample years, but there is no obvious common factor between these sites that might explain this pattern of mosquito presence. Labbatoir is an important urban center, but its location on Petite Terre could explain the differences observed relative to Mtsapere, the other urban center. Chiconi is situated on the west coast of the main island, and although it is quite big, it is surrounded by natural environment. This locality is isolated from the main urban centers of Mayotte, located on the east coast. The mountainous area between Chiconi and the east coast could constitute a natural barrier for mosquito movement, limiting expansion into this area. The two smallest localities in our study, Mtsamoudou and Dzoumogne, showed the greatest degree of change in both mosquito prevalence and relative abundance between 2007 and 2010, with the Ae. albopictus population becoming more dominant. Mayotte is urbanizing quickly, especially in suburban areas such as these two villages. In Mtsapere, the most urbanized locality in our study, the increase in Ae. albopictus population was less significant, as this species was already the dominant Aedes mosquito in 2007 (Bagny et al. 2009a). Mtsapere is near the main city, Mamoudzou, where urbanization first began and is still on-going. This area is also the main port of entry for the island for both visitors and goods.
Aedes albopictus was the dominant mosquito in the most densely populated areas (number of inhabitants per grid cell >10) and dominance of this mosquito was linked with the degree of building coverage within a sample grid cell. Other studies have shown Ae. albopictus to be the dominant mosquito in urban areas, especially when it was in the presence of Ochlerotatus triseriatus (Say) and Culex pipiens (Swanson et al. 2000; Barker et al. 2003; Carrieri et al. 2003), but it was dominant less often in the presence of Ae. aegypti (Gilotra et al. 1967). When Ae. albopictus competes with Ae. aegypti, there is generally a segregation in habitat preference, with Ae. aegypti breeding closer to human dwellings (Braks et al. 2003; Carbajo et al. 2006; Tsuda et al. 2006; Higa et al. 2010).
Several factors may help explain the difference in the outcome of Ae. albopictus-Ae. aegypti spatial distribution as observed in Mayotte compared to other regions as discussed above. The strain of Ae. albopictus found in Mayotte is likely the same strain that has invaded many areas around the world in the last 40 years (Delatte et al. 2011). Also, the invasion pattern on Mayotte is probably not the result of Ae. albopictus’ characteristics but is more likely linked to how the mosquito interacts with features of the invaded environment (Lockwood et al. 2008). Characteristics of the Mayotte urban landscape likely facilitated the establishment of Ae. albopictus (Bagny et al. 2009a). Human density in the relatively small urban areas of Mayotte is low (1,000–1,200 inhab/km²), which is far below the population density of Rio de Janeiro, Buenos Aires (15,000 inhab/km², Carbajo et al. 2006), or Florida, both areas where Ae. albopictus is seen typically in more rural areas. In these areas, the concentration of buildings is also very high in comparison with Mayotte. Similarly, the population density on Reunion Island, where Ae. albopictus also occurs in urban areas, is very low (900 inhab/km²). On both Reunion and Mayotte the undeveloped portions of the urban areas are mostly vegetated, and the small urban centers on both islands are separated from the rural areas by only a few kilometers. These features mean that urban areas on Mayotte meet the key requirements for Ae. albopictus: proximity to human hosts, anthropic larval habitats (containers), and resting areas for adult mosquitos (i.e., vegetated areas).
An alternative hypothesis to explain the successful establishment of Ae. albopictus in urban areas of Mayotte could be that Ae. aegypti populations in these areas did not provide a strong resistance to invasion. In Mayotte, the ecological niche of the local Ae. aegypti population is unlikely to be in highly urbanized areas, as is the case in many other countries, because the strain of Ae. aegypti on Mayotte comes from the rural subspecies of Ae. aegypti (Bagny et al. 2009a). That being the case, one can infer that in the absence of any urban species of mosquitos, Ae. aegypti was able to colonize these urban habitats despite being less well adapted to that environment.
Apart from competition between adults of these two mosquitos for oviposition sites, it is also possible that the decrease in the level of co-occurrence of larvae of the two species at sample sites in 2010 may have resulted from competition between larvae for resources, which may favor Ae. albopictus (Juliano 2009).
Still another hypothesis, formulated recently by Tripet et al. (2011), is the existence of asymmetric mating competition (with males of Ae. albopictus mating and sterilizing females of Ae. aegypit more often than the reverse), which may have contributed to the reduction of Ae. aegypti by invasive Ae. albopictus populations in the southern United States. While this type of interaction may be a factor in the events observed in Mayotte, further study would be required to determine if it really played a role or not.
The invasion of Mayotte by Ae. albopictus and its colonization of urban areas has changed the arbovirus vector risk on the island. The presence of Ae. albopictus in the most densely populated areas of Mayotte is of special interest for public health as this mosquito, which is potentially the main vector of some arboviruses, can facilitate disease transmission during epidemic events. Rural areas of Mayotte, which have urbanized faster than other parts of the island, are likely to experience Ae. albopictus establishment and expansion. This outcome, which has not been seen in other, bigger, urban areas, seems to have arisen because the density of urban areas on Mayotte is lower, leaving enough vegetation in developed areas to meet the ecological requirements of this invasive mosquito.
References
Bagny L, Delatte H, Elissa N, Quilici S, Fontenille D (2009a) Aedes (Diptera: Culicidae) vectors of arboviruses in Mayotte (Indian Ocean): distribution area and larval habitats. J Med Entomol 46:198–207
Bagny L, Delatte H, Quilici S, Fontenille D (2009b) Progressive decrease in Aedes aegypti distribution in Reunion Island since the 1900s. J Med Entomol 46:1541–1545
Barker CM, Paulson SL, Cantrell S, Davis BS (2003) Habitat preferences and phenology of Ochlerotatus triseriatus and Aedes albopictus (Diptera : Culicidae) in southwestern Virginia. J Med Entomol 40:403–410
Benedict MQ, Levine RS, Hawley WA, Lounibos LP (2007) Spread of the tiger: global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis 7:76–85
Braks MAH, Honorio NA, Lourenco-De-Oliveira R, Juliano SA, Lounibos LP (2003) Convergent habitat segregation of Aedes aegypti and Aedes albopictus (Diptera : Culicidae) in southeastern Brazil and Florida. J Med Entomol 40:785–794
Carbajo AE, Curto SI, Schweigmann NJ (2006) Spatial distribution pattern of oviposition in the mosquito Aedes aegypti in relation to urbanization in Buenos Aires: southern fringe bionomics of an introduced vector. Med Vet Entomol 20:209–218
Carrieri M, Bacchi M, Bellini R, Maini S (2003) On the competition occuring between Aedes albopictus and Culex pipiens (Diptera: Culicidae) in Italy. Environ Entomol 32:1313–1321
Chan YC, Chan KL, Ho BC (1971) Aedes aegypti (L.) and Aedes albopictus (Skuse) in Singapore-City. 1. Distribution and density. Bull World Health Org 44:617–627
Christophers SR (1960) Aedes aegypti (L.) the yellow fever mosquito: its life history, bionomics and structure. Cambridge University Press, Cambridge
Cox J, Grillet ME, Ramos OM, Amador M, Barrera R (2007) Habitat segregation of dengue vectors along an urban environmental gradient. Am J Trop Med Hyg 76:820–826
Delatte H, Bagny Beilhe L, Brengue C, Bouetard A, Paupy C, Fontenille D (2011) The invaders: phylogeography of dengue and chikungunya viruses vectors, Aedes albopictus, Aedes aegypti and Aedes mascarensis, on the South West islands of the Indian Ocean. Infect Genet Evol 11(7):1769–1781
Ellis AM (2008) Linking movement and oviposition behaviour to spatial population distribution in the tree hole mosquito Ochlerotatus triseriatus. J Anim Ecol 77:156–166
Gilotra SK, Rozeboom LE, Bhattacharya NC (1967) Observations on possible competitive displacement between populations of Aedes aegypti L. and Aedes albopictus Skuse in Calcutta. Bull WHO 37:437–446
Gratz NG (2004) Critical review of the vector status of Aedes albopictus. Med Vet Entomol 18:215–227
Hawley AH (1988) The biology of Aedes albopictus. J Am Mosq Contr Assoc 4:1–40
Higa Y, Thi Yen N, Kawada H, Hai Son T, Thuy Hoa N, Takagi M (2010) Geographic distribution of Aedes aegypti and Aedes albopictus collected from used tires in Vietnam. Am Mosq Contr Assoc 26:1–9
Juliano SA (2009) Species interactions among larval mosquitoes: context dependence across habitat gradients. Annu Rev Entomol 54:37–56
Juliano SA, Lounibos LP (2005) Ecology of invasive mosquitoes: effects on resident species and on human health. Ecol Lett 8:558–574
Kamgang B, Happi JY, Boisier P, Njiokou F, Herve JP, Simard F, Paupy C (2010) Geographic and ecological distribution of the dengue and chikungunya virus vectors Aedes aegypti and Aedes albopictus in three major Cameroonian towns. Med Vet Entomol 24:132–141
Lockwood JL, Hoopes M, Marchetti M (2008) Invasion ecology. Wiley Blackwell, New York
Lounibos LP (2007) Competitive displacement and reduction. J Am Mosq Contr Assoc 23:276–282
Maciel-De-Freitas R, Brocki R, Goncalves JM, Codeco CT, Lourenco-De-Oliveira R (2006) Movement of dengue vectors between the human modified environment and an urban forest in Rio de Janeiro. J Med Entomol 43:1112–1120
Paupy C, Delatte H, Bagny L, Corbel V, Fontenille D (2009) Aedes albopictus, an arbovirus vector: from the darkness to the light. Microb Infect 11:1177–1185
Swanson J, Lancaster M, Anderson J, Crandell M, Haramis L, Grimstad P, Kitron U (2000) Overwintering and establishment of Aedes albopictus (Diptera : Culicidae) in an urban La Crosse virus enzootic site in Illinois. J Med Entomol 37:454–460
Tabachnick WJ (1991) Evolutionary genetics and arthropod-borne disease. The yellow fever mosquito. Am Entomol 37:14–24
Taglioni F, Dehecq J-S (2009) L’environnement socio-spatial comme facteur d’émergence des maladies infectieuses- Le chikungunya dans l’océan Indien. EchoGéo 9 (Open Access). URL: http://echogeo.revues.org/11168
R Development Core Team (2011) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. URL http://www.R-project.org/
Tripet F, Lounibos LP, Robbins D, Moran J, Nishimura N, Blosser EM (2011) Competitive reduction by satyrization? Evidence for interspecific mating in nature and asymmetric reproductive competition between invasive mosquito vectors. J Am Trop Med Hyg 85:265–270
Tsuda Y, Suwonkerd W, Chawprom S, Prajakwong S, Takagi M (2006) Different spatial distribution of Aedes aegypti and Aedes albopictus along an urban-rural gradient and the relating environmental factors examined in three villages in northern Thailand. J Am Mosq Contr Assoc 22:222–228
Vezzani D, Velazquez S, Soto S, Schweigmann N (2001) Environmental characteristics of the cemeteries of Buenos Aires City (Argentina) and infestation levels of Aedes aegypti (Diptera: Culicidae). Mem Inst Oswaldo Cruz 96:467–471
Acknowledgments
The authors thank J. Thiriat, B. Zumbo and the technicians of the regional vector control laboratory of Mayotte (ARS-OI) for invaluable help with the field and the laboratory work and Mr Cieslak from “Préfecture de Mayotte” for the GIS data. We also thank anonymous reviewers and D. Simberloff for useful comments on the manuscript. We are grateful to VanDriesche scientific editing for the English improvement. This work was founded in part by CIRAD, IRD, the “Conseil Régional de la Réunion”, the “EU”, and “Agence Régional de Santé Océan Indien” (ARS OI).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bagny Beilhe, L., Arnoux, S., Delatte, H. et al. Spread of invasive Aedes albopictus and decline of resident Aedes aegypti in urban areas of Mayotte 2007–2010. Biol Invasions 14, 1623–1633 (2012). https://doi.org/10.1007/s10530-012-0177-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-012-0177-1