
Abstract
Release from natural enemies is frequently cited as an important factor contributing to plant invasions. But such effects are likely to be temporary—native herbivores can form new plant-herbivore associations and co-evolved insects might reach the new range. While the potential effects of the initial enemy release have been well studied, the consequences of any resumption of herbivory are poorly understood. Alternanthera philoxeroides is one of the most widespread invasive plants in China and is attacked both by a specialist herbivore introduced from the native range, Agasicles hygrophila, and a native beetle Cassida piperata Hope which has formed a new association. However, these insects are not found throughout the invaded range. To test the effect of the history of population exposure to herbivory on compensatory ability, plants were cultured from 14 populations around China that differed in whether A. hygrophila or C. piperata were present. Treatment plants were exposed to herbivory by A. hygrophila for a week until 50% of the leaf area was defoliated, then grown for 80 days. Plants from populations with prior exposure to herbivory (of any kind) accumulated more root mass than populations without prior exposure, indicating that prior exposure to insects can stimulate plant compensation to herbivory. We would recommend that potential changes in plant tolerance in response to prior exposure to herbivory are considered in invasive plant management plans that employ bio-control agents.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Most invasive plant species have been introduced to their new ranges without the full suite of natural enemies with which they co-evolved (e.g. natural enemy release) (Keane and Crawley 2002). In the absence of specialist herbivores, invasive species may reallocate resources from defence to growth, reproduction or both, as proposed by the Evolution of Increased Competitive Ability Hypothesis (EICA) (Blossey and Nötzold 1995). However, the enemy release, and the associated fitness gain, might be temporary as native herbivores tend to accumulate on non-native species over time (Siemann et al. 2006). Moreover, co-evolved natural enemies, such as herbivorous insects, might also eventually arrive either accidentally or deliberately (e.g. through classical biological control releases), although the character and intensity of the re-association between plant and herbivore can differ from the interaction in the native range (Garcia-Rossi et al. 2003).
In response to the re- or new-associated interactions with herbivores, invasive plants may undergo rapid evolutionary changes in defence, e.g. resistance and/or tolerance (Thompson 1998). For instance, in populations of invasive wild parsnips, Pastinaca sativa, there was an increase in toxic furanocoumarins content after re-association with Parsnip webworm, Depressaria pastinacella (Zangerl and Berenbaum 2005), leading to an increase in herbivore resistance. What has received less attention is the potential for invasive plants to evolve greater tolerance of herbivory after re- or new-association with herbivores.
Tolerance is the ability of a plant to re-grow and reproduce after herbivory (Strauss and Agrawal 1999). Plant compensatory ability, defined here as the difference in fitness between herbivory damaged and undamaged individuals of the same genotype, is known as an important measure of plant tolerance (Belsky 1986). Herbivory-induced compensatory growth for a single plant genotype can range from under-compensation if herbivory damage is not fully replaced, to equal-compensation if plants fully recover from herbivory, and overcompensation if plants have greater fitness when damaged (Maschinski and Whitham 1989). Proposed mechanisms for plant compensation include increases in photosynthetic ability, utilization of stored reserves, and changes in phenology and resource reallocation (Strauss and Agrawal 1999; Tiffin 2000). Empirical evidence indicates that herbivory may evolutionarily increase a plant’s compensatory ability, especially when the risk of damage is predictable and high (Juenger and Bergelson 2000; Lennartsson et al. 1997). Therefore, the re- or new-associated herbivores would be expected to create a selection pressure on the compensatory ability of invasive plants, particular if herbivores occur at high densities.
The evolutionary responses of invasive plants to herbivore have been well studied (Müller-Schärer et al. 2004), providing insights into many aspects of plant evolutionary biology, especially evolution of plant defence (Müller-Schärer et al. 2004; Thompson 1998). However, almost all these studies focus on sexual plants, while little information is available on the evolutionary responses of clonally propagated invasive plants to herbivores. In fact, some of the globally important invasive plants reproduce mainly clonally, e.g. Alternanthera philoxeroides (Julien et al. 1992) and Eichhornia crassipes (Li et al. 2006), in their non-native ranges. There is increasing evidence indicating that somatic mutation and genomic modification in clonal plants are frequent and provide genetic and epigenetic variation that contributes to their adaptive evolution, especially for clonally propagated domesticated plants (Prentis et al. 2008; McKey et al. 2010). Moreover, intra-clonal variation in plant defence and selective impacts of herbivory on plant defence has been reported in clonal plants (Monro and Poore 2004; Whitham and Slobodchikoff 1981). Therefore, herbivory can impose selection on defence of clonal invasive plants; indeed such effects are likely to be promoted by the invasion process (Prentis et al. 2008).
Here, we report the results of an experiment designed to examine the consequences of prior exposure to insect herbivores on the compensatory ability of the clonal invasive plant alligator weed, Alternanthera philoxeroides (Mart.) Griseb (Amaranthaceae). Specifically, we hypothesized that plant populations from sites with herbivores would show a greater capacity for compensation than populations from sites without them. We also tested whether the source of herbivory encountered (native insects, introduced insects, or both) influenced the form of plant compensation.
Materials and methods
Study species
Alternanthera philoxeroides is a perennial herbaceous plant with horizontal to vertical stems that can grow in aquatic and terrestrial habitats. Each stem consists of nodes that are capable of producing individual plantlets. Roots develop at closely spaced nodes along stems (Julien et al. 1995). The species is native to South America and has become a widespread invader in the Asia–Pacific region and the United States (Julien et al. 1995). It was first introduced into China (Shanghai) as a forage plant in the 1930s, and since then it has been distributed to as many as 20 provinces in Central and Southern China (Ma 2001), with its range still expanding in Northern China (Geng et al. 2007). According to Julien et al. (1992) A. philoxeroides produces seeds in its native South American range, but rarely sets seeds and propagates solely by vegetative means from stem and root buds in Australia, the United States and China.
Agasicles hygrophila, a host-specific leaf grazer of A. philoxeroides, has been widely distributed for biological control (Buckingham 2002). The beetle has curbed A. philoxeroides invasion in many infested rivers and ponds (Ma 2001; Sainty et al. 1998), but has failed to control the plant in semi-aquatic and terrestrial habitats despite occurring on A. philoxeroides in these habitats (Ma 2001). The beetle was first introduced to China in 1986, but has not spread to all areas affected by A. philoxeroides (Ma 2001), and so plant populations have different histories of re-association. In China A. philoxeroides is also defoliated by a native tortoise beetle, Cassida piperata Hope (Coleoptera: Cassididae) (Lin et al. 1990) in terrestrial habitats and occasionally in aquatic habitats (Lu and Ding, unpublished). Both A. hygrophila and C. piperata are aboveground herbivores. Adults and larvae of A. hygrophila feed on leaves and stems of A. philoxeroides, often producing feeding holes and “trenches” on leaves; adults and larvae of C. piperata only feed on leaves of A. philoxeroides, often producing tiny feeding holes. Previous studies have found that A. philoxeroides can compensate rapidly in terms of biomass production after damage by herbivory (natural and simulated) and mowing in terrestrial habitats, which may be related to its high root mass (Lu et al. 2010; Lu and Ding 2010; Sun et al. 2009; Wilson et al. 2007). Moreover, in common garden experiments we found that A. philoxeroides compensates for defoliation caused by A. hygrophila and C. piperata in a similar manner (Lu and Ding, unpublished).
In this study, we focused on herbivory by A. hygrophila. The A. hygrophila adult beetles used in our experiments were collected from A. philoxeroides in the suburbs of Wuhan, Hubei Province.
Plant sampling locations
In early spring 2007, we collected plants from 14 terrestrial populations (>10 km apart from each other) of A. philoxeroides in five Chinese provinces (Table 1); populations from the same province were assumed to be independent for purposes of this study. Based on biological control records (Ma 2001; Yang 2001), feeding records for C. piperata (Lin et al. 1990), and our 2006–2007 field surveys on insect feeding, these populations were categorized into four groups based on the presence (or absence) of A. hygrophila and C. piperata (Table 1). At each site we collected 10 plants from 4 locations (40 plants/population) >20 m apart in open fields near aquatic habitats (riverbank). We collected a stem (10–15 cm long) from each plant and immediately placed the stems in moist foam in coolers with dry ice for transport to the laboratory. In the laboratory we cut stems to 4–5 cm lengths; each cut stem contained a single node. Stem pieces were then planted vertically in plastic containers (50 × 40 × 30 cm deep) filled with a homogenized mixture of peat, topsoil and sand in a greenhouse under natural light at Wuhan Botanical Garden (Chinese Academy of Sciences). The containers were caged to exclude herbivores. To reduce the possible carryover impact of parental environment on plant compensation, we removed 4–5 cm stem pieces (each bearing one node) from the new shoots for our experiments after 10 weeks.
Herbivory treatments
The experiment was conducted in a greenhouse at Wuhan Botanical Institute/Wuhan Botanical Garden (Chinese Academy of Sciences) from April to early December, 2007. Mean daily air temperature in the greenhouse was 20–35°C from late April to September and 15–25°C from October to early December.
Three randomly selected stem pieces from the same population (stems from the same population were mixed together before planting) were planted vertically in a pot (16 cm in diameter, 14 cm in height) filled with a homogenized mixture of peat, topsoil and sand. Twenty days before herbivory treatment, we retained similar-sized plants and thinned the plants to one plant per pot to minimise plant size variation among pots. Depending on the availability of stem pieces, there were 34–46 plants per population.
Half the potted plants for each population received insects (herbivory treatment: 50% of the leaf area of each plant was removed by herbivores), and half served as controls (undamaged control) (giving 17–23 replicates for each treatment). Two days before herbivory treatment, half the shoots for each plant were caged. In the herbivory treatment, 6–8 A. hygrophila adults were released into each cage. After 7 days, all the caged shoots in the herbivory treatment were completely defoliated and the insects and cages were removed. Plants were then grown for an additional 80 days. During this time, plants were watered every 2 days and pot positions in the greenhouse were randomised every month. At harvest the numbers of stem and root buds were counted, and above and below ground biomass were separated and dried (80°C for 48 h) before weighing.
Data analysis
Prior to analysis, total, root and shoot mass, and root to shoot biomass ratio (R/S) were log10-transformed to achieve normality and homoscedasticity.
We carried out analyses with the full data set, including data for all harvested plants, to test the potential impacts of herbivory, insect exposure history (previously exposed or un-exposed to herbivory), population and their interactions on plant performance. We also analysed the data for plants with insect exposure history only (previously exposed to A. hygrophila, C. piperata, or both), to compare the impacts of previous exposure to different insect species on plant response to subsequent herbivory.
All measured plant traits were analysed with three-way nested ANOVAs. With the full data set, the ANOVA model included insect exposure history (previously exposed or un-exposed to herbivory) and herbivory treatment as fixed factors, and plant population as a random factor nested within insect exposure history. For the data for plants with insect exposure history alone, the ANOVA model included insect species (A. hygrophila, C. piperata, or both) and herbivory treatment as fixed factors, and plant population as a random factor nested within insect species. The effects of insect exposure history and insect species were tested over the nested population term. Differences between individual means were tested with t tests.
Data analyses were carried out using Statistical analysis system (SAS Version 8.1, SAS Institute).
Results
Effect of herbivory and insect exposure history on plant biomass and vegetative bud formation
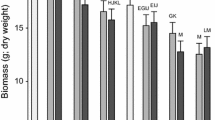
The interaction between herbivory and insect exposure history affected plant root mass, but did not affect plant shoot mass, R/S, or total biomass (Table 2, Fig. 1). In support of our prediction, herbivory significantly increased root mass (P = 0.0007, Fig. 1c) of plants with insect exposure history when compared to undamaged control plants, while herbivory did not affect root mass of plants without insect exposure history when compared to undamaged control plants (P > 0.05, Fig. 1c).

Effect of herbivory in the greenhouse experiment on a total, b shoot and c root mass of plants exposed or unexposed to herbivores in the field: means across populations. Values are means ±1 SE (back transformed for the total, shoot and root mass). Symbols above pairs of bars show probability (t test) that the two means did not differ: (no symbol) >0.05; *0.01–0.05; **0.001–0.01; ***<0.001
The interaction between herbivory and population had a significant effect on plant total and shoot mass, and marginally affected plant root mass (Table 2). Of the 14 plant populations, five overcompensated, eight equal-compensated and one under-compensated in terms of plant total biomass in response to herbivory (Fig. 2a). In support of our prediction, four of the five populations that overcompensated for herbivory (HG, XY, ZWY and LHL) were from locations with herbivores that removed 10–40% plant leaf area historically, while only one (YMC) was from a location with no herbivores and had no known history of herbivore interaction (Fig. 1a, Table 1), and the one (DL) that under-compensated for herbivory was from a location with no known insect exposure history (Fig. 2a). In three of the overcompensating populations (HG, ZWY and YMC), the shoot mass of damaged plants was significantly higher compared with that of the undamaged plants (Fig. 2b).

Impacts of herbivory in the greenhouse experiment on a total and b shoot mass, and c number of root buds of plants exposed or unexposed to herbivores in the field: means for individual populations. Values are population means ±1 SE (back transformed for the total and shoot mass). Symbols above pairs of bars show probability (t test) that the two means did not differ: (no symbol) >0.05; *0.01–0.05; **0.001–0.01; ***<0.001
The total number of vegetative buds was only affected by herbivory and plant population (Table 2), which did not support our predictions. The total number of vegetative buds was greater (14.57%, F 1, 426 = 1.787, P = 0.008) in damaged than in control plants.
The number of root buds was significantly affected by herbivory, plant population and their interaction, whereas the number of root buds was not affected by other factors or their interactions (Table 2), which also failed to support our prediction. The number of root buds of plants from HG, CM, HQ and ZWY populations increased under herbivory, while herbivory did not influence the number of root buds of plants from other populations (Fig. 2c). The number of stem buds was affected by plant population and was not affected by other factors or their interactions (Table 2).
Differential effects of previous exposure to different insect species
For the plants with insect exposure history, insect species (previously damaged by native, introduced, or both insects) did not influence plant total biomass, total number of vegetative buds, or numbers of root and stem buds (Table 3). None of the traits were affected by the interaction between herbivory and insect species, while plant total biomass, shoot mass and R/S were affected by the interaction between herbivory and plant population (Table 3).
Discussion
Compensatory growth can potentially facilitate a plant invasion (Schierenbeck et al. 1994; Wilsey and Polley 2006) and could be a factor that hampers the biological control of exotic plants (Garcia-Rossi et al. 2003). However, this has received little attention (Bossdorf et al. 2005; Müller-Schärer et al. 2004). We examined here the compensatory ability of A. philoxeroides populations across its large new range in China. As hypothesized, we found differences in compensatory ability between plants with and without insect exposure history. A. philoxeroides populations from locations with insect exposure history accumulated more root mass and showed greater compensatory ability in response to herbivory than populations from locations without insect exposure history. While there was a large inter-population variance, the type of herbivory (A. hygrophila, C. piperata, or both) was not found to affect the level of compensation.
Differences in compensatory ability between plant populations with different grazing histories have also been reported in Artemisia ludoviciana (Damhoureyeh and Hartnett 2002), Sorghastrum nutans (Damhoureyeh and Hartnett 2002), Agropyron smithii Rydb (Detling and Painter 1983; Polley and Detling 1988) and Bouteloua curtipendula var. caespitosa (Smith 1998). However, the differing intensities of compensatory response by populations of A. philoxeroides with varying herbivory exposure history, and the clonal nature of the plant, suggests that the effect seen here might be due to epigenetic inheritance (McKey et al. 2010; Prentis et al. 2008). Indeed, Gao et al. (2010) reported that genome-wide DNA methylation alternations enable A. philoxeroides to adapt to varying water availability rapidly.
In response to long-term herbivory, a plant may display evolutionary or plastic changes in physiology or morphology that may increase compensatory ability, e.g. forming more horizontally-oriented leaves, allocating fewer resources to new leaves or more resources to roots, or storing more reserves belowground (Carman and Briske 1985; Mack and Thompson 1982; McIntire and Hik 2002). For example, ramets of the clonal aquatic plant Ranunculus lingua that had experienced substantial herbivory produced larger but fewer rhizomes and experienced less reduction in most growth parameters when exposed to extensive herbivory, in comparison with ramets that had experienced less herbivory (Johansson 1994). The greater compensatory ability of A. philoxeroides with insect exposure history compared to plants without insect exposure history may have resulted primarily from increase in root storage mass, which has been reported to be highly correlated with its compensatory ability (Wilson et al. 2007; Lu and Ding 2010).
Our finding that A. philoxeroides from five populations (four with and one without herbivory exposure history) over-compensated herbivory provides insights into the plant’s physiological potential for compensation. However, whether the plant can overcompensate in response to herbivory under natural field conditions needs further clarification, since a plant’s compensation intensity is also influenced by other environmental conditions, including resource availability, time, frequency and intensity of damage, neighbouring plants, and duration of the recovery period (Maschinski and Whitham 1989; Strauss and Agrawal 1999). Schooler et al. (2006, 2007) found that herbivory significantly suppressed the growth of A. philoxeroides when the recovery period was just 5 weeks or less.
Our findings may hold broad implications for the biological control of invasive plants. Introducing and releasing natural enemies in some cases may favour, rather than suppress, the growth of target species through increased compensatory response to herbivory—an undesirable outcome. The ability of native herbivores to reduce plant performance and increase the mortality of invasive plants (Maron and Vilà 2001), has prompted the suggestion to use native herbivores as biological control agents (Cronin et al. 1999; Mitchell et al. 2006). However, the compensatory growth of A. philoxeroides in response to the native beetle in our study show that even the use of native species may not always result in control. Thus, to improve biocontrol efficacy and avoid potential negative effect of biocontrol agents on native plants, an explicit understanding of the target plant’s defence strategy is crucial before approval and introduction of foreign natural enemies, as well as the use of native herbivores.
References
Belsky AJ (1986) Does herbivory benefit plants? A review of the evidence. Am Nat 127:870–892
Blossey B, Nötzold R (1995) Evolution of increased competitive ability in invasive nonindigenous plants: a hypothesis. J Ecol 83:887–889
Bossdorf O, Auge H, Lafuma L, Rogers WE, Siemann E, Prati D (2005) Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia 144:1–11. doi:10.1007/s00442-005-0070-z
Buckingham GR (2002) Alligatorweed. In: van Driesche R, Blossey B, Hoddle M, Lyon S, Reardon R (eds) Biological control of invasive plants in the Eastern United States, pp 5–15, USDA Forest Service Publication FHTET-2002-04
Carman JG, Briske DD (1985) Morphologic and allozymic variation between long-term grazed and non-grazed populations of the bunchgrass Schizachyrium scoparium var. frequens. Oecologia 66:332–337. doi:10.1007/BF00378294
Cronin G, Schlacher T, Lodge DM, Siska EL (1999) Intraspecific variation in feeding preference and performance of Galerucella nymphaeae (Chrysomelidae: Coleoptera) on aquatic macrophytes. J North Am Benthol Soc 18:391–405
Damhoureyeh SA, Hartnett DC (2002) Variation in grazing tolerance among three tallgrass prairie plant species. Am J Bot 89:1634–1643
Detling JK, Painter EL (1983) Defoliation responses of western wheatgrass populations with diverse histories of prairie dog grazing. Oecologia 57:65–71. doi:10.1007/BF00379563
Gao LX, Geng YP, Li B, Chen JK, Yang J (2010) Genome-wide DNA methylation alterations of Alternanthera philoxeroides in natural and manipulated habitats: implications for epigenetic regulation of rapid responses to environmental fluctuation and phenotypic variation. Plant Cell Environ 33:1820–1827. doi:10.1111/j.1365-3040.2010.02186.x
Garcia-Rossi D, Rank N, Strong DR (2003) Potential for self-defeating biological control? Variation in herbivore vulnerability among invasive Spartina genotypes. Ecol Appl 13:1640–1649. doi:10.1890/01-5301
Geng YP, Pan XY, Xu CY, Zhang WJ, Li B, Chen JK, Lu BR, Song ZP (2007) Phenotypic plasticity rather than locally adapted ecotypes allows the invasive alligator weed to colonize a wide range of habitats. Biol Invasions 9:245–256. doi:10.1007/s10530-006-9029-1
Johansson ME (1994) Life history differences between central and marginal populations of the clonal aquatic plant Ranunculus lingua: a reciprocal transplant experiment. Oikos 70:65–72
Juenger T, Bergelson J (2000) The evolution of compensation to herbivory in scarlet gilia, Ipomopsis aggregata: herbivore-imposed natural selection and the quantitative genetics of tolerance. Evolution 54:764–777. doi:10.1111/j.0014-3820.2000.tb00078.x
Julien MH, Bourne AS, Low VHK (1992) Growth of the weed Alternanthera philoxeriodes (Martius) Grisebach, (alligator weed) in aquatic and terrestrial habitats in Australia. Plant Prot Q 7:102–108
Julien MH, Skarratt B, Maywald GF (1995) Potential geographical distribution of alligator weed and its biological control by Agasicles hygrophila. J Aquat Plant Manag 33:55–60
Keane RM, Crawley MJ (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17:164–170. doi:10.1016/S0169-5347(02)02499-0
Lennartsson T, Tuomi J, Nilsson P (1997) Evidence for an evolutionary history of overcompensation in the grassland biennial Gentianella campestris (Gentianaceae). Am Nat 149:1147–1155
Li W, Wang B, Wang J (2006) Lack of genetic variation of an invasive clonal plant Eichhornia crassipes in China revealed by RAPD and ISSR markers. Aquat Bot 84:176–180. doi:10.1016/j.aquabot.2005.09.008
Lin GL, Yang YZ, Hu JS (1990) Studies on biology and control of Alternanthera philoxeroiders. J Jiangsu Agric Coll 11:57–63
Lu XM, Ding JQ (2010) Flooding compromises compensatory capacity of an invasive plant: implications for biological control. Biol Invasions 12:179–190. doi:10.1007/s10530-009-9441-4
Lu X, Dai H, Ding J (2010) Con-specific neighbours may enhance compensation capacity in an invasive plant. Plant Biol 12:445–452. doi:10.1111/j.1438-8677.2009.00247.x
Ma RY (2001) Ecological adaptation for the intruduced biocontrol agent, Agasicles hygropghila, for Alligatorweed, Alternanthera philoxeroides, in China PhD. Chinese Academy of Agricultural Sciences, Beijing, China, p 120
Mack RN, Thompson JN (1982) Evolution in steppe with few large, hooved mammals. Am Nat 119:757–773
Maron JL, Vilà M (2001) When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 95:361–373. doi:10.1034/j.1600-0706.2001.950301.x
Maschinski J, Whitham TG (1989) The continuum of plant responses to herbivory: the influence of plant association, nutrient availability, and timing. Am Nat 134:1–19. doi:10.1086/284962
McIntire EJB, Hik DS (2002) Grazing history versus current grazing: leaf demography and compensatory growth of three alpine plants in response to a native herbivore (Ochotona collaris). J Ecol 90:348–359. doi:10.1046/j.1365-2745.2001.00672.x
McKey D, Elias M, Pujol B, Duputié A (2010) The evolutionary ecology of clonally propagated domesticated plants. New Phytol 186:318–332. doi:10.1111/j.1469-8137.2010.03210.x
Mitchell CE, Agrawal AA, Bever JD, Gilbert GS, Hufbauer RA, Klironomos JN, Maron JL, Morris WF, Parker IM, Power AG, Seabloom EW, Torchin ME, Vázquez DP (2006) Biotic interactions and plant invasions. Ecol Lett 9:726–740. doi:10.1111/j.1461-0248.2006.00908.x
Monro K, Poore AGB (2004) Selection in modular organisms: is intraclonal variation in macroalgae evolutionarily important? Am Nat 163:564–578. doi:10.1086/382551
Müller-Schärer H, Schaffner U, Steinger T (2004) Evolution in invasive plants: implications for biological control. Trends Ecol Evol 19:417–422. doi:10.1016/j.tree.2004.05.010
Polley HW, Detling JK (1988) Herbivory tolerance of Agropyron smithii populations with different grazing histories. Oecologia 77:261–267. doi:10.1007/BF00379196
Prentis PJ, Wilson JRU, Dormontt EE, Richardson DM, Lowe AJ (2008) Adaptive evolution in invasive species. Trends Plant Sci 13:288–294. doi:10.1016/j.tplants.2008.03.004
Sainty G, McCorkelle G, Julien M (1998) Control and spread of alligator weed Alternanthera pliloxeroides (Mart.) Griseb., in Australia: lessons for other regions. Wetlands Ecol Manage 5: 195–201. doi:10.1023/A:1008248921849
Schierenbeck KA, Mack RN, Sharitz RR (1994) Effects of herbivory on growth and biomass allocation in native and introduced species of Lonicera. Ecology 75:1661–1672. doi:10.2307/1939626
Schooler S, Baron Z, Julien M (2006) Effect of simulated and actual herbivory on alligator weed, Alternanthera philoxeroides, growth and reproduction. Biol Control 36:74–79. doi:10.1016/j.biocontrol.2005.06.012
Schooler SS, Yeates AG, Wilson JRU, Julien MH (2007) Herbivory, mowing, and herbicides differently affect production and nutrient allocation of Alternanthera philoxeroides. Aquat Bot 86:62068. doi:10.1016/j.aquabot.2006.09.004
Siemann E, Rogers WE, Dewalt SJ (2006) Rapid adaptation of insect herbivores to an invasive plant. Proc R Soc B 273:2763–2769. doi:10.1098/rspb.2006.3644
Smith SE (1998) Variation in response to defoliation between populations of Bouteloua curtipendula var. caespitosa (Poaceae) with different livestock grazing histories. Am J Bot 85:1266–1272
Strauss SY, Agrawal AA (1999) The ecology and evolution of plant tolerance to herbivory. Trends Ecol Evol 14:179–185. doi:10.1016/S0169-5347(98)01576-6
Sun Y, Ding JQ, Ren MX (2009) Effects of simulated herbivory and resource availability on the invasive plant, Alternanthera philoxeroides in different habitats. Biol Control 48:287–293. doi:10.1016/j.biocontrol.2008.12.002
Thompson JN (1998) Rapid evolution as an ecological process. Trends Ecol Evol 13:329–332. doi:10.1016/S0169-5347(98)01378-0
Tiffin P (2000) Mechanisms of tolerance to herbivore damage: what do we know? Evol Ecol 14:523–536. doi:10.1023/A:1010881317261
Whitham TG, Slobodchikoff CN (1981) Evolution by individuals, plant-herbivore interactions, and mosaics of genetic variability: the adaptive significance of somatic mutations in plants. Oecologia 49:287–292
Wilsey BJ, Polley HW (2006) Aboveground productivity and root-shoot allocation differ between native and introduced grass species. Oecologia 150:300–309. doi:10.1007/s00442-006-0515-z
Wilson JRU, Yeates A, Schooler S, Julien MH (2007) Rapid response to shoot removal by the invasive wetland plant, alligator weed (Alternanthera philoxeroides). Environ Exp Bot 60:20–25. doi:10.1016/j.envexpbot.2006.06.003
Yang ZH (2001) The establishment of Agasicles hygrophila population in suburb of Shanghai city. Shanghai Agric Sci 1:87–88
Zangerl AR, Berenbaum MR (2005) Increase in toxicity of an invasive weed after reassociation with its coevolved herbivore. Proc Natl Acad Sci USA 102:15529–15532. doi:10.1073/pnas.0507805102
Acknowledgments
We thank Wenfeng Guo, Yi Wang, Xia Jin, Hongjun Dai, Yan Sun and Kai Wu for their field and lab assistance. The manuscript was improved by comments from Richard N. Mack, John Wilson, Heinz Müller-Schärer, David Lodge, Ashley Baldridge, Matthew Barnes, Victoria Nuzzo, Sathyamurthy Raghu, Anjana Dewanji, Jianwen Zou and three anonymous reviewers. This work was funded by the Knowledge Innovation Program of the Chinese Academy of Sciences and the National Science Foundation of China (30871650 & 31100302) while preparing this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lu, X., Ding, J. History of exposure to herbivores increases the compensatory ability of an invasive plant. Biol Invasions 14, 649–658 (2012). https://doi.org/10.1007/s10530-011-0106-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-011-0106-8