
Abstract
Predictive tools for preventing introduction of new species with high probability of becoming invasive in the U.S. must effectively distinguish non-invasive from invasive species. The Australian Weed Risk Assessment system (WRA) has been demonstrated to meet this requirement for terrestrial vascular plants. However, this system weights aquatic plants heavily toward the conclusion of invasiveness. We evaluated the accuracy of the WRA for 149 non-native aquatic species in the U.S., of which 33 are major invaders, 32 are minor invaders and 84 are non-invaders. The WRA predicted that all of the major invaders would be invasive, but also predicted that 83% of the non-invaders would be invasive. Only 1% of the non-invaders were correctly identified and 16% needed further evaluation. The resulting overall accuracy was 33%, dominated by scores for invaders. While the overall accuracy increased to 57% when the points assigned to aquatic life forms were removed, 57% of the non-invaders required further evaluation rather than were identified as having low probability of naturalizing. Discrimination between non-invaders and invaders would require an increase in the threshold score from the standard of 6 for this system to 19. That higher threshold resulted in accurate identification of 89% of the non-invaders and over 75% of the major invaders. Either further testing for definition of the optimal threshold or a separate screening system will be necessary for accurately predicting which freshwater aquatic plants are high risks for becoming invasive.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Across plant taxa, freshwater aquatic and semi-aquatic species have been demonstrated to have high incidence of becoming invasive (Daehler 1998). Opportunities for invasion from this group are rising as trade in aquatic plants has increased in the U.S. and elsewhere because of increasing interest in water gardening, aquaria, and wetland restoration (Maki and Galatowitsch 2004). The numbers of American households with water gardens quadrupled between 1998 and 2003, resulting in $1.56 billion in retail sales for water gardens in 2003 (Crosson 2005). While both native and non-native plants are available, non-native species pose higher risk of invasion when grown or released in new habitats. For example, between 76 and 88% of the non-native aquatic invasive plants in southern New England escaped from cultivation (Les and Mehrhoff 1999). Similarly, 75% of the aquatic invasive plants in New Zealand are of horticultural origin (Champion and Clayton 2000).
The U.S. spent $100 million annually on control of non-native invasive aquatic plants in the 1990s (OTA 1993). National expenditures have likely increased substantially since then; the state of Florida alone spent an estimated $29.7 million to manage only a few species of aquatic non-native invasive plants in fiscal year 2008–2009 (Bureau of Invasive Plant Management 2008). These species also have costly ecological impacts, such as altering water chemistry and temperature, out-competing native species, and impeding flows and movement (e.g., Schmitz et al. 1993; Van et al. 1999; Champion and Clayton 2000; Ciruna et al. 2004). Preventing the introduction of new species likely to have these types of impacts would be far less costly than managing invaders once they are here (OTA 1993; Keller et al. 2007). As a result, development of tools that accurately predict the likely invasiveness of species is critical (Ciruna et al. 2004).
The Australian Weed Risk Assessment system (WRA; Pheloung et al. 1999) was designed to predict that virtually all major invaders would have a high probability of becoming invasive, incorrectly predict that non-invaders would become invasive less than 10% of the time, and require further evaluation of up to 30% of the species (primarily minor and non-invaders). Development of a secondary screen (Daehler et al. 2004) allowed resolution of many species requiring further evaluation. Across the temperate and tropical geographies in which the WRA has been tested, addition of the secondary screen resulted in the accurate identification of major invaders 90% of the time, on average. Non-invaders were accurately identified 70% of the time, on average, with 10% incorrectly identified as invasive and the rest requiring further evaluation (Gordon et al. 2008a). Receiver Operating Characteristic (ROC) analysis confirmed that the WRA differentiates well between non-invaders and invaders despite the prevalence of non-invaders relative to invaders in the full pool of introduced species (the “base rate” sensu Smith et al. 1999; Gordon et al. 2008a).
However, the vast majority of the species used for testing the WRA have been terrestrial and wetland vascular plants. While 5% of the species used in development of the WRA were aquatic (Pheloung 1995 and pers. comm.), almost all other tests of the WRA for which we could find the species lists (Daehler and Carino 2000; Daehler et al. 2004; Kato et al. 2006; Křivánek and Pyšek 2006; Gordon et al. 2008a; McClay et al. 2010) included 0–2% aquatic species. Nishida et al. (2009) were the exception: almost 20% of the 259 species tested for Japan were aquatic.
The WRA has 49 questions, each of which has points that range from −3 to 5 associated with positive or negative responses (Pheloung et al. 1999). The points are added to obtain a conclusion that a species has a low (total score <1; accept) or high (score >6; reject) probability of becoming invasive or requires further evaluation (scores from 1 to 6). The point assignment system reflects the higher invasion probability of species from some groups or life forms. Consistent with the finding of Daehler (1998), grasses, woody nitrogen fixers, and aquatic plants each receive additional points. The highest number of points is associated with aquatic species (5), reflecting the intent of the system to be particularly precautionary about further import of aquatic invaders. This approach is well supported by evidence that out of a potential 363 plant families, half of the 14 families found to have disproportionately high numbers of invasive species were entirely aquatic or semi-aquatic (Daehler 1998).
However, because the majority of introduced aquatic plant species (like their terrestrial counterparts) are not invasive (e.g., Champion and Clayton 2000), we examined whether the WRA may be overly precautionary given the increasing commercial importance of aquatic species in the U.S. An earlier assessment concluded that the WRA might be overly precautionary for New Zealand because questions not relevant for aquatic species (e.g., creates fire hazard) are included, while questions only relevant in aquatic habitats (e.g., threat to navigation) are omitted. These concerns led Champion and Clayton (2000, 2001) to develop a separate risk assessment system for aquatic species. While these authors did evaluate the WRA prior to developing their system, their assessment was limited to 21 species, predominantly invaders. We have expanded their approach to evaluate the WRA for 149 aquatic species, testing the hypothesis that the scoring system results in the prediction that virtually all aquatic plant species will be rejected as likely invaders. If true, over 90% of major aquatic invaders, like major terrestrial invaders, would be correctly rejected, but aquatic non-invaders would have a higher rejection rate than terrestrial non-invaders, unnecessarily restricting potential economic opportunities (Keller et al. 2007). We further examined whether altering the scoring for this group would improve the accuracy of the tool.
Methods
We used published (Coile 1995; Speichert and Speichert 2004) and internet-based aquatic plant lists and consulted with aquatic weed scientists and the aquarium and water garden industries in the U.S. to identify 65 naturalized, and 84 non-invasive aquatic species (“Appendix”). Nomenclature, authorities, and families are primarily from the Missouri Botanical Garden database, Tropicos (http://www.tropicos.org/); we used the Integrated Taxonomic Information system on-line database (www.itis.gov) or specific sources (Crusio 1979; Haynes and Holm-Nielsen 1994; Wunderlin and Hansen 2008) where necessary (see “Appendix”). We defined non-invaders as non-indigenous species that are in international trade but have no record of escaping cultivation in the U.S. Naturalized species, conversely, have reproductive populations outside of cultivation. Encyclopedias of horticulture and water gardening (e.g., Tricker 1897; Bisset 1905; Stodola 1967), on-line vouchered herbarium specimens, and other literature were used to document introduction dates to the U.S. or other countries. Once an introduction date into the U.S. prior to 1960 was obtained, we did not search for earlier dates since we were most interested in whether the species had been in the U.S. for at least 50 years. The species span 55 plant families and include all freshwater macrophyte growth forms (Cook et al. 1974: attached-floating, erect emergent, free-floating, sprawling emergent, submerged). Wetland species that do not also grow under fully aquatic conditions were not included. All species are non-indigenous to the contiguous United States and none were included in the original 371 species used by Pheloung et al. (1999) to develop the WRA.
We further divided the naturalized group into species for which we found record of naturalization, but not invasiveness in the U.S. (minor invaders: 32), and those that are documented to have negative ecological impacts sufficient to result in identification as invasive on regulatory or non-regulatory lists or other materials (major invaders: 33). All minor and major invaders on the list are documented in published floras (e.g., Wunderlin and Hansen 2008) or the USDA PLANTS database (http://plants.usda.gov).
The majority (77%) of the non-invaders has been in the global trade for over 50 years; at least 49% of those have been in the U.S. for that period without naturalizing (“Appendix”). We included more recently introduced species to increase the number of species tested. As a result, some of the non-invaders are only documented to have been in the U.S. for a few years (e.g., Elatine gratioloides). The combined short residence time and potentially low propagule pressure may explain higher than anticipated scores for some species assigned a priori to the non-invader category.
Four questions in the original WRA (Pheloung et al. 1999) were modified to reflect climatic and environmental conditions in the U.S. following Gordon and Gantz (2008). Species were assessed through review of literature and scientific internet sources. Questions were answered consistently with the guidance in Gordon et al. (2010). Because the WRA is intended for predictive use prior to import, we included no data on the invasiveness of the species in the U.S. when scoring those questions. We ran two iterations of the WRA: the points associated with aquatic species (question 5.01) were not modified (5); and no points assigned to aquatic species (0).
A secondary screen, designed to help resolve conclusions for species with scores resulting in the need for further evaluation, was originally developed and applied to a dataset with one aquatic species (Daehler et al. 2004). The screen includes questions about shade tolerance, density of growth, dispersal, and longevity for woody species and vines, and agricultural weediness, palatability to grazers, and density of growth for herbaceous species and vines. The screen was not designed for use on aquatic species (Daehler, pers. comm., 2010), so was not included in this study.
Three non-vascular species (Monosolenium tenerum, Ricciocarpos natans (liverworts) and Vesicularia dubyana (moss)) were included in the assessment because they occur in aquatic habitats in the wild and are used in aquaria or water gardens. For these species, spore data were substituted for seed data in the relevant questions. However, the results of this test of the WRA are primarily generated from vascular aquatic plant taxa.
We compared the mean number of questions answered for species that have been in the U.S. for over 50 years with for the entire dataset using a two-tailed independent t-test. We used a Pearson’s chi-square test to assess the effect of more recent introduction dates than 50 years on the results for non-invaders, testing the null hypothesis that the proportions of non-invaders accepted or rejected is independent of the length of the introduction period. We also used Pearson’s chi-square tests to evaluate differences in accuracy of the WRA on aquatic species compared to results for terrestrial and wetland species without the secondary screen (90% accurate for major invaders, 75% accurate for non-invaders; 10% inaccurate for non-invaders; 20% requiring further evaluation; Pheloung et al. 1999), and assess any differences in accuracy of the WRA when zero, rather than five points were assigned for the aquatic growth form. Our null hypothesis was that the WRA accuracy would be equivalent regardless of the type of species involved but that accuracy might increase if aquatic species were not assigned additional points. The requirement that expected values exceed four was violated for the number of major invaders accepted and the number of non-invaders present for fewer than 50 years that were accepted, so those results should be interpreted with caution. We also evaluated whether the single question of whether the species has naturalized outside its native range (question 3.01) would provide an accurate screening approach.
We evaluated the ability of the WRA to discriminate between non-invaders and invaders using the Receiver Operating Characteristic (ROC) curve. If the area under the ROC curve (AUC) = 1.0, the tool perfectly discriminates between the two categories; if the AUC = 0.5, no discrimination has occurred (Zhang and Pepe 2005; DeLong et al. 1988). We conducted this analysis with minor invaders categorized either as non-invaders or as invaders, since the correct outcome is not certain (Gordon et al. 2008a). We then examined the score that optimized correct identification of both invaders and non-invaders (Youden’s index = Sensitivity – [1 − Specificity]) to identify the threshold score likely to result in the highest WRA accuracy (Bewick et al. 2004; Nishida et al. 2009). Pearson’s chi-square analysis was used to compare the accuracy of the WRA for aquatic plants using the new thresholds with that for terrestrial plants as described above.
Results
Total scores for all species along with their a priori categorization and other data are presented in “Appendix”. We answered an average of 31.6 (range 25–39) of the 49 WRA questions per species. The average number of questions answered was higher (t = 2.83, df = 147, P = 0.005) for species that have been in the U.S. for over 50 years (mean = 32.1, SD = 2.98) than for more recent introductions (mean = 30.8, SD = 2.48). However, the proportions of non-invaders predicted to have a high probability of becoming invasive, not becoming invasive, or needing further evaluation was independent of time since introduction (i.e., whether all 84 non-invaders or only the 32 non-invaders present for over 50 years were included), when either five points (χ2 = 4.575, P = 0.102) or no points (χ2 = 2.85, P = 0.240) were assigned for aquatic species. Additionally, we found no correlation between the total WRA score and the years of introduction for any a priori category (non-invader: r = −0.01, P = 0.90; minor invader: r = −0.32, P = 0.07; major invader: r = −0.14, P = 0.42; Fig. 1). As a result, data for all species were included in the analyses below.

The WRA score corresponding to the first documented U.S. introduction date located for species by a priori category. Five points were assigned to aquatic species. No significant correlations were found (see text)
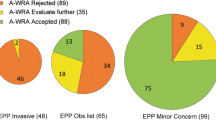
The WRA, using the five points as originally designed, correctly identified 100% of the major invaders (Table 1). Only one of the non-invaders (Cryptocoryne × willisii) was correctly predicted to have a low probability of becoming invasive, 83% were predicted to be invasive, and the remainder required further evaluation. Over 90% of the minor invaders were predicted to have a high probability of becoming invasive. Overall accuracy of predictions (assessed for non- and major invaders only and omitting the species requiring further evaluation) was 33%, dominated by the major invader category (Table 1). Accuracy of the WRA in identifying aquatic major invaders was equivalent to that found for terrestrial species in this group on average (χ2 = 3.300, P = 0.192), but significantly lower (χ2 = 541.5, P < 0.0001) than for terrestrial non-invaders, as hypothesized.
When no points were assigned to aquatic species, the primary result was that more non-invaders received scores of 1–6 rather than >6. As a result, the majority of non-invaders (57%) required further evaluation rather than being predicted to become invasive (35%; Table 2). While we saw a slight increase in the number of non-invaders predicted not to become invasive (8%), accuracy for major invaders decreased slightly to 94% (Table 2). The accuracy for aquatic non-invaders remained lower than for terrestrial species (χ2 = 199.1, P < 0.0001), while that for aquatic major invaders remained equivalent (χ2 = 1.033, P = 0.597).
The single question of whether a species has naturalized outside the U.S. accurately identified 94% of the major invaders and 84% of the non-invaders, resulting in an overall accuracy of 87% (99/114). Eighty-seven percent of the minor invaders would be predicted to be invasive using this question. While inaccuracy might be expected from species that have only recently been introduced into the global trade, the proportions of species predicted to be invasive in each a priori category remained the same when only species present in the trade for at least 50 years were included in the analysis (χ2 = 1.647, P = 0.439).
When minor invaders were categorized as non-invaders, the AUC = 0.88 (SE = 0.033), an area significantly different from 0.5 (P < 0.0001), indicating that the WRA discriminates between non-invaders and invaders. Similarly, when minor invaders were grouped with major invaders, the AUC = 0.90 (SE = 0.028, P < 0.0001). These AUC values are not different than those found for terrestrial species (Gordon et al. 2008a); however, the Youden Index demonstrates that this discrimination occurs at a threshold score of 19 or 16, when minor invaders are considered non-invasive or invasive, respectively.
The WRA accuracy increased when the Youden index values were used to identify threshold scores for predicting which species have a low (score <16) or high (score >19) risk of invasion (Table 3). We used the score ranges influenced by the minor invaders (16–19) to identify species that required further evaluation. Accuracy for non-invaders using these thresholds was significantly higher (χ2 = 9.56, P = 0.008) than for the WRA prediction of terrestrial species, while that for major invaders was lower (χ2 = 11.83, P = 0.003). Using this approach, almost 40% of aquatic minor invaders were rejected, compared to 60% of terrestrial species in this category (Table 3). Fourteen percent of the species required further evaluation.
Discussion
These results confirm that the WRA as originally developed is overly precautionary in its prediction of the likely invasiveness of aquatic plants, and that simply eliminating the “penalty” points assigned to aquatics does not produce a sufficiently accurate screening system for this group. While accuracy was high for the major invaders, essentially all of which were predicted to be invasive, the majority of non-invaders were predicted either to be invasive or required further evaluation. Only between 1 and 8% of the non-invaders were correctly predicted to be non-invasive using the WRA (Tables 1, 2).
While minor invaders were not used to calculate accuracy as described above, the WRA thresholds were understood to predict that roughly 60% of species in this category would be rejected, with the remainder requiring further evaluation (Pheloung et al. 1999). Other tests of the WRA confirmed this result (Gordon et al. 2008a). However, we found a significantly higher proportion of aquatic minor invaders were rejected regardless of the points assigned to this group (Tables 1, 2). Again, this tool appears overly precautionary when applied to aquatic species.
Decreasing the point penalty for aquatic plants, from the existing five points to no points, reduced the number of rejected non-invaders. However, the accuracy of these results was not increased because the majority of species then required further evaluation. Development of a secondary screening tool for aquatic species similar to that developed for terrestrial species (Daehler et al. 2004) could assist in developing a clear outcome for a higher proportion of these species.
Alternatively, leaving the WRA scoring as intended, but changing the threshold that distinguishes invaders from non-invaders resulted in significantly higher accuracy of this tool (Table 3). Development of the original thresholds was based on data from 371 species (Pheloung et al. 1999); we would recommend increasing the sample size of aquatic species tested before assuming that our empirically determined thresholds are appropriate for these life forms. However, the lower accuracy for identifying major invaders than found in most tests of the WRA with terrestrial species (Gordon et al. 2008a) suggests that development of a risk assessment tool specifically for aquatic plants (e.g., Champion and Clayton 2000) might be a better approach.
The one WRA study that included a significant proportion of aquatic plant species was for Japan (Nishida et al. 2009). Both native and non-native plants were included in this assessment. Of the 23 aquatic species categorized as major invaders, 100% were rejected, 79% of which had scores of 16 or higher. Similarly, all 25 (100%) minor invaders were rejected (scores of all rejected species were >10, reflecting the Youden’s Index identified for their dataset). The one non-invasive aquatic species included in the study had a score of four. The higher score threshold for rejection derived in this study may have partially resulted from the aquatic species included in the test. Although insufficient non-invaders were included for a clear comparison with our data, Nishida et al. (2009) results appear consistent with our findings.
Interestingly, the estimated year of introduction into the U.S. did not significantly influence the accuracy for the non-invaders. Similarly, the number of years a species has been in the global trade did not influence whether species naturalized elsewhere were likely to be invasive in the U.S. Although factors like residence time, cultivation, and propagule pressure have been documented to influence the probability of invasion (Mack 2000; von Holle and Simberloff 2005), the date of introduction appeared to have little influence on the score or accuracy. Most of these aquatic species are herbaceous and reproduce asexually as well as sexually. These characteristics may mean that invasive capacity is expressed more rapidly for this group than terrestrial species (e.g., Kowarik 1995). Aquatic species may be more like many tropical species, which can become invasive within a decade (Daehler 2009).
Some of the species apparently non-invasive in the U.S. may have a high probability of becoming invasive over time. Whether or not a species has been invasive elsewhere has repeatedly been demonstrated to be the best predictor of whether it will become invasive in a new habitat (Gordon et al. 2008b) and was a more accurate predictor of invasiveness for these aquatic species n the U.S. (87%) than was the full WRA (34–58%). As a result, apparent non-invaders with records of invasiveness elsewhere outside their native range may be species that need proactive management or regulation. For example, in our dataset, Limnocharis flava, and Typha orientalis show no evidence of being invasive in the U.S. but are major invaders elsewhere. Limnocharis flava (Sawyer and Perkins 1934) has been in the global trade and in the U.S. for over 80 years and may have already expressed any invasive potential of the genetic material present. However, while T. orientalis has been in the global trade since at least 1951 (Chittenden 1951), we found no record of it in the U.S. before 1986 (Denver Botanic Gardens: http://rbg-eb2.rbge.org.uk/multisite/multisite3.php). Given the aggressive colonization of this species in western Australia where it is not native (Zedler et al. 1990), it should be monitored for indication of incipient invasiveness in the U.S.
We did not find evidence that all species from a single genus were either invasive or not. While we could not fully test this hypothesis, as the species list was developed to maximize family and growth form diversity rather than species within genera, we found 12 genera with species in both the a priori invader and non-invader categories and 20 genera in which we tested multiple species that all fell within one a priori category (“Appendix”). The assumption that invasiveness may be predicted from the genus is not supported by these data (see also Burns 2006).
The need for a screening system for aquatic plants in the U.S. and other countries is clear (Ciruna et al. 2004). In New Zealand, 27% of the introduced freshwater aquatic plant species have naturalized, 50% of which have become invasive (Champion and Clayton 2000). The numbers are likely to be similar across other geographies with multiple types of aquatic habitats and environments. Countries or regions unable to be as precautionary as is Australia might use the WRA but increase the threshold score needed for rejection, as suggested by these data. Given the relatively high accuracy of the single question of whether the species has naturalized elsewhere and the rapidity of exhibiting that characteristic, this question could be a sufficient preliminary screening approach for any species with a history of introduction beyond its native range. However, increased prediction accuracy would likely result from a system specifically designed for aquatic plant species. Testing or refining the system used in New Zealand for accuracy in other countries (e.g., Champion et al. 2008) may facilitate development of an effective prevention mechanism.
References
Bewick V, Cheek L, Ball J (2004) Statistics review 13: receiver operating characteristic curves. Crit Care 8:508–512
Bisset P (1905) The book of water gardening. A. T. De la Mare, New York, p 199
Bureau of Invasive Plant Management (2008) Status of the Aquatic Plant Maintenance Program in Florida Public Waters. Annual Report Fiscal Year 2006–2007. Florida Department of Environmental Protection, Tallahassee, Florida. http://www.myfwc.com/docs/WildlifeHabitats/InvasivePlants_Aquatic06-07.pdf. Accessed May 20, 2009
Burns JH (2006) Relatedness and environment affect traits associated with invasive and noninvasive introduced Commelinaceae. Ecol Appl 16:1367–1376
Champion PD, Clayton JS (2000) Border control for potential aquatic weeds. Stage 1. Weed risk model. Science for conservation 141. Department of Conservation, Wellington, New Zealand
Champion PD, Clayton JS (2001) A weed risk assessment model for aquatic weeds in New Zealand. In: Groves RH, Panetta FD, Virtue JG (eds) Weed risk assessment. CSIRO Publishing, Victoria, Australia, pp 194–202
Champion PD, Burnett DA, Petroeschevsky A (2008) Risk assessment of aquatic plant trade species in Australia. Report to: New South Wales Department of Primary Industries and National Aquatic Weeds Management Group, NIWA Client Report AUS2008/001, NIWA Project NAU07901, Western Australia, Australia
Chittenden FJ (ed) (1951) The royal horticultural society dictionary of gardening, vol 4. Clarendon Press, Oxford, p 604
Ciruna KA, Meyerson LA, Gutierrez A (2004) The ecological and socio-economic impacts of invasive alien species in inland water ecosystems. Report to the Convention on Biological Diversity on behalf of the Global Invasive Species Programme, Washington, D.C., p 34
Coile NC (1995) Common plants of Florida’s aquatic plant industry. Florida Department of Agriculture and Consumer Services, Division of plant industry, Gainesville, Florida, p 132
Cook CDK, Gut BJ, Rix EM, Schneller J, Seitz M (1974) Water plants of the world. Dr. W Junk BV Publishers, The Hague, p 561
Crosson H (2005) Keeping aquatic plants in their place: Common sense tips to protect lakes and rivers. LandscapeOnline.com. http://www.plantright.org/library/pdfs/Crosson2005.pdf. Accessed May 20, 2009
Crusio W (1979) A revision of Anubias Araceae Primitiae Africanae 12. Meded Landbouwhogeschool Wageningen 79(14):1–48
Daehler CC (1998) The taxonomic distribution of invasive angiosperm plants: ecological insights and comparison to agricultural weeds. Biol Conserv 84:167–180
Daehler CC (2009) Short lag times for invasive tropical plants: evidence from experimental plantings in Hawai’i. PLoS ONE 4(2):e4462. doi:10.1371/journal.pone.0004462
Daehler CC, Carino DA (2000) Predicting invasive plants: prospects for a general screening system based on current regional models. Biol Invasions 2:93–102
Daehler CC, Denslow JS, Ansari S, Kuo H (2004) A risk-assessment system for screening out invasive pest plants from Hawaii and other Pacific islands. Conserv Biol 18:360–368
DeLong ER, DeLong DM, Clarke-Pearson DL (1988) Comparing the areas under two or more correlated receiver operating characteristic curves: a nonparametric approach. Biometrics 44:837–845
Gordon DR, Gantz CA (2008) Potential impacts on the horticultural industry of screening new plants for invasiveness. Conserv Lett 1:227–235
Gordon DR, Onderdonk DA, Fox AM, Stocker RK (2008a) Accuracy of the Australian weed risk assessment system across varied geographies. Divers Distrib 14:234–242
Gordon DR, Onderdonk DA, Fox AM, Stocker RK, Gantz C (2008b) Predicting invasive plants in Florida using the Australian weed risk assessment. Invasive Plant Sci Manag 1:178–195
Gordon DR, Mitterdorfer B, Pheloung PC, Ansari S, Buddenhagen C, Chimera C, Daehler CC, Dawson W, Denslow JS, LaRosa A, Nishida T, Onderdonk DA, Panetta FD, Pyšek P, Randall RP, Richardson DM, Tshidada NJ, Virtue JG, Williams PA (2010) Guidance for addressing the Australian weed risk assessment questions. Plant Prot Q 25(2):56–74
Haynes RR, Holm-Nielsen LB (1994) The Alismataceae. In: Flora Neotropica 64:1–112. New York Botanical Garden Press on behalf of Organization for Flora Neotropica, New York
Kato H, Hata K, Yamamoto H, Yoshioka T (2006) Effectiveness of the weed risk assessment system for the Bonin Islands. In: Koike F, Clout MN, Kawamichi M, De Poorter M, Iwatsuki K (eds) Assessment and control of biological invasion risk. Shoukadoh Book Sellers, Kyoto, Japan and IUCN, Gland, Switzerland, pp 65–72
Keller RP, Lodge DM, Finnoff DC (2007) Risk assessment for invasive species produces net bioeconomic benefits. PNAS 104:203–207
Kowarik I (1995) Time lags in biological invasion with regard to the success and failure of alien species. In: Pyšek P, Rejmánek M, Wade M (eds) Plant invasions—general aspects and special problems. SPB Academic Publishing, Amsterdam, pp 15–38
Křivánek M, Pyšek P (2006) Predicting invasions by woody species in a temperate zone: a test of three risk assessment schemes in the Czech Republic (Central Europe). Divers Distrib 12:319–327
Les DH, Mehrhoff LJ (1999) Introduction of nonindigenous aquatic vascular plants in southern New England: a historical perspective. Biol Invasions 1:281–300
Mack RN (2000) Cultivation fosters plant naturalization by reducing environmental stochasticity. Biol Invasions 2:111–122
Maki K, Galatowitsch S (2004) Movement of invasive aquatic plants into Minnesota (USA) through horticultural trade. Biol Conserv 118:389–396
McClay A, Sissons A, Wilson C, Davis S (2010) Evaluation of the Australia Weed Risk Assessment system for the prediction of plant invasiveness in Canada. Biol Invasions 12: 4085–4098
Nishida T, Yamashita N, Asai M, Kurokawa S, Enomoto T, Pheloung PC, Groves RH (2009) Developing a pre-entry weed risk assessment system for use in Japan. Biol Invasions 11:1319–1333
Office of Technology Assessment (1993) Harmful non-indigenous species in the United States. U.S. Congress/OTA-F-565, U.S. Government Printing Office, Washington, D.C
Pheloung PC (1995) Determining the weed potential of new plant introductions to Australia. A report on the development of a Weed Risk Assessment System commissioned by the Australian Weeds Committee and the Plant Industries Committee, Perth, Western Australia
Pheloung PC, Williams PA, Halloy SR (1999) A weed risk assessment model for use as a biosecurity tool evaluating plant introductions. J Environ Manage 57:239–251
Sawyer RV, Perkins EH (1934) Water gardens and goldfish. New York, A. T. De La Mare, New York and General Publishing Company, Ltd, Toronto, Canada, p 259
Schmitz DC, Schardt JD, Leslie AJ, Dray FA, Osborne JA, Nelson BV (1993) The ecological impact and management history of three invasive alien aquatic plant species in Florida. In: McKnight BN (ed) Biological pollution: the control and impact of invasive exotic species. Indiana Academy of Science, Indianapolis, Indiana, pp 173–194
Smith CS, Lonsdale WM, Fortune J (1999) When to ignore advice: invasion predictions and decision theory. Biol Invasions 1:89–96
Speichert G, Speichert S (2004) Encyclopedia of water garden plants. Timber Press, Portland, Oregon, p 386
Stodola J (1967) Encyclopedia of water plants. TFH Publications, Neptune City, New Jersey, p 368
Stuckey RL, Salamon DP (1987) Typha angustifolia in North America: a foreigner masquerading as a native. Am J Bot 74:757
Tricker W (1897) The water garden. AT De La Mare, New York, p 120
Van TK, Wheeler GS, Center TD (1999) Competition between Hydrilla verticillata and Vallisneria americana as influenced by soil fertility. Aquat Bot 62:225–233
Von Holle B, Simberloff D (2005) Ecological resistance to biological invasion overwhelmed by propagule pressure. Ecology 86:3212–3218
Wunderlin RP, Hansen BF (2008) Atlas of Florida Vascular Plants. [Landry SM, Campbell KN (application development), Florida Center for Community Design and Research.] Institute for Systematic Botany, University of South Florida, Tampa. http://www.florida.plantatlas.usf.edu/. Accessed May 6, 2009
Zedler JB, Paling E, McComb A (1990) Differential salinity responses help explain the replacement of native Juncus kraussii by Typha orientalis in Western Australian salt marshes. Austral J Ecol 15:57–72
Zhang Z, Pepe MS (2005) A linear regression framework for receiver operating characteristic (ROC) curve analysis. Washington: University of Washington Biostatistics Working Paper Series, paper 253
Acknowledgments
We appreciate J. Gardina and J. Chase’s assistance with literature review and for help with identification of aquatic plant species for this test from D. Keller, E. Elgin, J. Chase, R. Keller, J. Schardt, D. Schmitz, and R. Weaver. Comments from W.L. Chadderton, P. Champion, O. Harrod, C. Jacono, B. Mitterdorfer, D. Onderdonk, P. Pheloung and anonymous reviewers have improved this paper. This work was completed with funding from the Florida Fish and Wildlife Conservation Commission, Invasive Plant Management Section, Department of Defense, Army Corps of Engineers, and Florida and Michigan Chapters of The Nature Conservancy. The conclusions reported here are those of the authors and do not necessarily reflect the position or policy of the sponsoring agencies or organization.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gordon, D.R., Gantz, C.A. Risk assessment for invasiveness differs for aquatic and terrestrial plant species. Biol Invasions 13, 1829–1842 (2011). https://doi.org/10.1007/s10530-011-0002-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-011-0002-2