
Abstract
Invasive alien trees along river banks can reduce indigenous biodiversity, while their removal can restore it. We assessed here family- and species-level responses of river benthic macroinvertebrate assemblages to three riparian vegetation types (natural, alien trees, cleared of alien trees) in the Cape Floristic Region biodiversity hotspot. High species beta diversity of this highly endemic fauna meant that between-river, as well as seasonal effects, dominated assemblage patterns. SASS5, a qualitative, rapid bioassessment technique, based on the sensitivity of the families present, was used as a measure of river health and, indirectly, of water quality. SASS indicated a decline in water quality conditions after alien clearing, a likely response to the greater insolation and apparent erosion of cleared banks, resulting in elevated temperatures and suspended solids and lowered oxygen levels. Overall, cleared and natural sites were more similar to each other than to alien sites, suggesting some post-clearing recovery. However, many sensitive, endemic taxa survived in alien-invaded sites, and more than in the natural sites. These endemic species made use of shady, cool, high-oxygen levels under the alien tree canopy. However, endemics declined in overall abundance in sites cleared of aliens, being replaced by more tolerant, widespread taxa. Clearance of the alien trees opened up the rivers to sunny conditions, which had a major impact on community composition. Vegetation types, oxygen levels and river width were important environmental variables affecting these macroinvertebrate responses. Re-establishment of invertebrate biodiversity matched that of indigenous vegetation, with the most sensitive endemic taxa only recovering after establishment of bushy indigenous and shade-producing fynbos. Therefore, for biodiversity conservation objectives to be achieved, it is essential that indigenous plants are maintained and encouraged during and after clearing to ensure the recovery of endemic and sensitive taxa.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Little is known of the impact of invasive alien vegetation on highly endemic invertebrate faunas, such as those of rivers of the Cape Floristic Region (CFR), a global biodiversity hotspot (Myers et al. 2000). Dense stands of invasive trees along river margins change habitat structure, fire regimes, hydrology, and water quality, which in turn changes light regime, temperature, oxygen levels, leaf litter input, nutrient cycling, pH, turbidity, soil chemistry, erosion processes and sedimentation (Rutt et al. 1989; Davies and Day 1998; Maridet et al. 1998; Fischer et al. 2010). Alien trees, especially Acacia spp., have dense canopies that shade out habitats and prevent growth of indigenous under storey plants such as sedges, reeds and grasses, altering availability and quality of marginal habitats (Kinvig and Samways 2000; Samways et al. 1996) and soil (Fischer et al. 2010), causing a loss of marginal vegetation often used by benthic macroinvertebrate larvae as nursery or feeding areas or for shelter, and by adults as perches or oviposition sites. Alien riparian vegetation therefore affects both riparian and aquatic habitats.
The systematic removal of alien vegetation is being conducted by the national Working for Water Programme and is expected to considerably increase the long-term survival of rare and endemic species. Alien riparian trees are initially cleared by felling and burning, while follow-up treatments use herbicides and burning to prevent sprouting. The removal of alien trees is therefore a further disturbance to the river system, and may cause additional changes to water quality, and to in-stream and riparian habitats (Bunn et al. 1999). There is also a reduction in shade, resulting in increased water temperatures, thereby reducing the solubility of oxygen. Temperatures also fluctuate more widely after alien removal (Allan and Flecker 1993). Greater exposure to sunlight results in the growth of macrophytes and filamentous algae which are less palatable to herbivores (Bunn et al. 1999). Removal of riparian vegetation also increases runoff from catchments, decreases bank stability, increases erosion, and changes water quality, with concurrent changes of in-stream habitat as a result of changes in flow regime, substrate modification and vegetation changes (Vuori and Joensuu 1996). It is likely therefore, that alien tree clearing will affect, either directly or indirectly, benthic macroinvertebrate communities.
The CFR has high species beta diversity and many endemic freshwater species, many of which remain undescribed or undetected (Wishart et al. 2003). Descendants of temperate Gondwanan fauna, 64% of freshwater invertebrates in the CFR are endemic (Wishart and Day 2001). Yet this area is also heavily impacted by invasive alien plants (Richardson and van Wilgen 2004), raising concern for this irreplaceable fauna. It is this concern that prompted this study.
Apart from the preliminary work of Lowe et al. (2008), no previous research has determined the effects of alien tree invasion and removal on the highly endemic aquatic macroinvertebrate communities of the CFR. Therefore, we aim here to assess the effects on riparian habitats and benthic macroinvertebrate communities of as a result of alien tree invasion, and extent of recovery where the alien trees have been removed, in comparison with natural, untransformed sites in the same catchments.
Materials and methods
Study area
The Mediterranean-type CFR vegetation in the upper catchments is mostly sclerophyllous fynbos, on quartzitic sandstone, and is characteristically acidic and low in nutrients and dissolved solids (Day and King 1995). Each river and catchment in the CFR has its own signature of benthic macroinvertebrate fauna, resulting from a long period of geological and climatic stability coupled with isolation of individual catchments, and high levels of endemism (King and Schael 2001; Wishart et al. 2003). This suggested that more than one catchment should be studied to establish whether the alien tree removal and impacts are similar or not between catchments. We chose the upper regions, but without invoking the effects of zonation, of the Molenaars and Rondegat Rivers (Fig. 1), above any impoundments and where the only major disturbance was that of invasive alien trees.

Location of the two Western Cape rivers studied. 1 = Rondegat River, 2 = Molenaars River
Along each river, three vegetation types were identified: natural (indigenous riparian vegetation), alien (invasive alien trees) and cleared (alien trees removed >2 years previously). Five sites were selected from each of the three vegetation types at each river (i.e. 15 sites per river). Each site was 15 m long and was spaced at intervals of >20 m. We sampled during January and again in March 2004, generating a total of 60 samples. Alien trees included Acacia mearnsii De Willd., with some A. longifolia (Andr.) Willd. and Eucalyptus camaldulensis Dehnh., accounting for 70% cover at some sites. Cleared sites along the Molenaars River were characterized by palmiet reed, Prionium serratum L.f. (‘Prionium’) and short indigenous trees and bushes, while sedges were common in Rondegat cleared sites. Most natural sites had riparian thicket (consisting mostly of Brabejum stellatifolium L. and Metrosideros angustifolia [L.] Smith), referred to here as ‘tall indigenous trees’ to separate them from shorter indigenous bushes. Shading by these trees reduced under storey growth of Prionium and grasses, although fire in natural sites along the Rondegat River resulted in an increase in Prionium and short fynbos elements.
Benthic macroinvertebrate sampling
Numerous bioassessment methods using benthic macroinvertebrates have been developed (e.g. Chessman 1995; Chutter 1972; Rosenberg and Resh 1993; Wright et al. 1984). Biotic indices have been widely applied to stream ecosystems and, as well as providing an indirect measure of water quality (Rutherfurd et al. 2000; Simaika and Samways 2010), can also be used to measure disturbance, including that of agriculture and afforestation (Quinn et al. 1997). Using the South African Scoring System, a biotic index based on sampling macroinvertebrates, a score was assigned to each family, based on known sensitivity/tolerance of that family to changes in water condition (Dickens and Graham 2002). Sensitive invertebrate taxa used in macroinvertebrate scores include Ephemeroptera, Plecoptera, Trichoptera (EPT taxa) (Rosenberg and Resh 1993) and the Odonata (Dickens and Graham 2002; Smith et al. 2007; Simaika and Samways 2009, 2010; Samways and Sharratt 2010) (EPTO taxa), and were therefore a focus here.
Sampling methods and analyses
Benthic invertebrates were sampled at each site using the standardised SASS5 method (Dickens and Graham 2002). Ten replicates were collected from each site, five during January and five during March. All available biotopes were sampled: stones-in-current and stones-out-of-current (sampled collectively), marginal and aquatic vegetation (sampled collectively), gravel, sand and mud (sampled collectively) and handpicking or visual observation. Stones were kicked for 2–5 min, while a kick net (300 × 300 mm frame, 950 μm mesh) was held immediately downstream of the sampled area. Vegetation was sampled by sweeping for a total of two meters through the vegetation below the water level. Loose substratum was agitated for 30 s. Collected macroinvertebrates were tipped into a sorting tray and identified to species, family or higher taxon. EPTO taxa were collected, preserved in 80% ethanol and later sorted to species. Individuals of each family and species were counted. The macroinvertebrate (SASS5) score was then calculated according to families (or higher taxon for non-insect taxa) present. The total of these scores was the divided by the number of taxa present, giving the Average Score Per Taxon (ASPT). The ASPT is the standardized and therefore more robust measure (Dallas and Day 2007; Simaika and Samways 2010).
Riparian vegetation was classified into broad categories: sedges/grasses, alien Acacia spp., Prionium and indigenous trees and bushes (mostly M. angustifolia and B. stellatifolium but also including ericoid and proteoid fynbos shrubs, such as Erica caffra L. and Brachylaena neriifolia [L.f.] R. Br).
At each site, the following environmental variables (EVs) were measured: (1) river system (in increasing latitude: 1 = Rondegat, 2 = Molenaars, spanning less than 17′ latitude apart within the 19° longitude), (2) disturbance regime (1 = natural, 2 = cleared, 3 = alien), (3) dissolved oxygen, temperature, pH and electrical conductivity (with a YSI 556MPS water analyzer), (4) flow (time taken for a floating object to be carried 5 m), (5) mean width and depth of river, (6) % cobbles, gravel and sand in river substrate, (7) % cover of each vegetation category given above (in a 2 m marginal recording zone), (8) % cover of exposed soil on banks (in a 1 m marginal recording zone), (9) % canopy cover (% of river covered by trees), (10) coarse particulate organic matter (CPOM) (scored between one and five according to the amount of debris collected in the SASS sample), (11) total suspended solids (TSS) and, (12) percentage of the water surface with riffles (broken flow) and pools (no visible flow). River water was collected from high-flow areas and filtered through pre-weighed 45 μm GF/F filters within 24 h of sampling. Filters were oven-dried at 40°C for 24 h, re-weighed and the difference calculated.
Data analyses
Data analyses involved: (1) qualitative water quality assessment using macroinvertebrate scores, and (2) quantitative and multivariate analysis to determine community responses. The latter was performed at two levels of taxonomic complexity: family and species. Family and species abundances were calculated for EPTO only. The ASPT scores were compared between sites with cleared, alien and natural riparian vegetation. ANOVA and student’s t-test compared means. Results were considered significant at P < 0.05. Because macroinvertebrate scores do not conform to all the assumptions of normality, the Kruskal-Wallis test was used to confirm t-test results. The results were found to be consistent with those of the Kruskal-Wallis test, so only t-test results are presented. Mean abundance and richness were calculated for alien, cleared and natural sites in both rivers. Analysis of variance and student’s t-test were used to analyse differences between means. Multivariate community analysis of abundance data was done using PRIMER v5.0 software (Clarke and Warwick 2001). Sites with similar assemblage patterns were identified and grouped together in habitat clusters. This was achieved with the CLUSTER programme within the PRIMER software package. Hierarchical agglomerative clustering was performed on Bray-Curtis similarity matrices, using group-average linking, to produce dendrograms (Bray and Curtis 1957). The data were fourth-root transformed to remove heterogeneity of variance (Underwood 1998). Non-metric multi-dimensional scaling (NMDS) visually displayed the relationships between the sites in the ordination analysis.
Analysis of similarity (ANOSIM) was used to establish significance of differences between sites and habitat clusters. ANOSIM is a non-parametric permutation procedure applied to the similarity matrix which underlies the ordination. This method overcomes the problem of non-normal probability distributions resulting from dominance of zero values in multispecies abundance data (Clarke and Warwick 2001). Family or species assemblages responsible for distinguishing habitat clusters were identified with the SIMPER (similarity percentages) routine, that examines overall percentage contribution each taxon makes to average dissimilarity between habitat clusters. SIMPER also highlights the assemblages which contribute most to within-group similarities, these taxa therefore being characteristic of that habitat cluster. EVs that were important in determining faunal composition were determined with canonical correspondence analysis (CCA) using CANOCO version 4.5 software (ter Braak and Ŝmilauer 2002). This is a direct gradient analysis technique which uses multiple regression to select linear combinations of EVs that account for most of the variation in taxon scores on each axis.
‘Vegetation type’ (i.e. natural, cleared or alien) and ‘river system’ were included as nominal variables in the CCA. For the ordination diagrams, each vegetation type was converted to a dummy variable (with a value of one when present in a sample, and zero when absent) and represented as a centroid (average scores of samples belonging to that class). In addition to overall CCA, a partial ordination was performed to eliminate between-river and seasonal effects. This was achieved by specifying ‘river’ and ‘month’ as co-variables. Partial ordination yields an ordination diagram of residual variation in taxon data after the covariable is factored out by multiple linear regression.
Forward selection was used to rank EVs in order of importance according to eigenvalues produced (i.e. variation in taxon data accounted for by that variable) when each EV was considered individually. Monte Carlo permutation tests, using 199 unrestricted random permutations, tested significance of EVs on the fauna. EVs were first selected by forward selection and each tested individually (marginal effects). EVs were then tested collectively with addition of one EV at a time (conditional effects). Monte Carlo permutation tests replaced F- and t-tests in forward selection in multiple regression.
Results
Macroinvertebrate scores
Mean ASPT scores were highest in alien sites but there was no overall significant difference between vegetation types (F = 2.66, P = 0.08) (Table 1). However, the Rondegat River alone did show a significant difference between vegetation types, with natural site scores being significantly higher than cleared or alien site scores. The ASPT differed significantly between rivers, with the Molenaars River having the highest mean score.
Richness and abundance
There was no significant difference between mean number of families in each vegetation type (F = 1.62, P = 0.19). Species richness also did not differ significantly between vegetation type (F = 1.84, P = 0.15) but was slightly lower in alien sites (Table 2). There were 10 localized endemic species in each of the alien and cleared vegetation types, yet only eight in the natural vegetation. Lestagella penicillata (Teloganodidae), Baetis harrisoni, Pseudocloeon bellum, Demorepus capensis (Baetidae) and Pseudagrion A (probably endemic P. furcigerum) (Coenagrionidae) had significantly higher mean abundance in natural and cleared, than in alien sites. Afronurus harrisoni (Heptageniidae) and baetid C had the highest mean abundance in natural sites (Table 2). Baetid B and Chloroterpes B (Leptophlebiidae) had significantly high abundance in cleared sites, while Caenis capensis (Caenidae), Aeshna sp. (probably national endemic A. subpupillata) (Aeshnidae) and Cheumatopsyche sp. (Hydropsychidae) had significantly low abundances. Considering only endemics, L. penicillata had the highest mean abundance in natural sites. Indeed, L. penicillata was significantly more abundant in cleared and natural sites than in alien sites, while Athripsodes C was significantly more abundant in the cleared sites than in the natural or alien sites (Table 2).
Drivers of assemblage patterns
An ANOSIM (two-way crossed design with no replication) gave a statistical significance between rivers (R = 0.582, P = 0.1%) and between months (R = 0.460, P = 0.1%), but not overall between vegetation types (R = 0.130, P = 0.2%). However, there was a significant difference between natural and alien vegetation (R = 0.194, P = 0.1%) and between cleared and alien vegetation (R = 0.170, P = 0.2%), but no significant difference between cleared and natural vegetation (R = 0.094 respectively, P > 1%).
An NMDS ordination diagrams of sites according to species abundance showed a strong grouping according to rivers (Fig. 2a), but not according to vegetation type (Fig. 2b). Cleared and alien groups, in particular, showed great variability, while natural groups were more defined. Sites also grouped according to dominant vegetation composition (Fig. 2c). Indigenous bush and Prionium groups were evident. Dominant vegetation in cleared sites ranged from grasses and Prionium through to short and tall indigenous bushes and fynbos. Natural sites only had indigenous bushes and trees or Prionium as dominant vegetation type, grasses never dominating these sites. Therefore, cleared sites with indigenous bushes as dominant vegetation type were most similar to natural sites. These were followed by cleared sites with an abundance of Prionium. Cleared sites dominated by grasses and sedges were outliers and were least similar to natural sites.

NMDS ordination diagrams of morphospecies abundance data across sites, grouped according to river (a), vegetation type (b) and dominant vegetation composition (c). M Molenaars River, R Rondegat River, N natural, C cleared, A alien. A stress level of <0.2 indicates a useful representation
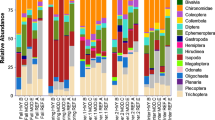
A SIMPER analysis of rivers and months was used to determine which taxa were responsible for these between-river and seasonal differences in invertebrate assemblages (Tables 3 and 4). In particular, Teloganodidae and Baetidae were far more abundant in the Molenaars River, while Caenidae and Gomphidae were more common in the Rondegat River. Species responsible for seasonal and between-river differences are listed in Tables 3 and 4. L. penicillata was primarily responsible for these differences. Euthraulus elegans also differed greatly between rivers and months. Most species were more abundant in the Molenaars River than the Rondegat River, with only Caenis capensis and Paragomphus sp. being more abundant in the Rondegat River. Most distinguishing families and all species were more abundant in March than in January (Table 4). Because of these between-river and seasonal differences, effects of alien disturbance on distribution patterns were analysed within each river and for each month. Characteristic families overlapped considerably between vegetation types. Moreover, few families consistently characterised a particular vegetation type in both rivers and during both months. Baetidae were characteristic of all disturbance regimes, with 15–20% contribution to overall abundance, whereas Heptageniidae characterised both cleared and natural sites.
Only Baetidae, Heptageniidae and Leptoceridae consistently characterised natural sites in both rivers and during both months, but they also characterised cleared and alien sites in at least one river. Similarly, Leptophlebiidae and Hydropsychidae consistently characterised alien sites, but also characterised natural and cleared sites in at least one river. Caenidae was characteristic of Rondegat sites only, and Libellulidae was characteristic of cleared sites along the Rondegat River only. Teloganodidae was clearly characteristic of natural sites along both rivers, but only during March. Although Teloganodidae also occurred in alien and cleared sites along the Molenaars River during March, it was less abundant in these sites.
A SIMPER analysis of species also showed great overlap between characteristic assemblages in each vegetation type (Table 5). There was little consistency across rivers and months, although B. harrisoni and A. harrisoni were consistently characteristic of all three vegetation types in both rivers and during both months. Baetid ‘C’ and P. bellum were characteristic of all natural sites, while Cheumatopsyche sp. was consistently characteristic of alien sites. However, all overlapped with other vegetation types in at least one river. When only March was considered, L. penicillata, P. bellum and Cheumatopsyche sp. characterised natural sites in both rivers and E. elegans was characteristic of alien sites. However, only L. penicillata showed little overlap with other vegetation types, having a markedly higher abundance in natural sites. With the exception of the baetids, P. bellum and baetid C, these characteristic species were either the most abundant representative of their families (i.e. Cheumatopsyche sp., E. elegans, B. harrisoni) or the only species within their family (i.e. L. penicillata, A. harrisoni). It is likely, therefore, that analysis at family level would give sufficient information.
To remove seasonal and between-river effects, ‘river’ and ‘month’ were partialled out as covariables in the CCA. This ordination of species across sites showed L. penicillata, P. bellum, A. harrisoni, baetid E, Anax sp., P. bellum and Pseudagrion A to be associated mainly with natural sites. Baetid C was associated mainly with cleared sites but its position near the origin indicated overlap between vegetation types. Paragomphus sp., Ecnomus sp., Orthetrum sp., Trithemis A and B, Hydroptila capensis and Pseudagrion B were associated most strongly with cleared sites, while E. elegans and Adenophlebia sp. were associated mainly with alien sites.
Response of invertebrates to environmental factors
‘River’ and ‘month’ were the most important EVs (eigenvalues of 0.06). Therefore, CCA was performed with these variables partialled out as covariables, thus negating seasonal and between-river effects. The Monte Carlo test confirmed that measured EVs accounted for a significant amount of variation of the first axis (F = 4.109, P = 0.004) and overall ordination (F = 1.702, P = 0.002). The species-environment correlation was strong (greater than 0.80 on all axes) and the selected EVs were responsible for most variation in macroinvertebrate distribution patterns (cumulative percentage variance of invertebrate data and invertebrate-environment relations = 58.9%). Bank exposure was strongly correlated with the first axis, while dissolved oxygen and Prionium were strongly correlated with the second axis.
Monte Carlo test of EVs considered individually (marginal effects) gave bank exposure, disturbance regime, indigenous bushes, Prionium, width, alien trees/canopy cover and dissolved oxygen as the most important variables in determining species distribution (Table 6). In addition, sedges (which included grasses) and flow were important when considered collectively (conditional effects). In addition sedges (which included grasses) were important when considered collectively (conditional effects).
Figure 3 shows the CCA ordination diagram based on family-level and species-level abundance. Important EVs were alien canopy cover, bank exposure, indigenous bushes and Prionium. A CCA was also performed on abundance of species across sites. With the exception of Aeshna sp. and Zygonyx sp., all Odonata species were associated with natural and cleared sites with an abundance of marginal vegetation that included indigenous bushes, grasses and sedges, and Prionium. These included Pseudagrion A and B, Trithemis A and B, Anax sp. and Paragomphus sp. Anax sp. and Pseudagrion A were associated mainly with natural sites, while Trithemis, Pseudagrion B and Paragomphus sp. were associated mainly with cleared sites with higher temperatures and an availability of pool biotopes. The ephemeropteran, P. bellum, and the trichopteran, Hydroptila, were also associated with these conditions, the latter species mainly in warm, cleared sites and the former in wide, natural sites. L. penicillata was in wide, deep natural sites with relatively strong flow rates and an abundance of indigenous bushes and sedges. A. harrisoni was associated with strong flow conditions. Euthraulus elegans, Aeshna sp., Cheumatopsyche sp. and Adenophlebia sp. were associated with high flow rates and alien trees, while A. bergensis B and C and Zygonyx sp. were associated with exposed banks, alien trees and sand and gravel substrates, in both cleared and alien sites.

CCA ordination diagram of environmental variables affecting the distribution patterns of the most common morphospecies across sites. Sites within each vegetation type are represented as centroids (N natural, C cleared and A alien)
The association of several species with marginal vegetation prompted SIMPER analysis of distribution patterns across dominant vegetation. Odonata species were the only ones to give characteristic assemblages that did not overlap across all three vegetation types. Pseudagrion A characterised sites with an abundance of indigenous vegetation and Prionium, while Aeshna sp. characterised sites with trees, whether alien or indigenous. Pseudagrion B characterised sites with high levels of Prionium and Orthetrum sp. characterised grassy sites. Paragomphus sp. and Trithemis A were common in sites with an abundance of grass, sedge and Prionium.
CCA performed on each disturbance regime in each river showed that different EVs were important in each river (Fig. 3; Table 7). Only suspended solids and percentage gravel were consistently important in cleared sites. Relatively few EVs were important in natural and alien sites, compared to a wide range of EVs in cleared sites, indicating that cleared sites varied greatly in EVs, with macroinvertebrates showing highly variable responses in cleared sites.
Discussion
In the Western Cape, each river and catchment has its own signature of benthic macroinvertebrate fauna (King and Schael 2001). This stems from a long period of geological and climatic stability, and the isolation of individual catchments, resulting in a unique assemblage of aquatic organisms. In particular, species with limited dispersal abilities (e.g. Plecoptera) have shown species and genetic differences between catchments (Wishart et al. 2003). These signature differences result in each river functioning in its own unique way and, as we show here also, responding to disturbance from alien vegetation in a different way. The result is that ecological data obtained from one river cannot necessarily be extrapolated to another, with each river having to be assessed individually to seek common trends.
Environmental responses to alien disturbance
Water quality variables that change through removal of riparian vegetation include temperature, light, fine sediments, salinity (measured as conductivity in this study), pH, dissolved oxygen, habitat quality and availability, allochthonous input of organic plant debris, shading and bank stability (King et al. 2003; Uys 2003). Here we found that alien disturbance affected water quality by lowering temperature, increasing oxygen and increasing suspended solids. The aliens here also affected the aquatic habitats by changing marginal vegetation and substrate composition. In turn, removal of alien Acacia trees resulted in greater exposure of the river and riverbanks to sunlight, encouraging growth of marginal grasses, sedges and Prionium reed. This reduced shading of the river also resulted in higher water temperatures in cleared sites than in natural ones. This is of particular concern in CFR rivers, as most endemic species are adapted to cold water conditions. Interestingly, adult Notonemouridae are barely affected by alien trees as they are adapted to total canopy cover, and are reduced by sudden and complete alien tree removal (Wishart et al. 2003).
Cleared sites are associated with sand and gravel substrate, high levels of suspended solids and high conductivity, all a result of erosion of exposed, alien-cleared banks (Maridet et al. 1996; Uys 2003; Vuori and Joensuu 1996). High silt levels are usually accompanied by low densities of benthic organisms and a reduction in many but not all species (Dallas and Day 1993; King et al. 2003; Rutherfurd et al. 2000).
Substrate composition is an important determinant of invertebrate distributions (Wright et al. 1984) and indeed we found here that sand and gravel substrates influenced assemblage patterns in cleared sites. For example, the relatively high number of certain Gomphidae in cleared sites was a response to higher availability of sandy substrates, their preferred habitats in the CFR (Samways and Wilmot 2003).
Linking SASS5 to disturbance by aliens
Low macroinvertebrate scores in alien sites were the result of low incidences of sensitive taxa, such as Teloganodidae, Sericostomatidae, Athericidae, Helodidae, Corydalidae and Aeshnidae, which are intolerant of shaded, alien-affected conditions. The mean ASPT was relatively high overall for alien vegetation in sites, indicating that where sensitive taxa are lost, they are not necessarily replaced by more tolerant taxa. Indeed, the incidence of tolerant taxa was lowest in alien sites (Table 2). This also suggests that water assessments using macroinvertebrate higher taxa, as in SASS5, are not ideal for monitoring riparian restoration projects in this endemic-rich region.
Cleared sites had scores that were consistently lower than alien scores, suggesting only marginal improvement in conditions or even deteriorating conditions after alien trees had been removed. The low incidence of high-scoring sensitive taxa in cleared sites suggests loss of these taxa as a result of impaired water quality conditions. Oxygen was an important determinant of assemblage patterns, and limiting for certain sensitive taxa in oxygen-poor cleared sites. High levels of suspended solids and sediment, as a result of erosion of cleared banks, may have compounded this effect. Those taxa affected included Sericostomatidae, Helodidae, Athericidae, Corydalidae and Aeshnidae. In contrast, these taxa, together with Heptageniidae, Teloganodidae and Philopotamidae, were responsible for the high scores in natural sites.
Where alien trees caused a loss of both sensitive and tolerant taxa, cleared sites saw a further loss of sensitive taxa, and their replacement with tolerant taxa, including most Coenagrionidae, Libellulidae, Hydroptilidae and Naucoridae. In turn, Corixidae, Notonectidae, Gerridae and Dytiscidae benefited from alien tree clearing.
In summary, the less sensitive, widespread taxa, including Hemiptera and Odonata, were probably responding to the increased availability of marginal vegetation in cleared sites. Eurytopic, vagile, widespread Odonata are known to be the first to colonise newly-created biotopes, such as dams (Steytler and Samways 1995). Temporally varying environments also favour generalists over specialists in relative fitness (Suhling et al. 2006), so that increased diurnal and seasonal fluctuations in temperature and insolation in exposed, cleared sites, favours more tolerant generalists. Rare, endemic or stenotopic species generally take longer to return, as shown by Odonata in the region (Samways and Sharratt 2010).
Abundance and richness of EPTO taxa
Alien sites had the lowest abundance and morphospecies richness. This agrees with other studies that found low diversities and richness in alien vegetation in terrestrial ecosystems (Samways et al. 1996; Donnelly and Giliomee 1985). Clearly, alien trees have a considerable effect on biodiversity.
Cleared sites had increased morphospecies richness and abundance, although both remained lower than in natural sites. Moreover, natural and cleared sites did not differ significantly in morphospecies richness and abundance, suggesting some measure of recovery. This was further supported by multivariate analysis of family-level abundance which also gave no significant difference between natural and cleared groups. However, there was a significant difference in morphospecies for the Rondegat River cleared and natural sites, but not the Molenaars cleared and natural sites, emphasising again that individual rivers have individual signatures. This may have been in part from the trajectory of indigenous vegetation recovery, as cleared sites along the Molenaars had good growth of indigenous Prionium, trees and shrubs, while Rondegat remained more exposed, with grass and sedges being prominent.
Between-river and seasonal effects largely obscured the effects of alien invasion and clearing on macroinvertebrate assemblage patterns. Different taxa dominated in each river and during each month, their responses to alien disturbance thus also differing. Each river and month therefore had to be analysed separately to seek consistent patterns. The Ephemeroptera and Odonata appeared to be most sensitive to alien disturbance, while the Plecoptera and Trichoptera showed variable responses.
Community responses to alien vegetation
Certain Ephemeroptera, Plecoptera, Trichoptera and Odonata species are excluded by alien trees, including sensitive and endemic taxa such as the teloganodid, L. penicillata, and the heptageniid, A. harrisoni, as well as a number of Baetidae, such as P. bellum, B. harrisoni, D. capensis and baetid C. Teloganodidae and Heptageniidae are grazers or scrapers which feed on periphyton (Barber-James and Lugo-Oritz 2003) and shading of alien sites prevents growth of their food source, affecting their populations.
Coenagrionidae and Hydroptilidae were absent from alien sites. Coenagrionidae use marginal vegetation as nursery and oviposition sites and are affected by absence of natural marginal vegetation under alien trees. The caenid, C. capensis, the hydropsychid, Cheumatopsyche sp. and the aeshnid, Aeshna sp., were not harmed or favoured by alien trees, suggesting they responded to factors common to alien and natural sites, such as oxygen levels or presence of riparian trees, whatever their origin, a situation that mirrors that on Mayotte Island (Samways 2003). In contrast, many Trichoptera, including the endemic and sensitive Glossosomatidae and Barbarochthonidae, favoured alien conditions. In particular, Leptoceridae in the A. bergensis group, together with Ecnomus and Macrostemum, were abundant in alien sites. The only Ephemeroptera that were abundant in association with alien vegetation were the leptophlebiids, Adenophlebia and Chloroterpes B, while Zygonyx sp. was the only odonate that increased with aliens. Notonemouridae also had a higher abundance and incidence in alien sites.
Resources that alien sites hold may include food or habitat availability, or water quality modifications (such as shading, lower temperatures and higher oxygen levels). Cummins et al. (1989) predicted that increased leaf litter would be followed by increased biomass of shredders. Shredders are known to be commonly associated with afforested and closed-canopied streams (Maridet et al. 1998; Weatherley et al. 1989). Greater availability of plant debris in alien sites may therefore have supported shredders, such as Notonemouridae, Leptoceridae and Barbarochthonidae.
Maridet et al. (1996) suggest that shaded, fast-flow conditions may be associated with an abundance of diatoms, an attractive food source for scrapers and generally associated with high flow and low light intensity environments (Campbell and Doeg 1989). Leptophlebiidae, which are scrapers or collector-gatherers (Barber-James and Lugo-Oritz 2003), occur in high velocity flows, with cobbled substrates under alien trees. Most case-building Trichoptera were unaffected by alien trees, suggesting that alien plant debris provided favourable material for case-building taxa, such as the A. bergensis group and Cheumatopsyche sp. It is likely that taxa unaffected by alien trees were responding to factors unrelated to alien disturbance, such as the cobbled substrate and faster flow rates under alien trees. Cobbled substrates are favoured by scrapers, such as Caenidae, Leptophlebiidae and Glossosomatidae, while Zygonyx is associated with principally riffles and rapids, irrespective of the bank vegetation.
Community responses to alien clearing
Some taxa increased in abundance and incidence when alien trees were removed. Odonata in particular increased, with only Aeshna sp. and Pseudagrion B not increasing in abundance after alien removal. Most odonates were most abundant in cleared sites, with the exception of Aeshna sp., Pseudagrion A and Anax sp., all of which were more abundant in natural sites. In addition to the Odonata, Hydroptilidae, Leptoceridae (A. bergensis C and L. helicotheca), Baetidae, Teloganodidae (L. penicillata) and Leptophlebiidae (E. elegans) increased where aliens were removed. Heptageniidae (A. harrisoni) also increased but only in March.
Alien removal exposes the river to increased sunlight, promoting macrophytes, algae and periphyton, with considerable consequences for food webs and faunal composition. Macrophytes are utilised by Leptoceridae (for food and shelter), hydroptilids (for shelter) and Odonata (for shelter and oviposition sites) (Bunn and Boon 1993; Maridet et al. 1998), which is why these taxa were mainly in cleared sites with an abundance of marginal vegetation.
Certain taxa may also have responded positively from water quality or habitat modification once alien trees were removed. Certain Leptoceridae responded to greater availability of grasses for case construction, while collector-gatherers such as Leptoceridae, Baetidae and Leptophlebiidae, increased with higher levels of suspended solids at cleared sites. Nevertheless, some taxa declined with clearing of alien trees. These were largely associated with tree canopies, whether alien or indigenous, such as Notonemouridae, Ecnomidae, the caenid, C. capensis, the hydropsychid, Cheumatopsyche sp., the aeshnid, Aeshna sp., the sericostomatid, Petroplax, and the leptocerid, Athripsodes C. Several other taxa, mainly Trichoptera, were also negatively affected. Zavaleta et al. (2001) pointed out that, while invasive plant species may provide usable food or habitat for indigenous fauna, rapid removal of these plant species, without restoration of indigenous vegetation, might leave this fauna without food or habitat. Sudden reduction in litterfall is also likely to affect energy flow, faunal composition and overall functioning of the river (Muotka and Laasonen 2002).
There was probably also the effect of sudden changes in water quality (such as increased temperatures and suspended solids or decreased oxygen levels), together with greater fluctuations in these variables and greater exposure to extreme values. Vuori and Joensuu (1996) found that logging of forests in Finland resulted in increased sand, silt, turbidity and suspended solids, causing a decline in shredders and an increase in filter feeders. This may explain the low abundance of Notonemouridae and Barbarochthonidae at cleared sites. Vuori and Joensuu (1996) also found that high silt loads in logged areas disrupted filter-feeding by Hydropsychidae, which may explain the low abundance of this group at cleared sites here.
Response of endemic taxa
Surprisingly, more narrow range endemic species were recorded at alien sites and cleared sites (ten species each) than at natural sites (eight species). However, abundance data gave a different picture. Recovery of endemic populations at cleared sites was limited, with abundance in cleared sites not matching those in natural sites. The sensitive sericostomatid, Petroplax, and the leptocerid, Athripsodes C, were reduced by alien invasion and again, to an even greater extent, by alien clearing. The sensitive teloganodid, L. penicillata, and the leptocerid, L. helicotheca, were harmed by alien invasion but recovered after clearing. The leptocerid, A. bergensis C, also increased with clearing. Glossosomatidae and Barbarochthonidae increased in association with aliens, yet were reduced by clearing. Cleared sites with short indigenous bushes and Prionium grouped with natural sites, while grassy sites were highly variable and grouped with alien sites. This suggests that recovery of sensitive, endemic taxa is delayed after clearing, and only occurs after recovery of indigenous Prionium and fynbos. Tolerant, widespread taxa therefore appear to be the first to recover, as with dragonflies in the region (Samways and Sharratt 2010). King and Schael (2001) noted that disturbed rivers lost their signature identities, perhaps through the loss of sensitive species. The highly variable dispersion of alien and cleared sites, compared with the more clustered dispersion of natural sites in the NMDS of species abundance supports this, with the ordination reflecting the more cosmopolitan and tolerant fauna of cleared and alien sites.
Environmental variables affecting faunal distribution patterns
Factors affecting benthic macroinvertebrates are numerous and interrelated (Wright et al. 1984), with a high degree of interaction, both directly and indirectly, between major variables, and with non-water-quality variables (e.g. substratum, current velocity, insolation) (Dallas and Day 1993), so that isolating the effect of a single variable is problematic. Between-river and seasonal factors dominated invertebrate responses in this study, as well as reducing the measured effect of specific EVs.
The Molenaars River was deeper and cooler with stronger flows and higher oxygen levels than the Rondegat River, resulting in physical factors explaining most between-river and seasonal differences (depth, temperature, conductivity), while vegetational factors (indigenous bushes, Prionium, alien trees, sedges and grasses and bank exposure), together with width and oxygen, mainly explaining distribution patterns within rivers and months. Oxygen probably interacted with vegetation type, so that cool, shaded alien sites had higher oxygen levels than warm, sunlit cleared sites.
Larval Odonata closely followed vegetation succession, as seen elsewhere (Osborn and Samways 1996), with marginal vegetation being particularly important (Rutt et al. 1989). Here, vegetation types were more important in determining invertebrate assemblages than was water chemistry or physical factors (with exception of oxygen and river width), supporting other results (Ormerod et al. 1987), although in Welsh streams, sites classified according to both marginal habitats and water chemistry (Rutt et al. 1989; Wright et al. 1984; Moss et al. 1987).
Seasonal effects on invertebrate abundance and distribution
Macroinvertebrate abundance was generally highest in autumn (March), although at variance with King’s (1983) findings where abundance was higher in summer (January), reasons for which are unclear. Weatherley et al. (1989) attributed higher autumnal abundance in Welsh streams to vertical migrations through the substrate to the surface, and that macroinvertebrate sizes large enough for capture were only attained by autumn.
Even moderate changes in water temperature affected growth rates and emergence times. Temperature probably interacts with day length to control emergence times and length of emergence periods (King 1983). Changes in temperature, as caused by alien disturbance, can cue life-history changes at inappropriate times (Rutherfurd et al. 2000). Warmer water in cleared areas may encourage faster growth and earlier eclosion. Under alien tree canopies, low temperatures and shade may reduce metabolic rates, delaying eclosion and increasing time taken to reach maturity. King (1981) found that ‘winter communities’ persisted for longer in upper river zones where temperatures were lower. Alien disturbance appears to have far-reaching effects on seasonal patterns of emergence, growth and survival, ultimately affecting faunal composition.
References
Allan JD, Flecker AS (1993) Biodiversity conservation in running waters. BioScience 43:32–43
Barber-James HM, Lugo-Oritz CR (2003) Ephemeroptera. In: de Moor IJ, Day, de Moor FC (eds) Freshwater invertebrates of Southern Africa, vol 7: Insecta I; Ephemeroptera, Odonata and Plecoptera. WRC report no. TT 207/03. Water Research Commission, Pretoria, pp 16–142
Bunn SE, Boon PI (1993) What sources of organic carbon drive food webs in billabongs? A study based on stable isotope analysis. Oecologia 96:85–94
Bunn SE, Davies PM, Mosisch TD (1999) Ecosystem measures of river health and their response to riparian and catchment degradation. Freshw Biol 41:333–345
Campbell IC, Doeg TJ (1989) Impact of timber harvesting and production on streams. Aust J Mar Fresh Res 40:519–539
Chessman BC (1995) Rapid assessment of rivers using macroinvertebrates: a procedure based on habitat-specific sampling, family level identification and a biotic index. Aust J Ecol 20:122–129
Chutter FM (1972) An empirical biotic index of the quality of water in South African streams and rivers. Water Res 6:19–30
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to statistical analysis and interpretation, 2nd edn. PRIMER-E Ltd, Plymouth
Cummins KW, Wilzbach MW, Gates DM, Perry JB, Taliaferro WB (1989) Shredders and riparian vegetations. BioScience 39:24–30
Dallas HF, Day DE (1993) The effect of water quality variables on riverine ecosystems: a review. WRC report no. TT 61/93. Water Research Commission, Pretoria
Dallas HF, Day JA (2007) Natural variation in macroinvertebrate assemblages and the development of a biological banding system for interpreting bioassessment data—a preliminary evaluation using data from upland sites in the south-western Cape, South Africa. Hydrobiol 575:231–244
Davies B, Day J (1998) Vanishing waters. University of Cape Town Press, Cape Town
Day JA, King JM (1995) Geographical patterns, and their origins, in the dominance of major ions in South African rivers. S Afr J Sci 91:299–306
Dickens CWS, Graham PM (2002) The South African Scoring System (SASS) version 5 rapid bioassessment system for rivers. Afr J Aquat Sci 27:1–10
Fischer DG, Hart SC, Schweitzer JC, Selmants PC, Whitham TG (2010) Soil nitrogen availability varies with plant genetics across diverse river drainages. Plant Soil. doi:10.1007/s11104-009-0260-2
King JM (1981) The distribution of invertebrate communities in a small South African river. Hydrobiol 83:43–65
King JM (1983) Abundance, biomass and diversity of benthic macro-invertebrates in a Western Cape river, South Africa. Trans R Soc S Afr 45:11–34
King JM, Schael DM (2001) Assessing the ecological relevance of a spatially-nested geomorphological hierarchy for river management. WRC report no. 754/1/01. Water Research Commission, Pretoria
King JM, Scheepers ACT, Fisher RC, Reinecke MK, Smith LB (2003) River rehabilitation: literature review, case studies and emerging principles. WRC report no. 1161/1/03. Water Research Commission, Pretoria
Kinvig RG, Samways MJ (2000) Conserving dragonflies (Odonata) along streams running through commercial forestry. Odonatologica 29:195–208
Lowe SR, Woodford DJ, Impson ND, Day JA (2008) The impact of invasive fish and invasive riparian plants on the invertebrate fauna of the Rondegat River, Cape Floristic Region, South Africa. Afr J Aquat Sci 33:51–62
Maridet L, Philippe M, Wasson JG, Mathieu J (1996) Spatial and temporal distribution of macroinvertebrates and trophic variables within the bed sediment of three streams differing by their morphology and riparian vegetation. Arch Hydrobiol 136:41–64
Maridet L, Wasson JG, Philippe M, Amoros C, Naiman RJ (1998) Trophic structure of three streams with contrasting riparian vegetation and geomorphology. Arch Hydrobiol 144:61–85
Moss D, Furse MT, Wright JF, Armitage PD (1987) The prediction of the macroinvertebrate fauna of unpolluted running-water sites in Great Britain using environmental data. Freshw Biol 17:41–52
Muotka T, Laasonen P (2002) Ecosystem recovery in restored headwater streams: the role of enhanced leaf retention. J Appl Ecol 39:145–156
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Ormerod SJ, Wade KR, Gee AS (1987) Macrofloral assemblages in upland Welsh streams in relation to acidity, and their importance to invertebrates. Freshw Biol 18:545–557
Osborn R, Samways MJ (1996) Determinants of adult dragonfly assemblage patterns at new ponds in South Africa. Odonatologica 25:49–58
Quinn JM, Cooper AB, Davies-Colley RJ, Rutherford JC, Williamson RB (1997) Land use effects on habitat, water quality, periphyton and benthic invertebrates in Waikato, New Zealand, hill-country streams. New Zeal J Mar Fresh Res 31:579–597
Richardson DM, van Wilgen BW (2004) Invasive alien plants in South Africa: how well do we understand the ecological impacts? S Afr J Sci 100:45–52
Rosenberg DM, Resh VH (eds) (1993) Freshwater biomonitoring and freshwater macroinvertebrates. Chapman & Hall, New York
Rutherfurd ID, Jerie K, Marsh N (2000) A rehabilitation manual for Australian streams, volumes 1 and 2. Cooperative Research Centre for Catchment Hydrology, and Land and Water Resources Research and development Corporation. Canberra, Australia. Accessible at www.rivers.gov.au
Rutt GP, Weatherley NS, Ormerod SJ (1989) Microhabitat availability in Welsh moorland and forest streams as a determinant of macroinvertebrate distribution. Freshw Biol 22:247–261
Samways MJ (2003) Threats to the tropical island dragonfly fauna (Odonata) of Mayotte, Comoro archipelago. Biodivers Conserv 12:1785–1792
Samways MJ, Sharratt NJ (2010) Recovery of endemic dragonflies after removal of invasive alien trees. Conserv Biol 24:267–277
Samways MJ, Wilmot B (2003) Odonata. In: de Moor IJ, Day JA, de Moor FC (eds) Freshwater invertebrates of Southern Africa volume 7: Insecta I; Ephemeroptera, Odonata and Plecoptera. WRC report no. TT 207/03. Water Research Commission, Pretoria, pp 16–142
Simaika JP, Samways MJ (2009) An easy-to-use index of ecological integrity for prioritizing freshwater sites and assessing habitat quality. Biodivers Conserv 18:1171–1185
Simaika JP, Samways MJ (2010) Comparative assessment of indices of freshwater habitat conditions. Ecol Indic. doi:10.1016/j.ecolind.2010.06.005
Smith J, Samways MJ, Taylor S (2007) Assessing riparian quality using two complementary sets of bioindicators. Biodivers Conserv 16:2695–2713
Steytler NS, Samways MJ (1995) Biotope selection by adult male dragonflies (Odonata) at an artificial lake created for insect conservation in South Africa. Biol Conserv 72:381–386
Suhling F, Sahlén G, Martens A, Marais E, Schutte C (2006) Dragonfly assemblages in arid tropical environments: a case study from western Namibia. Biodivers Conserv 15:311–332
Ter Braak CJF, Ŝmilauer P (2002) CANOCO reference manual and user’s guide to Canoco for windows: software for canonical community ordination (version 4.5). Microcomputer Power, Ithaca
Uys AC (2003) Development of river rehabilitation in Australia: lessons for South Africa. WRC report no. KV 144/03. Water Research Commission, Pretoria
Vuori K, Joensuu I (1996) Impact of forest drainage on the macroinvertebrates of a small boreal headwater stream: do buffer zones protect lotic biodiversity? Biol Conserv 77:87–95
Weatherley NS, Rutt GP, Ormerod SJ (1989) Densities of macroinvertebrates in upland Welsh streams of different acidity and land use. Arch Hydrobiol 115:417–431
Wishart MJ, Day JA (2001) Endemism in the freshwater fauna of the South-Western Cape, South Africa. Verh Internat Verein Theor Angew Limnol 28:1762–1766
Wishart MJ, Davies BR, Stewart BA, Hughes JM (2003) Examining catchments as functional units for the conservation of Riverine Biota and maintenance of biodiversity. WRC report no. 975/1/02. Water Research Commission, Pretoria, South Africa
Wright JF, Moss D, Armitage PD, Furse MT (1984) A preliminary classification of running-water sites in Great Britain based on macro-invertebrate species and the prediction of community type using environmental data. Freshw Biol 14:221–256
Zavaleta ES, Hobbs RJ, Mooney HA (2001) Viewing invasive species removal in a whole-ecosystem context. Trends Ecol Evol 16:454–459
Acknowledgments
This work was supported by the South African Working for Water Programme and the RUBICODE Coordination Action Project (Rationalising Biodiversity Conservation in Dynamic Ecosystems) funded under the Sixth Framework Programme of the European Commission (contract no. 036890), and by a Natural Sciences and Engineering Research Council of Canada (NSERC) PGS-D3 scholarship to John Simaika. We thank the anonymous referees for helpful and constructive comment which improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Samways, M.J., Sharratt, N.J. & Simaika, J.P. Effect of alien riparian vegetation and its removal on a highly endemic river macroinvertebrate community. Biol Invasions 13, 1305–1324 (2011). https://doi.org/10.1007/s10530-010-9891-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-010-9891-8