
Abstract
Seed dispersal is crucial for the success and spread of alien plants. Herbivores often establish a dual relationship with plants: antagonist, through herbivory, and mutualist, through seed dispersal. By consuming plants, herbivores may disperse large amounts of seeds, and can facilitate the spread of alien plants. However, seed dispersal of alien plants by herbivores has been largely uninvestigated. I studied factors associated with dispersal of alien and native seeds by the three most important vertebrate herbivores in SW Australia: emus (Dromaius novaehollandia), western grey kangaroos (Macropus fuliginosus) and European rabbits (Oryctolagus cuniculus). Overall frequencies of alien and native seeds dispersed by these herbivores were determined by differences among them in (1) the plant groups they predominantly disperse, that differed in frequencies of aliens versus natives, and (2) the predominant dispersal of aliens or natives within those plant groups. Emus and kangaroos (natives) tended to disperse predominantly alien seeds within plant groups (defined by life forms, dispersal syndromes, and diaspore size), whereas rabbits (alien) tended to disperse predominantly natives. This agrees with the hypothesis that herbivores will use predominantly plants that have evolved in different areas, because of less effective defences against new enemies. Overall frequencies were consistent with this pattern in kangaroos and rabbits, but not in emus. Kangaroos dispersed mostly plant groups that were mainly aliens (herbaceous species and small and medium sized dispersal units and seeds), which together with their predominant use of aliens over natives within groups resulted in the highest overall frequency of alien seeds (73%). Rabbits were similar to kangaroos in the type of plants dispersed, but their predominant use of natives over aliens within groups contributed to an overall predominance of native seeds in their pellets (88%). Emus dispersed mostly plant groups that were mainly natives (e.g. woody species with big diaspores), resulting in low overall frequency of alien seeds (11%), despite their predominant use of aliens over natives within plant groups. Thus, the within-groups trend pointed to a facilitative role of native herbivores of plant invasions through seed dispersal, but was obscured by the different use by herbivores of plant groups with different frequency of aliens.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
When arriving to a new area outside their native range, alien plants may leave their enemies and mutualists behind, but will also find new species with which they may establish new interactions. These interactions may be antagonistic (such as competition, predation and herbivory) or mutualistic (such as pollination and seed dispersal), and will determine both the biotic resistance and facilitation of the host community to the establishment and spread of new species (Levine et al. 2004; Richardson et al. 2000). Herbivores often establish a dual relationship with plants: while feeding on them (antagonists), they ingest large quantities of seeds and disperse a portion of them undamaged, thus acting as seed dispersers (mutualists) (Janzen 1984). Most research on the effects of herbivores on alien plant invasions has focused on their impacts through herbivory, whereas their facilitative effects through seed dispersal have been largely uninvestigated (Maron and Vilà 2001).
Seed dispersal determines to a large extent the ability of plants to colonize new areas and expand their ranges, and is critical to the success of alien plants (Richardson et al. 2000). Most research on seed dispersal has been centred on fleshy-fruited plants, chiefly shrubs or trees, often dispersed by frugivorous birds (Stiles 2000). The role of generalist herbivores has received less attention, despite their importance as seed dispersal agents, particularly for herbaceous species (Janzen 1984; Malo and Suárez 1995; Pakeman et al. 2002). In addition, herbivores, unlike frugivores, are not restricted to fleshy fruits, and may disperse seeds of a wide variety of species regardless of their adaptations for dispersal (e.g. seeds with adaptations for dispersal by wind, ants, water, or with no apparent morphological adaptations to dispersal, Calviño-Cancela et al. 2006, 2007, 2008) which, in some cases, may significantly increase the dispersal distances achieved by those plants (Higgins et al. 2003).
Some studies have highlighted the importance of the status of species (as alien or native) in determining the frequency and strength of interactions between them. Native herbivores are often expected to consume more native plants than aliens (alien plants thus gain enemy release in new areas, which favours their success: Enemy Release Hypothesis, Keane and Crawley 2002). This comes from a more general assumption of more frequent interactions between species that have evolved in the same areas. Accordingly, alien herbivores would consume predominantly alien plants, if they originate in the same areas. Through seed dispersal, this would lead to native herbivores dispersing native seeds more frequently than alien seeds. In contrast, alien herbivores would disperse alien seeds more frequently, which may favour their spread, creating a positive feedback among aliens that may facilitate invasions (“invasional meltdown”, Simberloff and Von Holle 1999).
The assumption of more frequent interactions between species that evolved in the same areas makes more sense for co-evolved specialists than for generalist species. Generalist herbivores are expected to view most plants as food (Bernays and Chapman 1994), regardless their origin, unless deterred by plant defences. In relation to this, it has been proposed that it is this lack of coevolution which would make plants more susceptible to new generalist enemies, against which they have not evolved effective defences (Parker et al. 2006). This would result in native herbivores feeding predominantly on alien plants, and alien herbivores on native plants (supported in a meta-analysis by Parker et al. 2006). Accordingly, through seed dispersal we would expect negative feedbacks between aliens (herbivores and plants) and natives (herbivores and plants), with native herbivores predominantly dispersing alien plants, which may favour their spread, and alien herbivores predominantly dispersing natives.
Real communities are often complex, and herbivores have a varied suite of plant groups to choose from. Behavioural and anatomical differences among herbivores lead to differences in the use of plant groups (e.g. woody vs. non-woody species, species with big vs. small fruits, etc.) (Cosyns and Hoffmann 2005; Davis et al. 2008). And these groups may differ in the abundance of alien species (e.g. aliens are often more frequently non-woody than woody), which may have an important influence in the relative frequency of aliens versus natives dispersed by herbivores.
I studied seed dispersal by the three most common generalist herbivores in an area where well preserved native shrublands come into contact with agricultural land in Western Australia. Agricultural land is the most important source of alien plants threatening native vegetation in this biodiversity hotspot (Dodd et al. 1993; Myers et al. 2000). The studied herbivores were emus, Dromaius novaehollandia L., western grey kangaroos, Macropus fuliginosus Desmarest, and European rabbits, Oryctolagus cuniculus L. Emus and kangaroos are natives in the area whereas rabbits are aliens. Native vegetation in the area is mostly woody whereas most alien plants are associated with agricultural land and are herbaceous. I analyzed differences among herbivores in the frequency of alien versus native seeds in their faeces and, with it, in their potential role in spreading alien plants, and discuss what causes these differences.
Materials and methods
Study site
The study was conducted in the Eneabba Plain (centred 275 km north of Perth, Western Australia, 29° 43′S, 115° 14′E). The climate is Mediterranean, mean annual rainfall is 510 mm, concentrated in winter, January and February are the hottest months (mean Tmax = 36°C, Tmin = 19–20°C) and June–August the coolest (mean Tmax = 19–21°C, Tmin = 9–10°C). The natural vegetation is kwongan (or heathland), a type of Mediterranean shrubland similar to the South African fynbos, that has exceptionally high species richness and endemism levels (Hopper and Gioia 2004).
Study herbivores
The focus herbivores in this study were selected for their potential ecological relevance, as they are the most abundant and widespread wild herbivores in SW Australia. All are declared pests of agriculture (Agriculture and Related Resources Protection Act 1976, Western Australian Department of Agriculture and Food).
In addition to their different status as alien or native, emus, western grey kangaroos and rabbits differ greatly in size, feeding habits, home ranges and thus seed dispersal potential (see Discussion for details). Emus feed on a great variety of plant material and are known to disperse large amounts of seeds (Davies 2002; Calviño-Cancela et al. 2006, 2008), although their effect on plant communities is still poorly known. Western grey kangaroos (kangaroos hereafter) are mostly grazers (Wann and Bell 1997). Despite the abundance of macropods in Australia, such as kangaroos, their role as seed dispersers has been mostly overlooked (but see Clifford and Drake 1985; Calviño-Cancela et al. 2008). European rabbits are considered the most ecologically damaging feral herbivore in Australia (Williams et al. 1995). Rabbits feed mostly on herbs and have been shown as important seed dispersers, most research being done in Europe and particularly in grasslands (e.g. Malo et al. 1995; Pakeman et al. 1999).
Sampling collection and processing
Faeces of emus, kangaroos and rabbits were collected in five sample dates (December 2005 and February, May, August and October 2006) in three sites of each type of habitat: (1) agricultural land (farms hereafter), (2) kwongan and (3) intermediate habitat (i.e. areas of kwongan 200 m away from farms). For farms, I collected faeces in 3-m-wide transects up to 2 km long alongside the edge of cultivated land and natural vegetation, in three farms separated 6 km on average. Farms were fenced, but the studied herbivores could enter and were often observed feeding inside farms. For the intermediate habitat, faeces were collected in transects in kwongan, 200 m away from the farm edges and running parallel to them. For kwongan, three sites were selected at least 2 km from farms, with an average distance of 6 km between sites.
Faeces where identified according to their shape and size. Faeces of emus and rabbits are unmistakeable, as there are no similar animals in the area. Regarding western grey kangaroos, there is another species of similar size in the region, the euro (Macropus robustus), which fecal pellets could be mistaken for those of western grey kangaroos, but they inhabit rock outcrops, and were not observed in the study area.
For emus, three fresh droppings (mean ca. 70 g per dropping) per site and sampling date were analyzed (nine per type of habitat per sampling date). For rabbits and kangaroos, ca. 150 g of fresh faecal material per site per sampling date was analyzed (450 g per habitat per sampling date). For rabbits, faeces were collected in latrines separated at least 50 m, with ca. 50 g of faecal material analyzed from each of three latrines per site per sampling date. For kangaroos, pellet groups, with ca. 5 to 15 faeces, were collected. I collected emu droppings and kangaroo’ pellet groups that were usually more that 5 m away from each other. Faecal samples were kept in paper bags, air dried until constant mass and weighted.
Faecal samples were broken up and visible seeds were collected and identified using reference collections. Apparently intact seeds were cut open and tested for viability by visual inspection of the embryo and endosperm (Gunn 2001). Only viable seeds were considered for the analyses. For Epacridaceae, the number of diaspores (i.e. seeds + endocarp, containing up to five seeds; endocarps hereafter) with at least one viable seed was counted. When the number of seeds (or endocarps in Epacridaceae) in a sample was higher than 15, the number of viable seeds was estimated from a subsample of a minimum of 10 individuals and up to 30 individuals when possible.
After collecting all visible seeds, the remaining material was set up for a germination trial to detect germinating seeds that had passed unnoticed by visual inspection. Because germination of many species in the study area occurs only after fire, the samples were divided in two halves: one half was treated with heat and smoke (Flematti et al. 2004) and the other was not treated. For the heat and smoke treatment, the faecal material was spread into an aluminium tray and saturated with boiled water (100°C). After cooling down, it was put into a plastic pot on top of a 1–2 cm layer of vermiculite and irrigated with diluted smoke water: 10 parts of water to one part of smoke water (Regen 2000 Smokemaster, Grayson, Melbourne, Australia). The non-treated portions were irrigated with water. In order to detect any potential seed rain into the greenhouse, controls with only vermiculite were also prepared (one for every ca. 10 samples). Faecal samples and controls were laid out randomly in the greenhouse and checked for seedling emergence every 3 days in the first 2 weeks, weekly in the first 2 months and more sparsely afterwards (usually fortnightly). Emerged seedlings were grown until identification was possible. The number of seedlings emerging from each sample was recorded, combining treated and non-treated halves.
Plant species were classified according to (1) status: native or alien, (2) life form: forbs (all herbs except graminoids), graminoids (grass-like species), woody (trees and shrubs) and parasitic vines, (3) dispersal syndrome, according to morphological attributes of diaspores: (a) unassisted, with no apparent structures that facilitate dispersal, (b) ant-dispersed, with elaiosomes, (c) endozoochorous, with fleshy structures, (d) exozoochorous, with structures that facilitate external adhesion to animals and (e) wind-dispersed, with structures that enhance air resistance, (4) seed size and (5) dispersal unit size.
Four size categories were used: small, medium, big and very big. Seed and dispersal unit length, width and thickness was measured with a calliper to the nearest 0.1 mm whenever possible, or taken from the literature. Size categories were assigned as follows: small: with length × width × thickness < 5 mm3; medium: 5–100 mm3; big: 100–1,000 mm3 and very big: >1,000 mm3. The sizes of seeds and dispersal units are subjected to different selective forces. Seed size is related to the probability of survival to the digestive process (Pakeman et al. 2002). Dispersal unit size limits the ability of the animal to handle, swallow and process it, depending on the size of its mouth and throat (Jordano 2000; Stiles 2000). In this study, dispersal unit size refers to the size of the item that is actually taken by the animal, i.e. a whole fruit or infructescence or a seed-containing fragment. Thus, a very big fruit (e.g. Carpobrotus modestus S.T.Blake) is classified as small, as the animals eat the fruit in small seed-containing fragments and, thus, the big size of the fruit does not represent any constraint for seed intake.
Statistical analyses
I performed a Multiple Correspondence Analysis (MCA) with STATISTICA 8.0 (StatSoft 2007) to examine the associations among plant traits and dispersers, reproducing the distances among them in a two-dimensional space. Since sizes of seeds and dispersal units were highly correlated (R = 0.834, P < 0.001, N = 95; Spearman rank correlation) I used only dispersal unit size, that provided better results. MCA codes data by creating several binary columns for each variable (one column per level); e.g. for dispersal syndrome, MCA creates five columns. For each column, plant species get a value of 1 if belonging to that category or otherwise get value 0. Since a plant species can only belong to one of those categories within each variable, this coding schema results in the addition of artificial columns. The inertia (variance) of the solution space is thus artificially inflated and the percentage of inertia explained by the first dimension is severely underestimated (Abdi and Valentin 2007). To correct this I applied the Benzécri formula (Benzécri 1979). Since axes in the MCA transformed space explain different amounts of inertia (variance), they were re-scaled for the distances in the graph to be representative of the true distances in the input space. Thus, Euclidean distances in the graph match the equivalent to Mahalanobis distances (Fukunaga 1990) in a PCA transform and can be visually assessed.
Likelihood-ratio tests (G tests, Sokal and Rohlf 1995) were used to assess the significance of the associations between pairs of variables. For life forms, parasitic vines (with only four species) were excluded.
To determine the factors that best predicted whether or not a species is dispersed by emus, kangaroos and rabbits, I used logistic models (formulated as Generalized Linear Models with binomial error distributions and logit link functions), with dispersal syndrome, alien status and life forms (as woody vs. non woody) as categorical variables, and dispersal unit size as continuous variable. I used best subset model building based on the model likelihood and selected the simplest models predicting plant dispersal by each herbivore.
As shown further in Results, herbivores differed in their use of plant functional groups, and plant groups differed in the frequency of native versus alien species. For this reason, in order to study the differences among herbivores in their dispersal of native versus alien plants, I first compared overall frequencies, and then frequencies within plant groups. The latter will allow us avoiding the confounding effect of the differential use by herbivores of plant groups with different proportion of natives versus aliens. The analyses within plant functional groups were only performed for those plant categories with enough alien and native species. This occurred with forbs, within life forms (there were only three native graminoids, N = 16 spp., and one alien woody species, N = 36 spp.), and with unassisted dispersal, within dispersal syndromes (ant and endozoochorous dispersal had only one and two aliens, N = 11 and 24 spp., and exozoochores had only two natives, N = 10 spp.). Very big seeds and dispersal units were removed from the analyses, as all were native, same as big seeds, with only three aliens (N = 14 spp.).
Results
Seeds from a total of 101 plant species were found in the faeces of emus, kangaroos and rabbits: 59 natives, 32 aliens and 10 unidentified species (Online resource 1). Kangaroos and rabbits were the most similar in regard to the plant species found in their pellets (abundance-based Jaccard index = 0.615), whereas emus and kangaroos were the least similar (0.360 for emus vs. kangaroos, and 0.532 for emus vs. rabbits). Emus showed the highest dispersal potential, with 4,018 propagules (viable seeds plus seedlings, seeds hereafter) per kg of faecal material, which is ca. 15 and 65 times the seed content per faecal mass in the pellets of rabbits (283 seeds/kg) and kangaroos (49 seeds/kg).
Associations among plant traits and herbivores
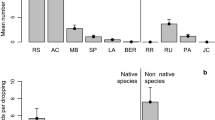
Herbivores differed in the plant types they dispersed; two big clusters of plant traits can be distinguished in Fig. 1 (see also Table 1), associated with dispersal by emus, on the left, and by kangaroos and rabbits, on the right. Herbivores differed in the proportion of species with different size of seeds (G6df = 47.1, P < 0.001) and dispersal units (G6df = 34.2, P < 0.001). Kangaroos and rabbits dispersed more species with small and medium-sized dispersal units and seeds, whereas emus dispersed higher frequencies of species with big and very big dispersal units and seeds. Herbivores differed also in the proportion of woody versus non-woody species (G2df = 8.2, P = 0.017): kangaroos and rabbits dispersed three times more non-woody than woody species, whereas emus dispersed similar quantities of woody and non-woody species.

Multiple correspondence analysis showing the associations among plant traits and dispersers. Projection on the first two dimensions, which explain 95% of the cumulative percentage of inertia. The axes were scaled according to the proportion of explained inertia for the distances in the graph to be representative of the true distances in the input space. The proportion of explained inertia were corrected with the Benzécri formula (Benzécri 1979). Unass: unassisted dispersal, Exoz and Endoz: exozoochorous and endozoochorous dispersal. Sm, Med, Big and Very big, refer to small, medium, big and very big dispersal units. Kang and Rab refer to kangaroo and rabbit, and Y and N to dispersal (Y) or not (N) by each animal
With main-effects only models, the factor that better predicted dispersal by the three herbivores was dispersal unit size, the only factor that was significant for all herbivores \( \left( {\chi_{1df}^{2} \ge 9.52,\;P < 0.005} \right), \) and a better predictor than seed size, which was not significant for rabbits. Status (as native vs. alien), considered alone, was only a good predictor of dispersal for kangaroos \( \left( {\chi_{1df}^{2} = 6.24,\;P = 0.01} \right), \) which dispersed more alien than native plants. For kangaroos, the simplest models that fit to the data best were the single effects models including the effect of dispersal unit size \( \left( {\chi_{1df}^{2} = 9.52,\;P < 0.002} \right) \) and including the effect of status \( \left( {\chi_{1df}^{2} = 6.63,\;P = 0.013} \right). \) For emus, the simplest models that fit the data best included the effect of dispersal unit size and dispersal syndrome \( \left( {\chi_{ 6df}^{2} = 30.33,\;P < 0.0001} \right) \) and the effect of the interaction between dispersal syndrome with status and dispersal syndrome with life form \( \left( {\chi_{ 5df}^{2} = 26.70,\;P < 0.0001} \right). \) For rabbits, the simplest best model was again that including the effect of dispersal unit size and dispersal syndrome \( \left( {\chi_{ 5df}^{2} = 23.58,\;P = 0.0003} \right), \) and the effect of life form and the interaction between status and dispersal syndrome \( \left( {\chi_{ 5df}^{2} = 21.91,\;P = 0.0006} \right). \)
Plant status (as native or alien) was associated with other plant traits: dispersal unit size (G3df = 18.93, P < 0.001), seed size (G3df = 8.86, P < 0.031), life form (G2df = 41.93, P < 0.001), and dispersal syndrome (G4df = 29.66, P < 0.001); aliens predominated among species with small seeds and dispersal units (52 and 57%), graminoids (81% were aliens), exozoochores (80%) and species with unassisted dispersal (59%). Natives predominated among species with big (81%) and very big (100%) dispersal units, woody species (97% were natives), and species dispersed by endozoochory (92%), ants (91%) and wind (77%). In addition, size of dispersal units and seeds was linked with life form (G6df = 56.94 and G6df = 61.39, P < 0.001): non-woody species had smaller seeds and dispersal units than woody plants.
Dispersal of native versus alien plants
Overall, emus dispersed significantly more native than alien plant species (G1df = 7.82, P = 0.005), whereas rabbits and kangaroos dispersed similar proportions of aliens and natives (G1df = 1.40 and 0.27, P > 0.2) (Fig. 2). Regarding the amount of seed dispersed, a better indicator of the strength of the interaction between plants and their disperser animals, emus and rabbits dispersed predominantly native seeds, whereas the opposite was true for kangaroos (G1df ≥ 63.1, P < 0.001, Fig. 2). The same general trends were observed in the three habitats (kwongan, intermediate and farms) for species and seeds (results not shown). As seen previously, herbivores differed in their use of plant functional groups, which differed in the frequencies of native versus alien species. For this reason, in addition to the overall frequencies of alien and native plants dispersed, I also analysed frequencies within plant groups, in order to test whether there was differential use of aliens or natives by the herbivores when removing the confounding effect of their differential use of plant groups with different proportion of natives versus aliens. As a general rule, emus and kangaroos (native herbivores) dispersed more alien plants than rabbits (alien herbivore) within plant groups. Differences were not significant for species frequencies but were significant for seed frequencies (G2df = 66.5, P < 0.001 for forbs, G2df = 93.0 and 90.0, P < 0.0001, for small diaspores and seeds, and G2df = 31.2, P < 0.001 for unassisted dispersal) (Fig. 2). Emus and kangaroos dispersed mostly alien seeds for forbs, unassisted species, and species with small diaspores and seeds, in contrast with rabbits, that dispersed lower proportions of aliens for those plant types (Fig. 2). For big dispersal units, kangaroos were removed from the analyses, as they dispersed only 4 species, and again emus dispersed more aliens than rabbits (Fig. 2), although this difference was not significant (G1df = 0.43, P = 0.512). The picture differed with medium sizes: both emus and rabbits dispersed mostly natives, in contrast with kangaroos (Fig. 2) (G2df = 164.8 and 163.5, P < 0.001, for medium diaspores and seeds).

Percentage of native (black) and alien (white) seeds dispersed by emus (E), western grey kangaroos (K) and European rabbits (R), overall and for each plant functional group with enough number of alien and native species. For the plant group “big dispersal units”, kangaroos were removed from the analysis, as they dispersed only four species. Results for seed size categories were very similar to those of dispersal unit sizes, thus only the later are shown. The numbers at the top of the bars are the sample sizes
Discussion
Within plant functional groups native herbivores (emus and kangaroos) tended to disperse higher percentages of alien seeds than rabbits. This agrees with the hypothesis of more frequent interactions between herbivores and plants that have evolved in different areas. Parker et al. (2006) explain this pattern by the lack of coevolution, making plants more susceptible to be consumed by new generalist enemies, against which they have not evolved effective defences. There was one exception to this pattern: Medium sized dispersal units and seeds. For this group, emus dispersed percentages of native seeds that were as high as those dispersed by rabbits. Additionally, this group made up an important proportion of all seeds dispersed, particularly for rabbits and emus, thus having a significant influence on the overall frequencies of aliens versus natives (Fig. 2). Most seeds in this group (>95%) were endozoochores, which were mostly natives (only 2 alien spp., N = 23) (Online resource 1). Endozoochores have fleshy nutritious fruits, which favours fruit consumption (and thus seed dispersal) by vertebrates. Kangaroos, like other herbivorous mammals, typically eat the fruits together with the foliage, whereas emus selectively consume fruits (pers. obs.). For this reason, the presence of effective deterrents in the leaves of native species might deter consumption by kangaroos but not by emus. Fruits are less likely to contain deterrents since they are adapted to encourage consumption by vertebrates. Thus the mechanism proposed in Parker et al. (2006), by which native herbivores would use predominantly alien species whereas alien herbivores would use predominantly native species, would not apply to endozoochous species. When removing endozoochores, the percentage of natives decreased for all herbivores in all plant groups, but in all cases rabbits dispersed the highest percentages of natives, as expected.
Despite this general trend within plant groups, the overall pattern (pooling together all plant species) did not follow this trend: emus, as rabbits, dispersed mostly native seeds. In addition to the differential use of alien versus native plants by alien versus native herbivores within plant groups, the overall pattern was also the result of their differential use of plant groups that differed in the frequencies of alien versus native species.
The type of plants dispersed by each animal depends on (1) what the animals ingest, determined by their feeding habits and palatability of plant species, and on constraints related to their ability to handle and swallow diaspores of differing sizes (Jordano 2000); and (2) the ability of propagules to survive the digestive process, which is determined by digestion effectiveness and size, shape and hardness of seeds (Janzen 1984). Size was indeed the most important factor determining the plant species dispersed by each herbivore. Emus were quite different in the types of species dispersed, whereas kangaroos and rabbits were rather similar (see also Davis et al. 2008). Emus are large birds with large mouth gapes and no teeth, and can handle and swallow large diaspores with no damage. The food is later processed in the gizzard, but the treatment there is probably milder for big seeds compared to the chewing apparatus of herbivorous mammals (Davies 2002). Kangaroos and rabbits have smaller mouth gapes and potent jaws, which explains the scarcity of big and very big dispersal units and seeds in their faeces. In addition, kangaroos and rabbits are grazers, feeding mostly on herb foliage (Wann and Bell 1997; Martin et al. 2007). Emus rely less on high fibre food, for their lower efficiency in digesting structural carbohydrates (Herd and Dawson 1984). Instead of large amounts of foliage, emus select relatively small quantities of conspicuous plant parts rich in nutrients, particularly fruits and seeds (Davies 2002). Thus, emus tended to disperse more woody species than kangaroos and rabbits. In addition, life form and size of diaspores and seeds were associated (this data set, Moles et al. 2005) and linked with other plant traits and, importantly, with alien versus native status. Thus, emus dispersed higher frequencies of woody species, which were linked with big and very big dispersal units and seeds, and dispersal by ants and wind, and all were linked with native status, which contributed to the high frequency of native seeds dispersed by emus, despite the predominance of aliens within plant groups in their droppings. Kangaroos tended to disperse herbaceous species and small and medium sized dispersal units and seeds. These traits were linked with exozoochorous and unassisted dispersal, and, more importantly, with alien status. Thus, kangaroos dispersed predominantly plant groups that were usually alien and, in addition disperse predominantly aliens over natives within groups, which resulted in the highest overall frequency of alien seeds among the three herbivores. For rabbits, despite being grazers like kangaroos, their predominant use of natives over aliens within groups together to their frequent use of endozoochores, that were mostly native, lead to an overall predominance of native seeds in their pellets.
The within groups trend of predominant use of alien plants by native herbivores and of native plants by alien herbivores (as in Parker et al. 2006) points to a role of native herbivores as facilitators of plant invasions through seed dispersal. However, in real communities, herbivores have a varied suite of plant groups to choose from, and this trend may be obscured by their different use of plant groups with different frequency of aliens, as it was the case in this study.
The assumption that alien herbivores will predominantly use native plants is valid only if alien plants and herbivores have overlapping native ranges. If alien plants come from different geographical areas, they are as novel to alien herbivores as native plants in the new place of arrival. In this study, 23 out of 32 alien species recorded shared their native areas with rabbits. The most common alien species in rabbit pellets was Arctotheca calendula. This plant is from South Africa, thus it is as novel for rabbits as Australian native plants.
Seed dispersal by herbivores can provide long travelling distances, which may be particularly important for species with otherwise limited dispersal capabilities, such as unassisted and ant-dispersed species (Calviño-Cancela et al. 2006, 2008). As in many other areas, most aliens in Western Australia are herbaceous (>80%, Peskett and Calviño-Cancela, unpublished data), for which dispersal by herbivores may play an important role (Janzen 1984; Pakeman et al. 2002), and most agricultural weeds are unassisted (≥70%, reviewed in Benvenuti 2007), for which herbivores can significantly increase dispersal ranges. Seed dispersal distances depend on how long seeds are retained inside the animal and how far the animal can move in that time. Emus have the largest dispersal potential, with seed retention times from 3 to 4 h to maximums of 100 days to many months for some very big seeds (Davies 2002), normal walking pace of 7 km/h, and average movements of 10–25 km/day (Davies 2002; Dawson et al. 1984), with records of more than 100 km in 15 days (Davies 2002). Western grey kangaroos use home ranges of 40–460 ha (Viggers and Hearn 2005). Mean retention time for solid markers in closely related eastern grey kangaroos is 32 h (Dellow 1982) to 39 h (Forbes and Tribe 1970). Rabbits are the most limited, with mean retention times of diaspores ca. 12 h (Cosyns et al. 2005) and home ranges of 0.15–10 ha (Stott 2003; Moseby et al. 2005), although regular movements of 400 m to their feeding areas have been observed in SW Australia (King et al. 1985).
As in many other regions, native vegetation in the study area is mostly woody; Alien plants come mostly from cultivated land and are mostly herbaceous. In this situation, the feeding habits of herbivores may become especially important in determining their potential role as dispersers of alien or native species. As a general rule, herbivores that are mostly grazers will tend to disperse more aliens, whereas those that use mainly woody species will tend to disperse more natives, regardless of their predominant use of aliens or natives within plant groups.
The benefits that alien plants obtain through enhanced seed dispersal by herbivores might be counteracted by negative effects through herbivory. However, although herbivory could potentially limit the abundance of certain species (see Maron and Vilà 2001 for a review), aliens in the study area come mostly from cultivated land, which can be considered as an inexhaustible source of aliens, and the impact of herbivores in reducing this supply can be considered negligible. In this situation, the effect of herbivores on alien plant invasions through seed dispersal is expected to overcome any possible negative effect through herbivory.
Efficient methods to limit the access of herbivores to cultivated land are recommended, as these would not only reduce damage to crops but also the spread of alien plants.
References
Abdi H, Valentin D (2007) Multiple correspondence analysis. In: Salkind N (ed), Encyclopedia of measurement and statistics, Sage
Benvenuti S (2007) Weed seed movement and dispersal strategies in the agricultural environment. Weed Biol Manag 7:141–157
Benzécri JP (1979) Sur le calcul des taux d’inertie dans l’analyse d’un questionnaire. Cahiers de l’Analyse des Données 4:377–378
Bernays EA, Chapman RF (1994) Host-plant selection by phytophagous insects. Chapman and Hall
Calviño-Cancela M, Dunn RR, van Etten E, Lamont BB (2006) Emus as non-standard seed dispersers and their potential for long-distance dispersal. Ecography 29:632–640
Calviño-Cancela M, Ayres Fernández C, Cordero Rivera A (2007) European pond turtles (Emys orbicularis) as alternative dispersers of “water-dispersed” waterlily (Nymphaea alba): effects on viability and germination. Écoscience 14:529–534
Calviño-Cancela M, He TH, Lamont BB (2008) Distribution of myrmecochorous species over the landscape and their potential long-distance dispersal by emus and kangaroos. Divers Distrib 14:11–17
Clifford HT, Drake WE (1985) Seed dispersal by kangaroos and their relatives. J Trop Ecol 1:373–374
Cosyns E, Hoffmann M (2005) Horse dung germinable seed content in relation to plant species abundance, diet composition and seed characteristics. Basic Appl Ecol 6:11–24
Cosyns E, Delporte A, Lens L, Hoffmann M (2005) Germination success of temperate grassland species after passage through ungulate and rabbit guts. J Ecol 93:353–361
Davies SJJF (2002) Ratites and tinamous. Bird families of the world, Oxford University Press
Davis NE, Coulson G, Forsyth DM (2008) Diets of native and introduced mammalian herbivores in shrub-encroached grassy woodland, south-eastern Australia. Wildlife Res 35:684–694
Dawson TJ, Read D, Russel EM, Herd RM (1984) Seasonal variation in daily activity patterns, water relations and diet of Emus. Emu 84:93–102
Dellow DW (1982) Studies on the nutrition of macropodine marsupials III. The flow of digesta through the stomach and intestine of macropodines and sheep. Aust J Zool 30:751–765
Dodd J, Martin RJ, Howes KM (1993) Management of agricultural weeds in Western Australia. Dep.of Agric, WA
Flematti GR, Ghisalberti EL, Dixon KW, Trengove RD (2004) A compound from smoke that promotes seed germination. Science 305:977
Forbes DK, Tribe DE (1970) The utilisation of roughages by sheep and kangaroos. Aust J Zool 18:247–256
Fukunaga K (1990) Statistical pattern recognition. Academic press
Gunn B (2001) Australian Tree Seed Centre Operations Manual. CSIRO
Herd RM, Dawson TJ (1984) Fibre digestion in the emu, Dromaius novaehollandie, a large bird with a simple gut and high rates of passage. Phys Zool 57:70–84
Higgins SI, Nathan R, Cain ML (2003) Are long-dispersal distance events in plants usually caused by non-standard means of dispersal? Ecology 84:1945–1956
Hopper S, Gioia P (2004) The southwest Australia floristic region: evolution and conservation of a global spot of biodiversity. Annu Rev Ecol Evol S 35:623–650
Janzen D (1984) Dispersal of small seeds by big herbivores: foliage is the fruit. Am Nat 123:338–353
Jordano P (2000) Fruits and frugivory. In: Fenner M (ed) Seeds: the ecology of regeneration in plant communities. CAB International, Wallingford, pp 125–165
Keane RM, Crawley MJ (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17:164–170
King DR, Wheeler SH, Oliver AJ (1985) The European rabbit flea, Spilopsyllus cuniculi, in southwestern Australia. Aust Wildlife Res 12:227–236
Levine JM, Adler PB, Yelenik SG (2004) A meta-analysis of biotic resistance to exotic plant invasions. Ecol Letters 7:975–989
Malo JE, Suárez F (1995) Herbivorous mammals as seed dispersers in a Mediterranean dehesa. Oecologia 104:246–255
Malo JE, Jiménez B, Suárez F (1995) Seed bank build-up in small disturbances in a Mediterranean pasture: the contribution of endozoochorous dispersal by rabbits. Ecography 18:73–82
Maron JL, Vilà M (2001) When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 95:361–373
Martin GR, Twigg LE, Zampichelli L (2007) Seasonal changes in the diet of the European rabbit (Oryctolagus cuniculus) from three different Mediterranean habitats in south-western Australia. Wildlife Res 34:25–42
Moles AT, Ackerly DD, Webb CO et al (2005) Factors that shape seed mass evolution. Proc Natl Acad Sci USA 102:10540–10544
Moseby KE, De Jong S, Munro N, Pieck A (2005) Home range, activity and habitat use of European rabbits (Oryctolagus cuniculus) in arid Australia: implications for control. Wildlife Res 32:305–311
Myers N et al (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Pakeman RJ, Engelen J, Attwood JP (1999) Rabbit endozoochory and seedbank build up in an acidic grassland. Plant Ecol 145:83–90
Pakeman RJ, Digneffe G, Small JL (2002) Ecological correlates of endozoochory by herbivores. Funct Ecol 16:296–304
Parker JD, Burkepile DE, Hay ME (2006) Opposing effects of native and exotic herbivores on plant invasions. Science 311:1459–1461
Richardson DM, Allsopp N, D’Antonio CM, Milton SJ, Rejmanek M (2000) Plant invasions—the role of mutualisms. Biol Rev 75:65–93
Simberloff D, Von Holle B (1999) Positive interactions of nonindigenous species: invasional meltdown? Biol Invasions 1:21–32
Sokal RR, Rohlf FJ (1995). Biometry. Freeman and Co
StatSoft, Inc. (2007) STATISTICA (data analysis software system), version 8.0. www.statsoft.com
Stiles EW (2000) Animals as seed dispersers. In: Fenner M (ed) Seeds: the ecology of regeneration in plant communities. CAB International, Wallingford, pp 111–124
Stott P (2003) Use of space by sympatric European hares (Lepus europaeus) and European rabbits (Oryctolagus cuniculus) in Australia. Mammal Biol 68:317–327
Viggers KL, Hearn JP (2005) The kangaroo conundrum: home range studies and implications for land management. J Appl Ecol 42:99–107
Wann JM, Bell DT (1997) Dietary preferences of the black-gloved wallaby (Macropus irma) and the western grey kangaroo (M. fuliginosus) in Whiteman Park, Perth, WA. J R Soc WA 80:55–62
Williams K, Parer I, Coman B, Burley J, Braysher M (1995) Managing vertebrate pests: rabbits. Bureau of Resource Sciences, Australia
Acknowledgments
I thank Byron Lamont for plant and seed identification and kind support during my stage in Curtin University (Perth, Australia), Ken Okamitsu for processing some of the samples, Peter Mioduszewski for assistance in the greenhouse, the Spanish Ministry of Education and Science for a postdoctoral fellowship in Australia and the Department of Environment and Conservation for permission to work in reserves. Plant identifications were confirmed with reference collections at the Western Australian Herbarium, Perth. During the preparation of the manuscript I was supported by a Isidro Parga Pondal contract (Xunta de Galicia). I thank Marc Cadotte and anonymous referees for their comments on previous versions of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Calvino-Cancela, M. Seed dispersal of alien and native plants by vertebrate herbivores. Biol Invasions 13, 895–904 (2011). https://doi.org/10.1007/s10530-010-9877-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-010-9877-6