
Abstract
Complex surveillance problems are common in biosecurity, such as prioritizing detection among multiple invasive species, specifying risk over a heterogeneous landscape, combining multiple sources of surveillance data, designing for specified power to detect, resource management, and collateral effects on the environment. Moreover, when designing for multiple target species, inherent biological differences among species result in different ecological models underpinning the individual surveillance systems for each. Species are likely to have different habitat requirements, different introduction mechanisms and locations, require different methods of detection, have different levels of detectability, and vary in rates of movement and spread. Often there is a further challenge of a lack of knowledge, literature, or data, for any number of the above problems. Even so, governments and industry need to proceed with surveillance programs which aim to detect incursions in order to meet environmental, social and political requirements. We present an approach taken to meet these challenges in one comprehensive and statistically powerful surveillance design for non-indigenous terrestrial vertebrates on Barrow Island, a high conservation nature reserve off the Western Australian coast. Here, the possibility of incursions is increased due to construction and expanding industry on the island. The design, which includes mammals, amphibians and reptiles, provides a complete surveillance program for most potential terrestrial vertebrate invaders. Individual surveillance systems were developed for various potential invaders, and then integrated into an overall surveillance system which meets the above challenges using a statistical model and expert elicitation. We discuss the ecological basis for the design, the flexibility of the surveillance scheme, how it meets the above challenges, design limitations, and how it can be updated as data are collected as a basis for adaptive management.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Non-indigenous species threaten ecosystems and biodiversity, with indigenous species severely impacted when non-indigenous species invade and establish (Case and Bolger 1991; Towns et al. 2006; Vitousek et al. 1997; Wanless et al. 2007). Island ecosystems are particularly vulnerable because species have often evolved in the absence of processes such as strong competition, herbivory or predation, that may result from the establishment of a non-indigenous species (see references within Courchamp et al. 2003). Furthermore, the absence of these processes allows introduced species to thrive. Invasive animals are considered a threat to biodiversity because they have the potential to cause community-level changes to ecosystems into which they invade and establish, through disequilibrium and changes to certain processes which leads to indigenous species declines and extinctions (Clark 1980; Dowding and Murphy 1994; Jones et al. 2008; Pickering and Norris 1996; Towns et al. 2006). They can also be agricultural pests and associated with the spread of human disease.
There is a global and local need to address invasive species (Hulme 2006; McKinney and Lockwood 1999), and to improve surveillance design for non-indigenous species (Broome 2007; Department of Conservation 2008; Russell et al. 2005). In addition to the environmental costs of invasion, the financial costs of control or eradication if an invasive species establishes can be high (Howald et al. 2007) and success is not always achieved (see examples in Myers et al. 2000). Therefore efforts to limit incursions (i.e. prevention) are generally preferred over eradication options (Myers et al. 2000), and investment in prevention should be increased (Leung et al. 2002). Early detection is key to reducing impacts, especially as many eradication programs (baiting and trapping) can negatively impact on indigenous species—although they may recover in the longer term. In the past, many invasive species, e.g. the black rat (Rattus rattus) and the house mouse (Mus musculus), have been able to invade extensively because surveillance measures were lacking, and responses to incursions have been reactive, occurring past the point of eradication. In the case of non-indigenous but native to the country species, surveillance is virtually non-existent.
The case study presented is Barrow Island, a 260 km2 island located 70 km off the west coast of Western Australia. It has recently come under the media spot-light due to the expanding oil industry on the island, and the potential impacts of this expansion on island flora and fauna. The Gorgon Project is a liquefied natural gas (LNG) development that will involve construction and operation of a gas plant on 300 ha of the 26,000 ha island. In particular, concern for the impacts of non-indigenous species has been demonstrated by the Western Australian Government, who imposed a condition to include a detection program with specified statistical power (“80 percent statistical power, or an appropriate alternative power”) for detecting non-indigenous species early enough to consider eradication without significant environmental consequences to native flora and fauna, with respect to any incursions that result from industrial activities (Government of Western Australia 2007). A condition of this nature on biosecurity surveillance design is unprecedented as far as we know in Australia. This presented certain surveillance design challenges, for which solutions are examined in this paper for vertebrate invaders.
Barrow Island has very high conservation value. On Barrow Island there are almost 400 species of plants, 13 terrestrial native mammal species, more than 110 bird species, 44 terrestrial reptile species, at least 1,261 species of terrestrial invertebrates, at least 59 taxa of subterranean fauna and 24 known endemic species. In 1908, Barrow Island was declared a Class A nature reserve for the protection of flora and fauna.
Barrow Island has maintained its Class A Nature Reserve status while being home to Australia’s largest onshore oilfield for over 40 years during which time more than 300 million barrels of oil have been produced. The introduction of exotic mammals has been prevented due to rigorous environmental management (Bamford Consulting Ecologists et al. 2005). Black rats (Rattus rattus) were present over a small part of the island in the 1970s and eradicated in 1991 (Morris 2002). House mice (Mus domesticus) have also been detected on the island in the past but were contained and eradicated before they could establish more widely (Morris 2002).
The design methodology was developed and described for an exemplar invertebrate, the big-headed ant (Pheidole megacephala) in an earlier paper (Barrett et al. 2009), and then applied to the black rat (Jarrad et al. in press). Here, we apply the design methodology further to cover a full suite of non-indigenous terrestrial vertebrate species, except birds, that potentially threaten Barrow Island. This was achieved by integrating the surveillance systems of multiple exemplar species into one overall comprehensive surveillance design, with the aim of practical implementation and reduced overall resource costs. We focus on ecological aspects of the design for vertebrates, as inherent biological differences among exemplar species result in different ecological models underpinning the individual surveillance systems for each. For example, they are likely to have different habitat requirements, introduction mechanisms and locations, require different methods of detection, have different levels of detectability, and vary in rates of movement and spread. However, the surveillance design methodology can be applied to all exemplar species with due consideration given to ecological differences. We also demonstrate the flexibility of the surveillance design and discuss how it can be updated as data are collected as a basis for adaptive management.
Materials and methods
Surveillance system based on one exemplar species
The surveillance system developed for individual exemplars on Barrow Island, described by Barrett et al. (2009) and Jarrad et al. (in press), is briefly summarized here.
Expert elicitation
For the individual surveillance designs, a group of six independent vertebrate specialists with local knowledge (the ‘experts’) were consultatively nominated to assist with expert judgment in the design and parameterization of the statistical model for the surveillance system (Fig. 1). The elicitation was held during two 1-day group workshops in late 2008, and followed the guidelines set out in Low Choy et al. (2009) where expert judgment provides information for the model, which can be updated as data become available. Group consensus judgments were sought in workshops such that the majority of opinion was captured, which is advantageous over other types of elicitation (see Burgman 2005; O’Hagan et al. 2006).

Process flow chart for determining the surveillance system. Surveillance system components (SSCs) consists of traps, surveys and incidental sighting, etc. Expert elicitation contributed at various stages of the model (asterisk) (Jarrad et al. in press)
Selection of ‘exemplar’ species
The potential non-indigenous species which may invade Barrow Island are virtually limitless. With no adequate techniques available with which to predict or prioritise invaders (Kolar and Lodge 2001), exemplar species were selected to provide a model that should cover the full range of possibilities. Here, we are concerned only with anthropogenic sources of introduction associated with the Gorgon Project (i.e. not with other sources, e.g. shipwrecks). The criteria for selection of an exemplar have previously been described (Barrett et al. 2009; Jarrad et al. in press) and included: threat of introduction to Barrow Island; known to be invasive; proximity to Barrow Island either on the Australian mainland or other overseas locations where materials may be sourced; has relatively well understood and reported biology and ecology; and, is able to be detected by a range of techniques applicable to other species. Importantly, the exemplar species were selected as representative of a range of other species. Birds were outside the scope of the study because they are mobile over long distances and discrimination regarding the mode of arrival would not be possible.
Based on the advice of experts, a set of six exemplar vertebrate species was selected: black rat, house mouse, mourning gecko (Lepidodactylus lugubris complex), little red tree frog (Litoria rubella), redback skink (Cryptoblepharus ustulatus), and pygmy python (Antaresia perthensis). The Asian house gecko (Hemidactylus frenatus) was also initially considered as an exemplar; however, after initially eliciting information for both geckos, the mourning gecko alone was selected since it is more difficult to detect (i.e. it is a non-vocal species), parthenogenic (Cuellar and Kluge 1974), and the same detection devices, entry points and habitat features applied to both.
Further details of the invasive biology for each of the exemplar species, including occurrence, biology, impacts, detection and eradication, are provided in Appendix 1 in ESM.
Risk mapping for targeting surveillance for individual non-indigenous species—introduction, habitat requirements and spread
In order to efficiently target surveillance, the risk of invasion was mapped across a large and heterogeneous landscape. The first stage involved stratifying the landscape spatially according to likelihood of entry and establishment which provides risk of invasion, such that numbers of surveillance system component (SSC) units can be calculated for each risk zone and locality (as described below). The second stage involves deployment of SSC units within zones and localities, and over time, using local fine scale spatial knowledge. The aim is to deploy the SSC units to maximize both power and cost both spatially within the zones and also temporally within a year. As discussed below, this relies on the skills and training of the operational staff carrying out the surveillance.
In the first stage, experts identified potential points of entry and habitat attributes for successful establishment for each exemplar. These were prioritised using relative importance weights (RIWs) for each entry point or habitat attribute, determined through eliciting pairwise comparisons and the Analytic Hierarchy Process (Saaty 1987). Pairwise comparisons were made using a linguistic scale ranging from equal preference (e.g. to both entry points in a pair) to absolute preference (e.g. of one establishment attribute over another), related to a numerical scale of 1–9. Entry points and habitat attributes for establishment were assigned to features that could be mapped using spatial layers available in a geographical information system (GIS), and cumulative RIWs were assigned to GIS polygons which resulted in maps of likelihood of entry and establishment. Likelihood of entry was multiplied with likelihood of establishment for each GIS polygon (via a GIS overlay), resulting in a map of invasion risk. Each entry point was extended to a larger area using a buffer distance which reflected the reported or elicited probable home range of each exemplar, as a measure of rate of movement and spread during the period immediately after arrival. The vegetation classes were based on Mattiske et al. (1993, 1997). GIS services were provided by Chevron Australia. Invasion risk was categorized into four levels (Table 1) using the combined distribution of entry and establishment RIWs. A detailed example of risk mapping for the black rat exemplar is provided by Jarrad et al. (in press).
Statistical model for detection of non-indigenous species
The statistical model underpinning the surveillance design has been previously described (Barrett et al. 2009; Jarrad et al. in press). The overall equation (Eq. 1) is given below and the conceptual model is shown in Fig. 1. In brief, detection is determined by the probability that the surveillance system will detect at least one individual of the target species, given that it is present in the sampling frame (F) when the population reaches a specified threshold (K) of independent individuals. The value of K is set to a number of individuals that is large enough to be detected (i.e. in the context of surveillance the detection of a single or very few invaders is often extremely difficult), but small enough to be effectively eradicated without significant environmental consequences and costs.
Each non-indigenous species has a risk (R j ) of invasion in risk zone j of the sampling frame, j = 1,…,4. Each of the i types of SSC i in the surveillance system has a different probability of detecting a non-indigenous species (σ i ) and cost of implementation (C i ). Assuming that each SSC i is independent, it contributes to a ‘utility’ (β i ) to the overall power (β). The number of SSCs of type i that should be deployed in zone j is given by the proportional areas of each zone.
Merger of individual SS into an integrated SS
For the surveillance system (SS) to detect more than one exemplar, the spatial relationships between the risk zones for each species needed to be established across the island. The difference in SSs designed to detect a single non-indigenous species versus multiple non-indigenous species lay primarily in the differences between their individual risk profiles across the target area. Invasion risk maps for each exemplar were integrated using a maximisation approach. For vertebrates, the risk zones for each exemplar largely overlap but are not identical in placement and/or magnitude. The risk zones for each exemplar species were overlaid using GIS, and for every point on the map, the maximum risk zone of any exemplar was adopted. If the risk zones do not occupy the exact space on the landscape but overlap (even if just to a small degree), the total area occupied by the overlapping risk zones is used when determining sampling frame. The new areas (i.e. sampling frame; F) were then used to recalculate an integrated surveillance system. Finally, the power of the SS to detect all vertebrate exemplars was recalculated.
This method was advantageous in terms of practicality of implementation, conservativeness, and simplicity. However, it is likely to result in a more costly surveillance system because the enlarged sampling frame increases the number of SSC units required; as a consequence the resultant power is much greater than 0.8. Overall, however, resource costs are reduced through integration compared with implementing surveillance plans for each exemplar species individually.
Utilising the total area of the overlapping risk zones to calculate F is appropriate for Zone 1. For Zone 2, the total overlapping area is computed and the area now encompassed by the integrated Zone 1 is subtracted. Any of the Zone 2 areas enclosed by the integration of Zone 1 are treated as Zone 1, and similarly for Zone 3; this is both mathematically and pragmatically acceptable. In the specific cases where an exemplar has risk of invasion equivalent to Zone 4 only, at a locality, then its risk of invasion remains at that level for that locality.
Results
Introduction, habitat requirements and spread for individual exemplars
Likely entry points for non-indigenous species of vertebrates were identified at three vessel and aircraft entry points on Barrow Island (a marine offloading facility, a barge landing site, and the airport) and five additional sites where human and cargo activity occurs (the LNG plant construction area, horizontal direct drilling site on the west coast, Gorgon construction village, existing oilfield construction camp, and existing oilfield base). Buffering around all entry points was set to distances of 100 m for rats (Dowding and Murphy 1994), geckos, skinks and snakes, and 50 m for mice and frogs, judged to be appropriate for Barrow Island by the experts.
Pairwise comparisons indicated that experts considered the marine offloading facility a relatively more important potential entry point than the remaining seven potential entry points—although there is variation among the actual RIW values depending on the exemplar species (Table 2). Expert judgments were largely based on amounts and frequencies of material, people, aircraft and vessels arriving at each locality.
Establishment attributes were identified (Table 3) and their relative importance judged by experts (Table 4). For example, experts considered human inhabited areas relatively more important than; vegetated and non-vegetated areas for mice. There are no sources of free-standing surface water on the island, except in the episodic cases of extreme rainfall associated with cyclonic depressions resulting in temporary surface water, which would otherwise be considered an important attribute for establishment in some exemplars.
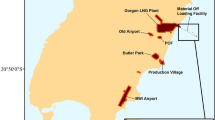
Risk maps were assembled for both entry and establishment using GIS. Risk of invasion was calculated as their product, and was categorsied using thresholds shown in Table 1. Risk is highest in Zone 1 and lowest in Zone 4. Areas of high and moderate invasion risk were observed at discrete localities on the island as a result of risk mapping. The lowest risk zone (Zone 4) has been excluded from the surveillance design because the risk of invasion is so small during the early stages of invasion that it would require an unrealistically large surveillance effort to contribute to the overall power of detection. This small risk of invasion and establishment in Zone 4 is nevertheless addressed by a more conventional ecological monitoring program on Barrow Island.
Based on consultation with experts and utilising an invasion probability tree to estimate and partition the likelihood of exemplar species occupying zones 1–3 during the early stages of invasion, the ratio of risk across integrated zones 1–3 was set to 55:40:5 for rats, mice and geckos, for frogs was set to 100:0:0 (spread to Zone 2 or 3 in the period immediately after arrival not likely), and for skinks 25:37.5:37.5 and snakes 55:45:0 (no Zone 3). Spread of risk across reflected behavior of the exemplar species and the likelihood of remaining near to points of entry (i.e. usually Zone 1).
Surveillance system design, components and deployment—methods of detection and levels of detectability
For the detection of non-indigenous species of terrestrial vertebrates on Barrow Island the surveillance system incorporates 14 detection methods (SSCs; Appendix 2 in ESM) deployed across the landscape according to entry and establishment risk (Fig. 2). The combination of these SSCs was considered by experts to provide the best overall strategy to detect the range of non-indigenous terrestrial vertebrates. For rodents, the benefits of using multiple detection methods has been identified (Dilks and Towns 2002; Russell et al. 2008a) recognising that usual methods used in eradication of populations (e.g. traps and bait stations) were not successful in detecting the presence of rodents when they were in small numbers.

Invasion risk map for vertebrates on Barrow Island. Invasion risk was calculated by multiplying likelihood of entry and establishment. Zone 1 (or High) indicates a higher risk compared to Zone 2 (Medium), etc. Areas of negligible risk (Zone 4) will be monitored as part of the long term ecological monitoring program on the island
Following consultation with experts, the surveillance system was designed with K selected to be a population size of 10 for each exemplar. In other applications or for another set of exemplars, the value of K may vary for individual species. It is noted that as K increases the total number of SSCs required decreases because it requires less units of SSCs to detect a larger number of individuals (Barrett et al. 2009; Jarrad et al. in press).
The surveillance system relies on inputs (i.e. detection footprints, detection probabilities, etc.) that have been supported by data in the literature where possible, but usually required expert judgments for data gaps (Appendix 2 in ESM). A description of the costs estimates indicate that structured biologist surveys have the highest cost per single performance to normal protocol for rats (3 h), and cameras have the lowest cost (15 min) (Table 5).
The resulting integrated surveillance system consists of a number of SSCs including Elliott traps, cage traps, barrier pitfall traps, ink pads, structured (or formal) surveys by biologists, chew cards, hair traps along tunnels, remote cameras, barrier funnel traps, Scentinel traps®, and incidental sightings by both biologists (n i. = 2,296, 920, 2,523, 3,261, 88, 54, 129, 322, 4,935, 474, and 3,022, across zones 1–3) and non-biologist staff (engaged workers and passive workers, n i1 = 5,068 and 5,044, respectively, in Zone 1).
The Gorgon construction workforce will be large and some will be trained in awareness of non-indigenous species and will contribute to the power of the surveillance effort (designated ‘engaged workers’). The majority of workers will not be trained (‘passive workers’) and therefore are assumed to have a low probability of detection and small detection footprint. The number of passive workers is high compared to the other SSCs, however, this is not problematic because there are likely to be around 2,500 passive workers on Barrow Island at any peak construction activities, and each worker observation period is limited to 3 weeks (one standard employee rotation). When the integrated SS (Table 6) is compared to the individual SSs (Appendix 2 in ESM), numbers of SSCs are larger, so the power to detect any exemplar is greater than the nominal 0.8 which each individual exemplar design was calculated to meet. Moreover, in the process of deployment, the numbers of SSC units were often increased above the power requirement. For example, one unit of ink pads was required in Zone 3, divided over six localities, each locality receiving 0.167 of a unit. These fractions were rounded up to whole units at each locality for practicality, such that six ink pads were specified in the surveillance system. Overall, the power of the integrated design was computed to be 0.9999.
Discussion
Inherent biological differences among exemplar species result in different ecological models underpinning the individual surveillance systems for each. By integrating these individual surveillance systems in a formal manner, the surveillance design methodology can be applied to all species with due consideration given to ecological differences. There were commonalities among exemplars in certain aspects of the surveillance design. For example, entry points were common to all exemplars. In particular, the MOF was considered the most important for all exemplars which is not surprising given it has the highest volume of material arriving and temporary storage. Volume of material and frequency of vessels arriving at locations were important considerations for introduction, as were modes of arrival such as through airport luggage, shipped food stores, etc. There was variation between exemplars in the importance of the remaining entry points. Certain habitat requirements were common among species, for example human inhabited areas were important for rats and mice, and vertical shelters were important for the skink, frog and gecko. These commonalties among species resulted in maps for individual species that had areas of Zone 1 and 2 centered around entry points, such that the integration of risk zones was logical. Although the integration resulted in larger areas of Zone 1 and 2 for most species when compared to areas for an individual species, it reduced resources compared with designing for each species individually.
The need for better non-indigenous vertebrate species surveillance has been advertised widely (Broome 2007; Department of Conservation 2008; Russell et al. 2005). Dealing with many unknown threats through developing surveillance systems for a range of non-indigenous exemplar species means that all exemplars are addressed, without the need for prediction of which ones would be most likely to incur or prioritization of non-indigenous species. This was an essential initial step to designing a surveillance system for all non-indigenous species of vertebrates, because predicting and prioritizing incursions is virtually impossible (Kolar and Lodge 2001) and would almost certainly be less efficient than using a system to detect all vertebrate incursions. Most of the exemplars were species typically associated with invasive characteristics, such as broad habitat requirements, wide dispersal or rapid reproduction. However, some species were not typically invasive, for example the little red tree frog; which is indigenous on the Australian mainland and has limited dispersal distances (pers com Paul Doughty, WA Museum). Nonetheless, it poses a threat, and all potential vertebrate threats were considered.
Another novel aspect of the surveillance design was incorporating many SSCs, including traditionally used traps and biological surveys, but also incidental sightings reported by workers on the island. Since there will be a large number of workers on the island, their collective numbers create a substantive power contribution that can be included using this design even if the vast majority are passive towards non-indigenous species and therefore have a low probability of detection and small detection footprint. The island workers (comparable to the public population in other applications) and biologist surveys are included to detect all exemplars, with the more cryptic species such as the python having a lower sighting probability (σ). For vertebrate species such as rats and mice, sightings of individuals and evidence of presence is likely to be obvious, and sighting of the little red tree frog is also judged to be likely in bathrooms and toilets.
The use of multiple SSCs has the further benefit of broadening the potential for detecting exemplified species. For this to be effective, the surveillance system relies on the similarities between the exemplar and the species it is designed to represent, such that the design for each would need to be very similar for the same level of power of detection to apply to both. Recent studies show greater success of detecting rat incursions (where in some cases they were deliberately released) on small islands by using a variety of detection methods (Russell et al. 2007, 2008a, b). Moreover, there were many SSCs that could potentially detect more than one of the exemplars, such as barrier pitfall traps which can detect the pygmy python, house mice and redback skink. The exemplars generally had different levels of detectability (σ) for devices that were able to detect more than one exemplar, such as ink pads.
There is a constraint in the integrated design, such that, if a trap is occupied by an exemplar then a less than the optimal number of traps will be free to detect other exemplars. However, if one species of rat is detected, then delimiting surveillance will be stepped up so all other species will be detected if present.
In the context of biosecurity surveillance design, power is defined as the ability of the system to detect a non-indigenous species if it is present. Emphasizing the statistical power of detection on a surveillance system is an enormous advance on current practices. In this application the focus is on power to detect, and an increase in the power of a surveillance system coincides with a decrease in the Type II error rate (the probability of falsely declaring the non-indigenous species to be absent). In the context of biosecurity surveillance design the probability of non-indigenous species presence going undetected is assumed to be far more likely and have greater consequence than mistaken or false detection (a Type I error), even though false detection may be inconvenient due to unnecessary response actions. There is a trade-off between Type I and Type II error rates, such that the lower the Type II error rate the higher the Type I error rate. This may not be the case on other contexts were the financial cost of response actions outweighs the cost of overlooking the non-indigenous species presence.
The power of the integrated design was computed to be much greater than the required 0.8. In light of the broad nature of the Ministerial requirement, a conservative approach was taken here which ensures that each exemplar species design would itself have power of at least 0.8. This might not be required in other surveillance situations. The methodology also enables design of integrated surveillance schemes to meet an overall power of 0.8, but this comes at a cost, in that the power to detect any individual exemplar may be less than 0.8.
From a management perspective, it is logical that statistical power requirements be placed on programs for surveillance, monitoring and eradication, that in practice generally do not incorporate power (e.g. Nielsen et al. 2009) but instead are heavily influenced by budget. In this application, whether the target species have been detected (observed or eradicated) can be estimated within certain bounds. Therefore protecting and conserving environmentally important areas can be done with statistical and practical confidence. It is inevitable that quantitative risk conditions such as the one placed on the Gorgon Project will become more common in future.
We also point out that actual numbers of incursions is likely to be very low given the extensive quarantine program, of which surveillance is only one component. Hence even if a power of 0.8 implies a 20% chance of non-detection due to the surveillance, the incursion is likely to be detected in other parts of the quarantine program and longer term ecological monitoring on the island. Moreover, the conservative nature of the integrated design gives an overall power of usually 0.9 or greater. Also, consideration must be given to the environmental damage and logistical impracticality of saturating an area with SSCs in an effort to achieve greater statistical power. In addition, many invaders may not survive due to the harsh climate on Barrow Island and Allee effects (Courchamp et al. 1999).
There is uncertainty associated with the expert judgments on model parameters, such as the probability of detection of a certain exemplar by a certain SSC, and how a non-indigenous species may behave on arrival which could influence measures of habitat suitability and spread. In addition, the different types of SSCs were limited for some exemplars—usually for the same exemplar where the biology and ecology were largely undocumented. Therefore, using expert elicitation was valuable in that it allowed us access to information that we could not have obtained in any other way in the short term. Experts provided information and insight into exemplar species biology and ecology when little or no published information was available, e.g. invasive impacts of the little red tree frog, redback skink and pygmy python. Moreover, the elicitation process built ownership by a stakeholder group. Although we are not currently able to quantify the uncertainty of expert elicitations, the next stage of our work examines the sensitivity of the model to the uncertainty in the elicitations. Moreover, the surveillance design is intended to be updated as new information and data become available, such that at each iteration of the surveillance the data collected will feed back into the design to refine the elicited parameter values. There is also uncertainty associated with the risk mapping, including mapping resolution, scale of the GIS layers available, and how well proxy GIS layers represent the variable of interest. These types of uncertainty are common to spatial modelling in general.
Non-indigenous reptile species are increasingly becoming wide-spread problems around the world. Examples include the brown tree snake on Guam (Fritts and Rodda 1998), cane toads in Australia (Catling et al. 1999), Burmese python in the United States (Rodda et al. 2009), Caribbean frogs on the Hawaiian Islands (Kraus and Campbell 2002) and various others listed in Pitt et al. (2005). The negative impacts of these species are increasingly documented and include threatening indigenous species, species extinctions and ecosystems disruptions. Despite the obvious threat and recent spread, there are few techniques developed to detect, monitor, control or eradicate these invasive reptiles and amphibians (Pitt et al. 2005; Usher 1989). Therefore, there is potentially substantial uncertainty regarding expert elicitations for SSCs. Invasive reptiles and amphibians may become more substantial problems in the future as their populations increase and expand, e.g. under scenarios of climate change (Kearney et al. 2008; Phillips et al. 2008).
This raises the question of why techniques for the detection (and eradication) are lacking for these groups of vertebrates. Many are cryptic and therefore possibly difficult to detect or perhaps they are not seen as a problem or priority for political, social, or economic reasons. For example they may not be agricultural pests and therefore not costly financially when present; there may be a lack of public awareness; they may not be present at the scale of other invasive species such as rats or mice; they may not have a history of spreading disease, or they may not have a direct impact on humans. Further research is recommended for detection and eradication of species in these groups. Even for widespread and otherwise well-known invasive species, such as the black rat, the lack of relevant data readily available for surveillance design was surprising.
With commencement of the Gorgon Project, the surveillance systems will be the first in Australia, and possibly world-wide, of its type. Ecologically based and statistically correct design methods help to ensure that operational programs are efficient, effective and consistent with stakeholder objectives. Programs designed without statistical input are at risk of not achieving their objectives, through failing to take some extra steps at the outset. This work can be incorporated into surveillance, monitoring and eradication programs more broadly.
References
Bamford Consulting Ecologists, Biota, RPS Bowman Bishaw Gorham (2005) Gorgon Development on Barrow Island: technical report: mammals and reptiles. Perth
Barrett S, Whittle P, Mengersen K, Stoklosa R (2009) Biosecurity threats: the design of surveillance systems, based on power and risk. J Environ Ecol Stat. doi: 10.1007/s10651-009-0113-4
Broome K (2007) Island biosecurity as a pest management tactic in New Zealand. In: Witmer GW, Pitt WC, Fagerstone KA, Clark CA (eds) Proceedings of an international symposium managing vertebrate invasive species. University of Nebraska, Lincoln, pp 104–107
Burgman M (2005) Risks and decisions for conservation and environmental management. Cambridge University Press, 502 pp
Case TJ, Bolger DT (1991) The role of introduced species in shaping the distribution and abundance of island reptiles. Evol Ecol 5:272–290
Catling PC, Hertog A, Burt RJ, Wombey JC, Forrester RI (1999) The short-term effects if cane toads (Bufo marinus) on native fauna in the Gulf Country of the Northern Territory. Wildl Res 26:161–185
Clark DB (1980) Population ecology of Rattus rattus across a desert-montane forest gradient in the Galapagos Islands. Ecology 61:1422–1433
Courchamp F, Clutton-Brock T, Grenfell B (1999) Inverse density dependence and the Allee effect. Trends Ecol Evol 14:405–410
Courchamp F, Chapuis J-L, Pascal M (2003) Mammal invaders on islands: impact, control and control impact. Biol Rev 78:347–383
Cuellar O, Kluge AG (1974) Natural parthenogenesis in the gekkonid lizard Lepidodactylus lugubris. J Genetics 61:14–26
Department of Conservation (2008) (Draft) Island biosecurity best practice manual. Department of Conservation, Wellington
Dilks P, Towns D (2002) Developing tools to detect and respond to rodent invasions of islands: workshop report and recommendations. In: Department of Conservation (ed) DOC Science Internal Series 59, Wellington
Dowding JE, Murphy EC (1994) Ecology of ship rats (Rattus rattus) in a kauri (Agathis australis) forest in Northland, New Zealand. N Z J Ecol 18:19–28
Fritts TH, Rodda GH (1998) The role of introduced species in the degradation of island ecosystems: a case history of Guam. Annu Rev Ecol Syst 29:113–140
Government of Western Australia (2007) Statement that a proposal may be implemented (pursuant to the Environmental Protection Act 1986). Gorgon Gas Development: Barrow Island Nature Reserve. Ministerial Environment Statement No. 748. State of Western Australia
Howald G, Donlan CJ, Galván JP, Russell JC, Parkes J, Samaniego A, Wang Y, Veitch CR, Genovesi P, Pascal M, Saunders A, Tershy B (2007) Invasive rodent eradication on islands. Conserv Biol 21:1258–1268
Hulme PE (2006) Beyond control: wider implications for the management of biological invasions. J Appl Ecol 43:835–847
Jarrad FC, Barrett S, Murray J, Parkes J, Stoklosa R, Mengersen K, Whittle P (in press) Improved design method for biosecurity surveillance and early detection of nonindigenous rats. N Z J Ecol 34. http://www.nzes.org.nz/nzje/new_issues/NZJEcol_JarradIP.pdf
Jones HP, Tershy BR, Zavaleta ES, Croll DA, Keitt BS, Finkelstein ME, Howald GR (2008) Severity of the effects of invasive rats on seabirds: a global review. Conserv Biol 22:16–26
Kearney M, Phillips BL, Tracy CR, Christian KA, Betts G, Porter WP (2008) Modelling species distributions without using species distributions: the cane toad in Australia under current and future climates. Ecography 31:423–434
Kolar CS, Lodge DM (2001) Progress in invasion biology: predicting invaders. Trends Ecol Evol 16:199–204
Kraus F, Campbell EW III (2002) Human-mediated escalation of a formerly eradicable problem: the invasion of the Caribbean frogs in the Hawaiian Islands. Biol Invasions 4:328–332
Leung B, Lodge DM, Finnoff D, Shogren J, Lewis M, Lamberti G (2002) An ounce of prevention or a pound of cure: bioecomonic risk analysis of invasive species. Proc R Soc Lond Ser B 269:2407–2413
Low Choy S, O’Leary R, Mengersen K (2009) Elicitation by design in ecology: using expert opinion to inform priors for Bayesian statistical models. Ecology 90:265–277
Mattiske EM et al. (1993) Flora and vegetation Barrow Island. Unpublished report prepared for Western Australian Petroleum Pty Ltd (WAPET), Perth
Mattiske Consulting Pty Ltd (1997) Assessment of revegetation on seismic, Barrow Island. Unpublished report prepared for Western Australian Petroleum Pty Ltd., Perth
McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol Evol 14:450–453
Morris KD (2002) The eradication of the black rat (Rattus rattus) on Barrow and adjacent islands off the north west coast of Western Australia. In: Veitch CR, Clout MN (eds) Turning the tide: the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, Cambridge, pp 219–225
Myers J, Simberloff D, Kuris AM, Carey JR (2000) Eradication revisited: dealing with exotic species. Trends Ecol Evol 15:316–320
Nielsen SE, Haughland DL, Bayne E, Schieck J (2009) Capacity of large-scale, long-term biodiversity monitoring programmes to detect trends in species prevalence. Biodivers Conserv 18:2961–2978
O’Hagan A, Buck CE, Daneshkhah A, Eiser JR, Garthwaite PH, Jenkinson DJ, Oakley JE, Rakow T (2006) Uncertain judgments: eliciting experts’ probabilities. Wiley, West Sussex
Phillips BL, Chipperfield JD, Kearney MR (2008) The toad ahead: challenges of modelling the range and spread of invasive species. Wildl Res 35:222–234
Pickering J, Norris CA (1996) New evidence concerning the extinction of the endemic murid Rattus maclaeri from Christmas Island, Indian Ocean. Aust Mammal 19:19–25
Pitt WC, Vice DS, Pitzler ME (2005) Challenges of invasive reptiles and amphibians. In: Nolte DL, Fagerstone KA (eds) Proceedings of the 11th wildlife damage management conference. Wildlife Damage Management working group, Traverse City
Rodda GH, Jarnevich CS, Reed RN (2009) What part of the US mainland are climatically suitable for invasive alien pythons spreading from Everglades National Park? Biol Invasions 11:241–252
Russell JC, Towns DR, Anderson SH, Clout MN (2005) Intercepting the first rat ashore. Nature 437:1107
Russell JC, Clout MN, Towns DR (2007) Early detection and eradication of invading rats. In: Witmer GW, Pitt WC, Fagerstone KA (eds) Managing vertebrate invasive species: proceedings of an international symposium. USDA/APHIS/WS, National Wildlife Research Center, Fort Collins, pp 268–272
Russell JC, Beaven BM, MacKay JWB, Towns DR, Clout MN (2008a) Testing island biosecurity systems for invasive rats. Wildl Res 35:215–221
Russell JC, Towns DR, Clout CR (2008b) Review of rat invasion biology: implications for island biosecurity. In: Department of Conservation (ed) DOC Science Internal Series 286, Wellington
Saaty TL (1987) Risk; its priority and probability: the analytic hierarchy process. Risk Anal 7:159–172
Towns DR, Atkinson IAE, Daugherty CH (2006) Have the harmful effects of introduced rats on island been exaggerated? Biol Invasions 8:863–891
Usher MB (1989) Ecological effects of controlling invasive terrestrial vertebrates. In: Drake JA (ed) Biological invasions: a global perspective, Scientific Committee on Problems of the Environment (SCOPE). Wiley, Chichester
Vitousek PM, D’Antonio CM, Loope LL, Rejmanek M, Westbrooks R (1997) Introduced species: a significant component of human-caused global change. N Z J Ecol 21:1–16
Wanless RM, Angel A, Cuthbert RJ, Hilton GM, Ryan PG (2007) Can predation by invasive mice drive seabird extinctions? Biol Lett 3:241–244
Acknowledgments
The authors would like to acknowledge the support of Chevron Australia Pty Ltd and the Australian Government’s Cooperative Research Centre’s Program. We thank the following vertebrate specialists for attending the elicitation workshops: Andrew Burbidge; Paul Doughty (Western Australian Museum); Ric How (Western Australian Museum); Keith Morris (Department of Environment and Conservation, WA); Dorian Moro (Chevron Australia Pty. Ltd.); John Parkes (Landcare Research, NZ); and Roy Teale (Biota Environmental Sciences Pty Ltd). GIS services were provided by Chevron Australia.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Jarrad, F.C., Barrett, S., Murray, J. et al. Ecological aspects of biosecurity surveillance design for the detection of multiple invasive animal species. Biol Invasions 13, 803–818 (2011). https://doi.org/10.1007/s10530-010-9870-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-010-9870-0