
Abstract
Resource availability and propagule supply are major factors influencing establishment and persistence of both native and invasive species. Increased soil nitrogen (N) availability and high propagule inputs contribute to the ability of annual invasive grasses to dominate disturbed ecosystems. Nitrogen reduction through carbon (C) additions can potentially immobilize soil N and reduce the competitiveness of annual invasive grasses. Native perennial species are more tolerant of resource limiting conditions and may benefit if N reduction decreases the competitive advantage of annual invaders and if sufficient propagules are available for their establishment. Bromus tectorum, an exotic annual grass in the sagebrush steppe of western North America, is rapidly displacing native plant species and causing widespread changes in ecosystem processes. We tested whether nitrogen reduction would negatively affect B. tectorum while creating an opportunity for establishment of native perennial species. A C source, sucrose, was added to the soil, and then plots were seeded with different densities of both B. tectorum (0, 150, 300, 600, and 1,200 viable seeds m−2) and native species (0, 150, 300, and 600 viable seeds m−2). Adding sucrose had short-term (1 year) negative effects on available nitrogen and B. tectorum density, biomass and seed numbers, but did not increase establishment of native species. Increasing propagule availability increased both B. tectorum and native species establishment. Effects of B. tectorum on native species were density dependent and native establishment increased as B. tectorum propagule availability decreased. Survival of native seedlings was low indicating that recruitment is governed by the seedling stage.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Understanding local and regional processes leading to plant invasions, especially mechanisms that control dominance of native vs. exotics species, is a major topic in plant ecology that has important implications for land management (Burke and Grime 1996; Tilman 1997; Davis et al. 2000; D’Antonio et al. 2001; Von Holle and Simberloff 2005; Chambers et al. 2007; Thomsen and D’Antonio 2007). Recruitment and species diversity are strongly influenced by availability of both resources and propagules (Foster and Tilman 2003; Foster and Dickson 2004). In functionally diverse communities, species tend to fully utilize available resources through spatial and/or temporal partitioning (Kemp 1983; Hooper 1998) and exhibit periodic recruitment from the propagule pool. However, disturbances that partially or totally remove resident vegetation can reduce competition and increase availability of resources leading to establishment of opportunistic native and exotic species (Burke and Grime 1996; Davis et al. 2000; Chambers et al. 2007).
A major factor in determining the ability of ruderal species, especially annual grasses, to dominate disturbed ecosystems is soil N availability (McLendon and Redente 1991; Young et al. 1999). Inorganic soil nitrogen can increase following removal of native perennial vegetation by grazing (Schuman et al. 1999), and especially immediately after fire (Neary et al. 1999; Rau et al. 2007). Annual species typically respond to soil nitrogen increases through higher growth rates and seed production (Monaco et al. 2003) which allows them to quickly occupy space, deplete resources and out compete perennial neighbors (Lowe et al. 2003). In contrast, perennial species often exhibit lower potential growth rates and reproductive output, but are capable of tolerating reduced resource levels which can represent an advantage in nutrient-poor environments (Chapin 1980; Aerts and van der Peijl 1993). Consequently, resource reductions have the potential to decrease population size or competitiveness of annual invasive species and provide opportunities for perennial plant establishment in invaded sites (Paschke et al. 2000; Blumenthal et al. 2003).
Invasion rates also are influenced by propagule supply of both natives and invaders (Tilman 1997; D’Antonio et al. 2001; Davis 2005; Von Holle and Simberloff 2005; DiVittorio et al. 2007). Determining the importance of resource availability vs. propagule supply is difficult because the two interact to influence plant community structure and resistance to invasion (Hamilton et al. 1999; Foster et al. 2004; Foster and Dickson 2004; Davis 2005; Thomsen and D’Antonio 2007). Resistance of native plant communities to addition of invader propagules is often higher when herbaceous perennial grasses and forbs exist in sufficient abundance to competitively suppress or exclude new colonizers (Tilman 1997; D’Antonio et al. 2001; Chambers et al. 2007). However, if propagule pressure is sufficiently high, periodic increases in resources like soil moisture or N can increase invasion rates (Davis et al. 2000; D’Antonio et al. 2001). Even relatively low numbers of invader propagules may be sufficient for establishment if severe or repeated disturbance reduces or eliminates resident species (D’Antonio et al. 2001).
In sagebrush-steppe of western North America, the exotic annual grass, Bromus tectorum L. (cheatgrass), is rapidly displacing native plant species and causing widespread changes in ecosystem processes that are negatively affecting native plant and animal populations (D’Antonio and Vitousek 1992; Knapp 1996). Before B. tectorum was introduced, sagebrush ecosystems were comprised mainly of shrubs and perennial bunchgrasses and exhibited fire return intervals of about 30–110 years (Whisenant 1990). Severe overgrazing by livestock following settlement (circa 1860) resulted in the decline of native herbaceous populations and expansion of B. tectorum (Knapp 1996). The invasive annual increased fine fuels (Whisenant 1990) and rate of fire spread (Link et al. 2006). An annual grass-fire cycle now exists with fire return intervals less than 10 years in much of the sagebrush steppe (Whisenant 1990).
The changes in sagebrush steppe can be explained largely by fluctuations in resources related to overgrazing and altered fire regimes, and to altered plant community composition and reduced input of native propagules (D’Antonio and Vitousek 1992; Chambers et al. 2007). Intact sagebrush-steppe vegetation with a high percentage of native perennial grasses is capable of sequestering resources and restricting B. tectorum establishment (Anderson and Inouye 2001; Booth et al. 2003; Beckstead and Auspurguer 2004) even after fire (Chambers et al. 2007). Removal of native perennial grasses increases soil resource availability, especially water and N, and favors B. tectorum establishment (Melgoza and Nowak 1991; Chambers et al. 2007). Because B. tectorum establishment, growth and reproduction decline with reduced N supply, N reduction should decrease biomass and seed production of the annual invader (Paschke et al. 2000; Monaco et al. 2003). Establishment of native perennial species, which are relatively more tolerant of low nutrient conditions (Monaco et al. 2003), should increase due to reduced competition from B. tectorum.
Changes in abundance of native species has influenced availability of propagules (seed rain and seed bank) and, consequently, the capacity for native species establishment. Sagebrush-steppe seed banks often are dominated by annual grasses, forbs, and the shrub Artemisia tridentata, but maintain a perennial herbaceous component (Koniak and Everett 1982; Wehking 2002). Following fire and conversion to B. tectorum dominance, seed banks of native species, especially perennials, decline precipitously (Humphrey and Schupp 2001). Prolific seed production of B. tectorum (10,000 or more seeds m−2) ensures high propagule pressure of the invader (Young et al. 1969; Humphrey and Schupp 2001). Because recruitment of native perennials is limited by seed availability, seed addition should increase the probability of establishment.
Little is known about the relative importance of resource vs. propagule availability in determining establishment of native perennial species in sagebrush-steppe dominated by B. tectorum. We conducted a mechanistic experiment to evaluate the interacting effects of resource and propagule availability on establishment of B. tectorum and a functionally diverse mixture of native perennial species. Resource availability was manipulated using sucrose addition to promote microbial immobilization of soil N. Propagule availability was manipulated by seeding different densities of both B. tectorum and a functionally diverse mixture of native perennial herbaceous and shrub species. We asked three questions: (1) How does resource availability (inorganic N) influence establishment of B. tectorum and a functionally diverse mixture of native perennial species? (2) How does propagule availability of native perennial species and B. tectorum influence establishment of B. tectorum vs. native perennial species? (3) How do resource availability and propagule availability interact to determine establishment and persistence of B. tectorum and native perennial species?
Materials and methods
Study area
The study was conducted near Winnemucca, Nevada, USA (lat 41°12′N, long 117°23′W; elevation ~1,524 m), in the Great Basin sagebrush steppe of western N.A. Historically, native vegetation at the site was representative of a Wyoming big sagebrush ecological type (Artemisia tridentata Nutt. ssp. wyomingensis Beetle and Young/Poa secunda J. Presl (West and Young 2000). Currently, vegetation is dominated by B. tectorum L. (cheatgrass) and introduced annual forbs such as Draba verna L. and Sisymbrium altissimum L. Conversion to annuals likely occurred after an extensive wildfire in summer 1999. Mean annual temperature is 9.5°C, with average minimum temperatures of −8.2°C in January and maximum mean temperatures of 33.1°C in July (Western Research Climate Center 2007). Annual precipitation is about 300 mm. Long-term average precipitation for the water year (October–September) at the nearest climate station (lat 41°50′N, long 117°55′W; elevation ~1,390 m) is 238 mm, and average precipitation was 102, 118, and 150% for study years 2004, 2005 and 2006, respectively. Soils are coarse-loamy, mixed, superactive, mesic Xeric Petrocambids (Denny 2002). The study area (~25 ha) was grazed by livestock from the late 1800s until fall 2002 when the study site was fenced.
Experimental design
The experiment was a randomized, split plot design implemented in 2003 and 2004 at separate locations within the study area. To examine the effect of reducing available N on B. tectorum and native perennial species, six main-plots (23.0 × 15.0 m each) were established in both the 2003 and 2004 growing seasons (n = 12). Each year, three randomly selected main-plots received sucrose application to reduce available N (rate equivalent to 150 g C·m−2) and three were left untreated (control). The sucrose rate was similar to that used in other nitrogen reductions studies in semi-arid ecosystems (Paschke et al. 2000). Each main-plot had 20 split-plots that were seeded with a factorial combination of a native perennial species mixture at one of four densities (0, 150, 300, and 600 viable seeds m−2, with equal numbers of all six species) and B. tectorum at one of five densities (0, 150, 300, 600, and 1,200 viable seeds m−2). The native perennial seed mixture included the shrub A. tridentata ssp. wyomingensis, forbs Achillea lanulosa Nutt and Sphaeralcea grossularifolia (Hook and Arn.) Rybd., and grasses P. secunda, Elymus multisetus M.E. Jones and Pseudoroegneria spicata (Pursh) A. Löve. The species used in the seed mixture are members of the regional species pool that represent a range of life histories, functional groups and rooting patterns. The highest level of B. tectorum density was included to more closely reflect the high seed levels in the seedbanks of this species (Humphrey and Schupp 2001). Individual split-plots were 1.5 × 2.5 m each and were separated by 2.0 m buffer strips. Split-plots were seeded once and monitored over two growing seasons. The number of growing seasons after seeding (a repeated measure) was treated as a split-split-plot factor. Each iteration of the experiment was used as the blocking factor and treated as a random effect to account for annual variation and seeding location. Thus, when all data from both iterations of the study were included in a single analysis, the experiment could be considered a completely randomized block design with a split-split plot treatment structure.
To reduce density, biomass and seed rain of B. tectorum and other annuals, the herbicide glyphosate was applied as a one-time treatment after emergence but prior to flowering the spring before each set of treatments was implemented. Seeding occurred in fall (Late October), and the same experimental protocols were repeated in 2003 and 2004. Seeds of B. tectorum and A. tridentata were collected locally, cleaned and stored in paper bags under cool conditions (~4°C) until use. Seeds of the remaining perennial species were provided by the USDA-NRCS Aberdeen Plant Materials Center (Aberdeen, ID). Seed purity and viability were determined (Association of Official Seed Analysts 2000) prior to calculating seeding rates based on pure live seed.
After herbicide application and prior to seeding, we permanently marked all plots and manually removed litter, weeds, bunchgrasses and other conspicuous perennial species. The soil surface was raked to prepare the seedbed, and then the appropriate seed combination (mixed with 80 g of rice hulls to facilitate even distribution) and half of the total sucrose amount was hand-broadcast over each split-plot. To incorporate seeds and sucrose into the soil surface and to reduce seed loss, the soil surface was packed using a manual roller-packer and covered with a biodegradable jute mesh (~2.5 cm opening). The remaining half of the sucrose was broadcast at the beginning of the following spring (March). A permanent 1.0 m2 quadrat and nested 0.1 m2 quadrat were established within each individual seeded split-plot to assess, respectively, native species and B. tectorum densities. During the first and second growing seasons after seeding, all non-seeded species were periodically removed from the plots.
Soil sampling and analyses
To assess sucrose effects on soil available N and Phosphorus (P), we placed two mixed-bed ion-exchange resin capsules at 15 cm depth in a subset of split plots seeded in 2003. Resin capsules are mesh-covered spheres, about 2.7 cm in diameter, that contain both high-capacity anion and cation exchange resins (Unibest Inc, Bozeman, MT). Resin technology integrates nutrient availability during the period of time the capsules are in the soil (Yang et al. 1991). The selected split plots corresponded to four B. tectorum/native species mixture seeding combinations (0/0, 300/0, 0/300 and 300/300 seeds·m−2). Resin capsule measurements were conducted during six consecutive time periods (~6 months each): (1) October 2003–April 2004, (2) April 2004–October 2004, (3) October 2004–April 2005, (4) April 2005–October 2005, (5) October 2005–May 2006 and (6) May 2006–October 2006. In general, the sampling period from October to April (May) was representative of fall through winter months, whereas the April (May) to October period was representative of spring through summer months (period of vegetation growth). Resin capsules were exchanged after each measurement period.
In the laboratory, resin capsules were washed thoroughly with deionized water and dried overnight at 60°C. To quantify sorbed N and P anions, capsules were placed in 50 mL polypropylene tubes to which 40 mL of 1 N HCl was added. Tubes were shaken for 1 h, then centrifuged and the clear liquid decanted. Quantification of orthophosphate (vanomolybdate chemistry) and nitrate ions (NO3 −) in decanted liquid was done simultaneously using a Lachat flow-injection system. Blank values for ammonium (NH4 +) were similar, or even slightly higher, than those observed in the field samples. High NH4 + values in the blanks likely resulted from NH4 + release from amides in the resins during HCl extraction (R. Blank, pers. obs.). As a consequence, subtracting the field values for NH4 + from the blank values often resulted in NH4 + values that were less than zero. Thus, NH4 + was excluded from statistical analyses. To make data comparable for NO3 − values were divided by days that resin capsules were in the soil and expressed as μ moles of sorbed anion·day−1.
Vegetation sampling
Individual split-plots seeded in fall 2003 were censused in June 2004, 2005 and 2006. Split-plots seeded in fall 2004 were censused in June 2005 and 2006. We considered the time when B. tectorum reached maturity (late June) as end of growing season. To assess establishment of native species, we recorded density of perennial individuals that were alive at each census date in 1.0 m2 permanent quadrats that were located in the center of the lower half of each split-plot. These plots were used only to census plants during the study. For B. tectorum, we counted number of plants in the 0.1 m2 nested quadrat. We also collected 15 randomly chosen B. tectorum plants per split plot to assess aboveground biomass, seed biomass and seed number each growing season after seeding. Both seeds and plants were placed in a paper bag and transported to the lab. Seeds were separated from herbaceous biomass and the herbaceous component was oven-dried (60oC) to a constant weight. Seeds were air-dried on a lab bench. We recorded total aboveground, herbaceous and seed biomass, and number and weight of filled seeds. During fall, B. tectorum seeds were returned to the split-plot from which they were harvested and broadcast onto the plot surface to ensure that seed banks were not influenced by sampling.
To assess treatment effects on growth of B. tectorum and native perennial species during the first growing season after seeding, we measured height and diameter (basal diameter for grasses, mean canopy diameter for shrubs and forbs) of 15 plants for B. tectorum and 1–15 plants, depending on availability in a given treatment, for each native seeded species. For grasses, we recorded number of tillers of each individual plant. These measurements were labor intensive and were conducted for all seeded species only in 2004 for plots seeded in 2003. We repeated these measurements in June 2006 in the same set of plots to assess status of individuals that survived the third growing season.
Statistical analyses
Statistical analyses were conducted using SAS 9.1 software (SAS Institute Inc 2002). Box Cox transformations were used to meet analysis of variance (ANOVA) assumptions of normality and equal variance for all data. In general, data were analyzed using a mixed effects ANOVA with a split-plot structure in SAS PROC MIXED. For soil NO3 − and orthophosphate availability, we averaged daily amounts of μ moles of NO3 − and ortho-P sorbed to the resins across seeding treatments and sampling period. Sucrose treatment was the main-plot factor and sampling period was the split-plot factor. Native species density, B. tectorum density, biomass and number of seeds per plant, and B. tectorum biomass m−2 and number of seeds m−2 were analyzed with sucrose as the main-plot factor, the factorial combination of four (native mixture) × five (B. tectorum) seeding densities as the split-plot factor, and number of growing seasons after seeding as the split-split-plot factor. Analyses of plant growth variables (height, diameter and tillering) for B. tectorum and native perennial species were based on mean values obtained from plants collected in each individual split-plot for each species. Sucrose was the main-plot factor, and the factorial combination of four (native mixture) × five (B. tectorum) seeding densities was the split-plot factor.
For significant factors and interactions, least squares means were compared using the Tukey–Kramer test. All tests were performed at the 0.05 significance level, and all means are presented as untransformed values ± standard error (SE).
Results
Soil data
Sucrose application decreased soil resin available NO3 −, but the effect varied over time (sucrose x sampling period: F 5,86 = 6.0, P = < 0.0001). A 69% decrease in nitrate availability occurred for the sampling period when sucrose was applied (October 2003–April 2004), but no differences in control and sucrose-amended plots were detected thereafter (Fig. 1a). In control plots, the highest NO3 − availability occurred during the first sampling period. Also, NO3 − in control plots was higher in mid-fall through early spring than in mid-spring through early fall during the first 2 years (Fig. 1a). Soil NO3 − concentrations in sucrose-amended plots followed a pattern similar to control plots, although differences among sampling periods were not significant. Soil orthophosphate availability showed no effects of sucrose addition, but like NO3 − was highest during the first sampling period (sucrose × sampling period: F 5,86 = 35.4, P = < 0.0001; Fig. 1b).

a Soil available NO3− (μmol·day−1) and b soil available ortho-P (μmol day−1) in control (no sucrose) and sucrose addition plots for the six sample periods. Asterisk indicates significant differences among sucrose treatments for a given sampling period (P < 0.05). Values are mean ± 1 SE
B. tectorum and native species plant density
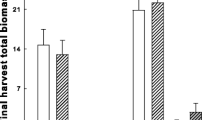
Over the two growing seasons of the experiment, there were 31% less B. tectorum plants in sucrose-addition plots (926 ± 90 plants m−2) than in control plots (1,347 ± 119 plants m−2) (sucrose: F 1,12 = 23.1, P = 0.0004). Bromus tectorum density increased with increasing seeding rate (B. tectorum seeding density: F 3,181 = 28.8, P = < 0 .0001; Fig. 2), but was not affected by native species seeding densities. The density of B. tectorum also increased over time (year: F 1,192 = 988.8, P = < 0.0001; Fig. 2) from 270 ± 17 plants m−2 in the first growing season to 1,996 ± 120 plants m−2 in the second season.

Mean (±1 SE) number of B. tectorum plants for the different B. tectorum seeding densities (150, 300, 600 and 1,200 seeds m−2) at the end of the first and second growth years after seeding. Different lowercase letters indicate significant differences across seeding densities and growth years (P < 0.05)
Density of native perennial seedlings was unaffected by sucrose addition over all levels of native species seed addition. The number of native seedlings did not differ between control and sucrose plots in the presence of B. tectorum, but was lower in sucrose plots than in control plots when grown without B. tectorum (sucrose × B. tectorum seeding density: F 8,168 = 2.6, P = 0.035; Fig. 3). As shown in Fig. 3, levels of native perennial density observed in sucrose plots not seeded with B. tectorum were similar to those in sucrose plots seeded with B. tectorum at the highest density (1,200 seeds m−2). Increasing seeding density of native species increased establishment of native seedlings during the first growing season after seeding (Fig. 4). However, by the end of the second growing season, numbers of perennial seedlings decreased significantly and were similar among all levels of native seeding treatments (native seeding density × year: F 2,180 = 24.1, P = < 0.0001; Fig. 4).

Mean (±1 SE) number of native perennial plants in control and sucrose addition plots evaluated over all native species seeding densities and for each B. tectorum seeding density (150, 300, 600 and 1,200 seeds m−2). The asterisk indicates significant differences between control and sucrose addition treatments (P < 0.05)

Mean (±1 SE) seedling density of native perennial species evaluated over all B. tectorum seeding densities for each native species seeding density (150, 300 and 600 seeds m−2). Values shown are for the end of the first and second growth year after seeding. Different lowercase letters indicate significant differences across seeding densities and growth years (P < 0.05)
The effect of B. tectorum seeding density on number of native seedlings differed with level of native seeds added (B. tectorum seeding density x native seeding density: F 8,168 = 2.1, P = 0.035). When native species were seeded at 150 and 300 seeds m−2, overall establishment tended to be low and similar numbers of native seedlings established across all B. tectorum seeding levels (Fig. 5). However, when native species were seeded at the highest density (600 seeds m−2), seedling numbers were highest in plots seeded with B. tectorum at 150 seeds m−2 and lowest in plots seeded with B. tectorum at 1,200 seeds m−2 (Fig. 5). An average of 1.8 more native seedlings occurred in plots seeded with B. tectorum at 150 seeds m−2 than in those seeded with B. tectorum at 1,200 seeds m−2.

Mean (±1 SE) seedling density of native perennial species for all B. tectorum seeding densities (0, 150, 300 600 and 1,200 seeds m−2) and native species seeding densities (150, 300, and 600 seeds m−2). Different lowercase letters indicate significant differences across seeding densities of native perennial species and B. tectorum (P < 0.05)
When plots seeded in 2003 were surveyed 3 years after seeding (2006), only the bunchgrasses E. multisetus, P. secunda and P. spicata were present. Numbers of perennial plants observed after 3 years were low (0.1 ± 0.1, 0.3 ± 0.1 and 0.7 ± 0.3 plants m−2, respectively for 150, 300 and 600 native seeds m−2 treatments), but did not differ from those recorded in the previous year (0.6 ± 0.2, 0.7 ± 0.2 and 2.1 ± 0.7 plants m−2, respectively, for 150, 300 and 600 native seeds m−2 treatments).
B. tectorum and native species growth measurements
Sucrose addition negatively affected B. tectorum height (sucrose: F 1,6 = 39.3, P = 0.0008), basal diameter (sucrose: F 1,6 = 18.1, P = 0.0054), and number of tillers per plant (sucrose: F 1,6 = 10.0, P = 0.0019). After the first growing season, B. tectorum height was 18% lower (control = 29.6 ± 0.7 cm vs. sucrose = 24.4 ± 0.8 cm), basal diameter was 31% lower (control = 3.1 ± 0.2 mm vs. sucrose = 2.2 ± 0.1 mm), and number of tillers per plant was 18% less (control = 2.9 ± 0.1 vs. sucrose = 2.4 ± 0.1) (P < 0.05).
Seeding density also affected basal diameter (B. tectorum seeding density: F 3,90 = 5.1, P = 0.0025) and tiller production (B. tectorum seeding density: F 3,90 = 65.1, P = 0.0008) of B. tectorum. Plant diameter was lowest in plots seeded with the highest density (1,200 seeds m−2) (2.1 ± 0.1 mm) and highest in plots seeded with 150 or 600 seeds m−2 (2.9 ± 0.3 mm). Tiller production was lower in plots seeded with the highest density of B. tectorum (2.2 ± 0.1), intermediate in plots seeded with 600 seeds m−2 (2.6 ± 0.2) and higher in plots seeded with 150 or 300 seeds m−2 (2.9 ± 0.2).
Sucrose addition and seeding density treatments had no effect on E. multisetus and P. spicata growth. In the first growing season, E. multisetus seedlings averaged 7.0 (±0.3) cm in height, 3.0 (±0.2) mm in diameter and 2.4 (±0.1) tillers across all treatments. Pseudoroegneria spicata seedlings averaged 7.2 (±0.4) cm in height, 1.4 (±0.1) mm in diameter and had 1.5 (±0.1) tillers. Growth of P. secunda seedlings was not affected by sucrose addition but it was reduced by B. tectorum presence (B. tectorum seeding density; height: F 4,90 = 7.7, P = < 0.0001; diameter: F 4,90 = 7.7, P = < 0.0001; tillers: F 4,90 = 8.1, P = < 0.0001). Plants growing without B. tectorum were taller (1.5 ± 0.4 vs. 0.4 ± 0.1 cm), wider (1.9 ± 0.4 vs. 0.5 ± 0.1 mm) and produced more tillers per plant (2.2 ± 0.5 vs. 0.4 ± 0.1) than plants growing with B. tectorum.
Height and diameter of A. lanulosa seedlings were similar across native seeding treatments in control plots (7.8 ± 1.30 mm tall, 5.5 ± 1.0 mm wide), but increased with increasing native seeding density in sucrose plots (sucrose x native seeding density; height: F 2,90 = 3.4, P = 0.036; diameter: F 2,90 = 3.7, P = 0.027). Seedlings were smaller in sucrose plots seeded with 150 native seeds m−2 (3.6 ± 1.7 mm tall, 3.1 ± 1.4 mm wide) and larger in sucrose plots seeded with 600 native seeds m−2 (5.8 ± 1.0 mm tall, 5.8 ± 1.4 mm wide). Growth of A. lanulosa was not influenced by B. tectorum seeding density.
A. tridentata seedling size was not affected by sucrose addition alone, but in sucrose plots seedlings were smallest in the 150 native seeds m−2 treatment and largest in the 600 native seeds m−2 treatment (sucrose x native seeding density; height: F2,90 = 5.4, P = 0.006; diameter: F2,90 = 6.1, P = 0.003). Diameter of A. tridentata seedlings was larger in plots seeded with B. tectorum (B. tectorum seeding density; diameter: F4,90 = 2.8, P = 0.030). Diameter was smallest in plots without B. tectorum (4.7 ± 1.1 mm wide), but largest in plots seeded with 300 B. tectorum seeds m−2 (6.7 ± 1.0 mm wide).
Because the number of native perennial plants present in 2003-seeded plots was low, plant growth variables obtained for perennial species in year three could not be statistically analyzed. At the end of the third growing season, surviving E. multisetus plants averaged 22.3 (±1.5) cm in height and 14.0 (±2.1) cm in diameter, and had 7.5 (±1.1) tillers and 2.0 (±0.5) inflorescences. P. secunda plants were 8.5 ± 3.3 cm tall, 10.0 ± 0.1 mm wide and had an average of 6.0 (±1.8) tillers and 0.8 (±0.3) inflorescences per plant. P. spicata individuals averaged 23.1 (±2.3) cm in height and 16.7 (±7.5) cm in diameter, and had 7.8 (±0.8) tillers and 0.8 (±0.5) inflorescences.
B. tectorum biomass and seed number
On an individual plant basis, sucrose reduced B. tectorum biomass and seed numbers by 58 and 63%, respectively, during the first year after seeding (sucrose × year; biomass: F 1,192 = 108.1, P = < 0.0001; seeds F 1,192 = 101.7, P = < 0.0001) (Fig. 6a and b). In sucrose amended plots, biomass and seed number per B. tectorum plant were similar in both growth years. In contrast, in control plots biomass and seed number per plant decreased by 72 and 76%, respectively, from the first to the second growth year and were slightly lower than in sucrose plots (Fig. 6a and b).

a Aboveground biomass per plant, b number of seeds per plant, c aboveground biomass m−2, and d number of seeds m−2 of B. tectorum (mean ± 1 SE) in control and sucrose addition treatments at the end of the first and second growth year after seeding. Differentlowercase letters indicate significant differences across sucrose addition treatments and growth years (P < 0.05)
Overall plant biomass and seed numbers per plant decreased with increasing B. tectorum seeding density (B. tectorum seeding density; biomass: F 3,181 = 18.0, P = < 0.0001; seeds: F 3,181 = 14.3, P = < 0.0001). Biomass and number of seeds produced by individual B. tectorum plants were highest in the 150 seeds m−2 treatment (0.34 ± 0.03 g plant−1; 37 ± 4 seeds plant−1) and lowest in the 1,200 seeds m−2 treatment (0.21 ± 0.02 g plant−1; 22 ± 2 seeds plant−1).
On an area basis, B. tectorum biomass and seed numbers were lower in sucrose plots, but only during the first year after seeding (sucrose × year; biomass: F 1,192 = 81.2, P = < 0.0001; seeds: F 1,192 = 46.7, P = < 0.0001). During the first growing season after seeding, sucrose addition decreased biomass by 63% and seed production by 66%, but the effect was no longer significant by the second growing season (Fig. 6c and d). A twofold increase in biomass and seed numbers from the previous year in control plots and a 5–6-fold increase in sucrose plots resulted in similar B. tectorum biomass and seed production on an area basis for both treatments (Fig. 6c).
Biomass and seed number increased with increasing B. tectorum seeding density on an area basis (B. tectorum seeding density; biomass: F 3,181 = 6.6, P = 0.0003; seeds: F 3,181 = 3.1, P = 0.027). Biomass was lower on plots seeded with 150 and 300 seeds m−2 (158.6 ± 20.7 g m−2 and 182.5 ± 24.4 g m−2, respectively) than plots seeded with 1,200 B. tectorum seeds m−2 (221.1 ± 26.3 g m−2). Also, the number of seeds in plots originally seeded at 150 seeds m−2 (16,675 ± 2,299 seeds m−2) was lower than in the highest density plots (21,019 ± 2,238 seeds m−2).
Discussion
Effects of resource availability
Reduction of available NO3 − following sucrose application was consistent with other studies that have shown lower NO3 − levels after adding a labile source of C to the soil (Jonasson et al. 1996; Reever Morgan and Seastadt 1999; Paschke et al. 2000). Addition of sucrose causes immobilization of N in the microbial biomass (Jonasson et al. 1996). Sucrose effects on soil NO3 − were substantial but did not lead to long-term N immobilization. Repeated application of labile sources of C like sucrose are often necessary to promote long-term soil N reduction (Reever Morgan and Seastadt 1999; Paschke et al. 2000).
Sucrose application and the reduction of available NO3 − drastically reduced B. tectorum density and growth during the first year after treatment. Decreased growth of B. tectorum is common following carbon additions and decreases in available soil nitrogen (Young et al. 1999; Paschke et al. 2000; Monaco et al. 2003). Our data also showed that low N availability had strong negative effects on B. tectorum seed production. However, the effects of sucrose on nitrogen availability were ephemeral, and by the second growing season, B. tectorum density markedly increased. As a result, both aboveground biomass and seed production on an area basis were similar for all plots regardless of initial sucrose treatment. Increases in B. tectorum densities reduced growth and seed output of individual plants indicating strong intraspecific competition for available resources (Sheley and Larson 1997). However, aboveground biomass and seed production on an area basis were similar in all plots indicating that constraints acting at the individual plant level may be buffered at the population level. These results agree with Chambers et al. (2007) and provide further evidence that B. tectorum exhibits a high degree of plasticity in seed production in response to variations in available resources. Short-term resource limitations that temporarily reduce B. tectorum density are unlikely to decrease populations of this invasive grass over the long-term.
In contrast to the responses observed for B. tectorum, C addition did not negatively affect establishment of native perennial species, except in bare plots where the invasive grass was not present. Soil nitrate was reduced by sucrose application during the time of perennial seedling establishment regardless of B. tectorum seeding treatment. If lowered nitrate levels were causing reduced perennial establishment in bare plots, significant reductions in native perennial density should have been evident in all sucrose-treated plots. Our results suggest that in plots seeded with low densities of B. tectorum and relatively high densities of natives, the presence of the annual grass may have facilitated native perennial seedling establishment by providing shade and protection from wind and desiccation. Slow-growing perennial species from arid environments have life history traits (e.g., increased leaf longevity, low relative growth rates) that allow them to tolerate low resource levels (Chapin 1980; McLendon and Redente 1992; Aerts and van der Peijl 1993). We observed that B. tectorum was more sensitive to N limitation than perennial species as shown elsewhere (Lowe et al. 2002; Monaco et al. 2003). However, we failed to detect any evidence that native perennial seedlings were favored under N-limiting conditions. While C addition can effectively reduce weed density and biomass, the responses of perennial species often are mixed (Reever Morgan and Seastadt 1999; Blumenthal et al. 2003; Suding et al. 2004). Prior research showed that B. tectorum was reduced and native perennial grass establishment was enhanced after addition of 58 g C m−2 (Young et al. 1999), but long-term additions (160 g C m−2 year−1) (Paschke et al. 2000) may be required to increase abundance of native perennial species. Nitrogen reductions following C additions only occur near the soil surface and, thus, likely have a greater effect on seedlings of the annual B. tectorum than maturing plants of perennials with progressively greater rooting depths.
Effects of propagule availability
We observed a positive relationship between propagule availability and B. tectorum establishment. During the first year, B. tectorum establishment and seed production on an area basis likely were limited by propagule supply. By the second growing season after sucrose application, density and seed availability of B. tectorum increased on an area basis in all treatments. These increases in plant density were the consequence of natural seed rain at the end of the first growing season, which ranged from 5,000 to 15,000 seeds m−2 and was ~4–12-fold higher than the highest initial seeding density of 1,200 seeds m−2 (data not shown). Our findings agree with Humphrey and Schupp (2001) and provide further evidence that short-term depletion of B. tectorum seed banks often are followed by large increases in B. tectorum seed production.
Increasing seed availability of perennial species also enhanced seedling emergence and establishment regardless of sucrose treatment. Propagule limitation is not uncommon (Foster and Tilman 2003; Foster et al. 2004), particularly in early successional habitats (Turnbull et al. 2000), and is a major factor preventing recolonization of invaded and/or disturbed communities (Foster et al. 2004). Seed addition studies generally indicate that native species establish or increase in abundance as a result of seed addition (Hamilton et al. 1999; Turnbull et al. 2000; Foster and Tilman 2003; Sheley et al. 2006). Also, higher establishment of native perennial grasses has been theorized and observed with increasing seeding density in other invaded rangelands of the Western US (Pyke and Archer 1991; Sheley et al. 2006). Despite increased emergence with higher seeding densities, we observed generally low levels of seedling emergence over the 2 years of our experiment. The levels of seed addition that we used (150–600 seeds m−2) may still have been limiting to native seedling establishment, but other factors also played a significant role.
Competitive interactions and indirect effects
Generally low survival of native perennial seedlings during the study likely resulted from a combination of different factors. Native perennial species establishment clearly was affected by competition with B. tectorum. Native perennial seedlings were able to emerge and coexist with B. tectorum if B. tectorum density did not exceed 300 plants m−2 (initial seeding density <600 B. tectorum seeds m−2). However, when B. tectorum density increased to ~450 B. tectorum plants m−2 (seeding density = 1,200 B. tectorum seeds m−2), native perennial establishment declined indicating a less favorable environment for native seedlings. Despite relatively high first year survival of perennial seedlings, few individuals survived through the second growth year. Seedlings of perennial sagebrush steppe species often exhibit low survival rates due to harsh environmental conditions (Pyke 1990; Chambers 2000; Chambers and Linnerooth 2001). Precipitation during the active growing season (March–June) of the second year was only about 50% of the long-term average and likely contributed to the low survival rates. Also, the substantial increase in B. tectorum density during the second year markedly increased competition for both soil water and N and reduced the probability of native perennial seedling survival. Although many native perennial species are capable of tolerating relatively low resource conditions (Chapin 1980), seedlings of native perennial species are generally poor competitors for soil resources as has been shown for sagebrush steppe species (Arredondo et al. 1998; Booth et al. 2003; Humphrey and Schupp 2004) and for the native perennial grass, Nasella pulchra, in invaded California grasslands (Hamilton et al.1999).
Although B. tectorum may suppress native perennial seedlings, the outcome of competition between annual invaders and native perennials may shift with life stage. Mature native perennials are more deeply rooted, have larger root masses, and are capable of efficiently capturing soil resources and limiting available water and nutrients for annuals invaders (Yoder and Caldwell 2002; Booth et al. 2003). Despite low initial survival of seeded perennials, individuals that survived beyond the second year were able to reproduce in the third year. These results indicate that native perennials have the potential to establish and form viable populations in invaded areas.
Conclusions
Soil nitrogen availability is clearly a limiting factor for B. tectorum. Establishment, growth and reproduction of this annual invader decreased under reduced N availability but both the sucrose-induced decrease in soil N and its effects on B. tectorum were short-lived. As predicted, establishment of perennial natives was less affected by lowered N, but native perennials did not appear to gain any competitive advantage when N was reduced. Bromus tectorum was negatively affected by intense intraspecific competition whereas native seedlings were more likely to be affected by interspecific competition with the annual grass. The effects of B. tectorum on native seedlings were density dependent and appear to involve thresholds below which native seedlings can successfully establish and persist. The competitive effects of B. tectorum on the native species were greatest at the seedling stage including the first 2 years of establishment. Establishment of native perennials was low, but individuals that survived the first 2 years lived through the third year and produced seed. Mature individuals can be extremely competitive for available resources (Booth et al. 2003; Humphrey and Schupp 2004) indicating that the seedling stage is critical for successful reintroduction of native species into B. tectorum-dominated sites.
Longer term reductions of soil N could increase the ability of perennial species to establish in these degraded ecosystems (McLendon and Redente 1992; Paschke et al. 2000). Because sugaring is not a viable approach for large areas, an alternative and more realistic approach for reducing available nitrogen could be to promote resource use by increasing abundance and cover of residual perennial vegetation (Prober et al. 2005; Prober and Lunt 2009). Removal of resident perennial vegetation through poorly timed or intensive livestock grazing and other disturbances has decreased the resistance of sagebrush ecosystems to B. tectorum throughout the region (Anderson and Inouye 2001; Chambers et al. 2007). Propagule supply clearly plays a major role in structuring the plant community during the process of vegetation recovery (D’Antonio et al. 2001; Foster et al. 2004). In contrast to B. tectorum, which is neither seed limited nor dispersal limited, native perennial species are seed limited due to depleted seed banks, the absence of mature seed-bearing individuals in the community, and limited dispersal from surrounding areas (Young and Evans 1989). Adding propagules of native perennial species can partially overcome this limitation, but the numbers required are large. Common seeding rates used in the region are about 150–300 seeds m−2, but our results and previous research (977–1,577 seeds m−2 in Sheley et al. 2006) indicate that higher seed availability could be necessary to increase seedling establishment. Regional propagule pools determine the availability of key species and, for that reason, play a major role in community assembly and resilience after disturbance (Foster et al. 2004). Restoration of propagule pools at both the local (i.e. seed bank) and at the landscape level (i.e. dispersal) could increase the likelihood of native seedling establishment and enhance the probability of native propagule availability after disturbances or in years when environmental conditions are more conducive to native seedling establishment.
References
Aerts R, van der Peijl MJ (1993) A simple model to explain the dominance of low-productive perennials in nutrient-poor habitats. Oikos 66:144–147
Anderson JE, Inouye RS (2001) Landscape-scale changes in plant species abundance and biodiversity of a sagebrush steppe over 45 years. Ecol Mon 71:531–556
Arredondo JT, Jones TA, Johnson DA (1998) Seedling growth on Intermountain perennial and weedy annual grasses. J Range Manage 51:584–589
Association of Official Seed Analysts (2000) Tetrazolium testing hand book. Contribution no. 29 to the handbook on seed testing. Las Cruces, NM
Beckstead J, Auspurguer CK (2004) An experimental test of resistance to cheat grass invasion: limiting resources at different life stages. Biol Invasions 6:417–432
Blumenthal DM, Jordan NR, Russelle MP (2003) Soil carbon addition controls weeds and facilitates prairie restoration. Ecol Appl 13:605–615
Booth MS, Caldwell MM, Stark JM (2003) Overlapping resource use in three great basin species: implications for community invisibility and vegetation dynamics. J Ecol 91:36–48
Burke MJW, Grime JP (1996) An experimental study of plant community invisibility. Ecology 77:776–790
Chambers JC (2000) Seed movements and seedling fates in disturbed sagebrush steppe ecosystems: implications for restoration. Ecol Appl 10:1400–1413
Chambers JC, Linnerooth AR (2001) Restoring riparian meadows currently dominated by artemisia using alternative state concepts: the establishment component. Appl Veg Sci 4:157–166
Chambers JC, Roundy BA, Blank RR, Meyer SE, Whittaker A (2007) What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum? Ecol Mon 77:117–145
Chapin FS (1980) The mineral nutrition of wild plants. Ann Rev Ecol Syst 11:233–260
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire cycle, and global change. Ann Rev Ecol Syst 23:63–87
D’Antonio C, Levine J, Thomsen M (2001) Ecosystem resistance to invasion and the role of propagule supply: a California perspective. J Mediterranean Ecology 2:233–245
Davis MA (2005) Invasibility: the local mechanism driving community assembly and species diversity. Ecography 28:696–704
Davis MA, Grime JP, Thompson K (2000) Fluctuating resources in plant communities: a general theory of invasibility. J Ecol 88:528–534
Denny DW (2002) Soil survey of Humboldt County, Nevada, east part. Part 1. United States Department of Agriculture, Natural Resources Conservation Service
DiVittorio CT, Corbin JD, D’Antonio CM (2007) Spatial and temporal patterns of seed dispersal an important determinant of grassland invasion. Ecol Appl 17:311–316
Foster BL, Dickson TL (2004) Grassland diversity and productivity: the interplay of resource availability and propagule pools. Ecology 85:1541–1547
Foster BL, Tilman D (2003) Seed limitation and the regulation of community structure in oak savanna grassland. J Ecol 91:999–1007
Foster BL, Dickson TL, Murphy CA, Karel IS, Smith VH (2004) Propagule pools mediate community assembly and diversity-ecosystem regulation along a grassland productivity gradient. J Ecol 92:435–449
Hamilton JG, Holzapfel C, Mahall BE (1999) Coexistence and interference between a native perennial grass and non-native annual grasses in California. Oecologia 121:518–526
Hooper DU (1998) The role of complementarity and competition in ecosystem responses to variation in plant diversity. Ecology 79:704–719
Humphrey LD, Schupp EW (2001) Seed banks of Bromus tectorum-dominated communities in the Great Basin. West N Am Nat 61:85–92
Humphrey LD, Schupp EW (2004) Competition as a barrier to establishment of a native perennial grass (Elymus elymoides) in alien annual grass (Bromus tectorum). J Arid Environ 58:405–422
Jonasson S, Vestergaard P, Jensen M, Michelsen A (1996) Effects of carbohydrate amendments on nutrient partitioning, plant and microbial performance of a grassland-shrub ecosystem. Oikos 75:220–226
Kemp PR (1983) Phenological patterns of Chihuahuan desert plants in relation to the timing of water availability. J Ecol 71:427–436
Knapp PA (1996) Cheatgrass (Bromus tectorum L) dominance in the Great Basin desert: history, persistence, and influences to human activities. Global Environ Change 6:31–52
Koniak S, Everett RL (1982) Seed reserves in soils of successional stages of pinyon woodlands. Am Midl Nat 108:295–303
Link SO, Keeler CW, Hill RW, Hagen E (2006) Bromus tectorum cover mapping and fire risk. Int J Wildland Fire 15:113–119
Lowe PN, Lauenroth WK, Burke IC (2002) Effects of nitrogen availability on the growth of native grasses and exotic weeds. J Range Manage 55:94–98
Lowe PN, Lauenroth WK, Burke IC (2003) Effects of nitrogen availability on competition between Bromus tectorum and Bouteloa gracilis. Plant Ecol 167:247–254
McLendon TE, Redente EF (1991) Nitrogen and phosphorus effects on secondary succession dynamics on a semi-arid sagebrush site. Ecology 72:2016–2024
McLendon TE, Redente EF (1992) Effects of nitrogen limitation on species replacement dynamics during early secondary succession on a sagebrush site. Oecologia 91:312–317
Melgoza G, Nowak RS (1991) Competition between cheat grass and two native species after fire: implications from observations and measurements of root distribution. J Range Manage 44:27–33
Monaco TA, Johnson DA, Norton JM, Jones TA, Connors KJ, Norton JB, Redinbaugh MB (2003) Contrasting responses of Intermountain West grasses to soil nitrogen. J Range Manage 56:282–290
Neary DG, Klopatek CC, DeBano LF, Ffolliott PF (1999) Fire effects on belowground sustainability: a review and synthesis. For Ecol and Manage 122:51–71
Paschke MW, McLendon T, Redente EF (2000) Nitrogen availability and old-field succession in a shortgrass steppe. Ecosystems 3:144–158
Prober SM, Lunt ID (2009) Restoration of Themeda australis swards suppresses soil nitrate and enhances ecological resistance to invasion by exotic annuals. Biol. Inv 11:171–181
Prober SM, Thiele KR, Lunt ID, Koen TB (2005) Restoring ecological function in temperate grassy woodlands: manipulating soil nutrients, exotic annuals and native perennial grasses through carbon supplements and spring burns. J Appl Ecol 42:1073–1085
Pyke DA (1990) Comparative demography of co-occurring introduced and native tussock grasses: persistence and potential expansion. Oecologia 82:537–543
Pyke DA, Archer S (1991) Plant-plant interactions affecting plant establishment and persistence in revegetated rangeland. J Range Manage 44:550–557
Rau BM, Blank RR, Chambers JC, Johnson DW (2007) Prescribed fire in a great basin sagebrush ecosystem: dynamics of soil extractable nitrogen and phosphorus. J Arid Environ 71:362–375
Reever Morgan KJ, Seastadt TR (1999) The effects of soil nitrogen reduction on non-native plants in restored grasslands. Rest Ecol 7:51–55
SAS Institute Inc. (2002) SAS 9.1.2 software. Cary, NC, USA
Schuman GE, Reeder JD, Manley JT, Hart RH, Manley WA (1999) Impact of grazing management on the carbon and nitrogen balance of a mixed-grass rangeland. Ecol Appl 9:65–71
Sheley R, Larson L (1997) Cheat grass and yellow star thistle growth at three soil depths. J of Range Manage 50:146–150
Sheley RL, Mangold JM, Anderson JL (2006) Potential for successional theory to guide restoration of invasive-plant-dominated rangeland. Ecol Mon 76:365–379
Suding KN, LeJeune KD, Seastadt TR (2004) Competitive impacts and responses of an invasive weed: dependencies on nitrogen and phosphorus availability. Oecologia 141:526–535
Thomsen MA, D’Antonio CM (2007) Mechanisms of resistance to invasion in California grassland: the roles of competitor identity, resource availability, and environmental gradients. Oikos 116:17–30
Tilman D (1997) Community invasibility, recruitment limitation, and grassland biodiversity. Ecology 78:81–92
Turnbull LA, Crawley MJ, Rees M (2000) Are plant populations seed-limited? A review of seed sowing experiments. Oikos 88:225–238
Von Holle B, Simberloff D (2005) Ecological resistance to biological invasion overwhelmed by propagule pressure. Ecology 86:3212–3218
Wehking PM (2002) The role of the seed bank in the restoration of a big sagebrush dominated riparian corridor to a dry meadow. MSc. Thesis, University of Nevada-Reno, USA
West NE, Young JA (2000) Vegetation of Intermountain valleys and lower mountain slopes. In: Barbour MG, Billings WD (eds) North American terrestrial vegetation. Cambridge University Press, New York, NY, pp 255–284
Western Research Climate Center (2007) Historical climate information. Available http: www.wrcc.dri.edu. Accessed 2 Dec 2007
Whisenant SG (1990) Changing fire frequencies on Idaho’s Snake River plains: ecological and management implications. Proceedings—symposium on cheatgrass invasion, shrub die-off and other aspects of shrub biology and management. USDA General Technical Report INT-276, Ogden, Utah, USA
Yang JE, Skogley EO, Georgitis SJ, Schaff BE, Ferguson AH (1991) Phytoavailability soil test: development and verification of theory. Soil Sci Soc Am J 55:1358–1365
Yoder C, Caldwell M (2002) Effects of perennial and nitrogen pulses on growth and nitrogen uptake of Bromus tectorum. Plant Ecol 158:77–84
Young JA, Evans RA (1989) Dispersal and germination of big sagebrush (Artemisia tridentata) seeds. Weed Sci 37:201–206
Young JA, Evans RA, Eckert RE (1969) Population dynamics of downy brome. Weed Sci 17:20–26
Young JA, Blank RR, Clements CD (1999) Nitrogen enrichment and immobilization on the dynamics of an annual grass community. In: Eldridge D, Freudenberger D (eds) Proceedings of the VI international rangeland congress. Townsville, Queensland, Australia
Acknowledgments
We thank Carlos Wilson, Jacob Landmesser, Kendra Moseley, Lisa Ellsworth and Tye Morgan for technical assistance. George Fernandez, Dave Turner and Dave Board provided statistical advice, and Thomas A. Jones, Thomas Monaco, Dan Ogle and Loren St. Jones helped with study implementation. Comments from Carla D’Antonio, Dale Johnson, Tara Forbis and anonymous reviewers improved the manuscript. Research was funded by the USDA Initiative for Future Agricultural and Food Systems (CREES Agreement 2001-52103-11322) with additional support from the USDA Forest Service, Rocky Mountain Research Station, the Nevada Agricultural Experiment Station, and the USDI Bureau of Land Management. Any use of trade names is for descriptive purposes only and does not imply endorsement by U.S. government.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mazzola, M.B., Chambers, J.C., Blank, R.R. et al. Effects of resource availability and propagule supply on native species recruitment in sagebrush ecosystems invaded by Bromus tectorum . Biol Invasions 13, 513–526 (2011). https://doi.org/10.1007/s10530-010-9846-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-010-9846-0