
Abstract
Biological and habitat characterization of the non-indigenous invasive species Xenostrobus securis was undertaken in the Ria de Vigo. This study included genetic identification of mussel samples collected from introduced and endemic areas, and the assessment of mussel size, population abundance, geographic distribution, pathological condition, and sediment composition of substrata type. The mussel had a marked patchy distribution, being more abundant in brackish sites with fine sediments and high organic matter content. Pathological analysis revealed that X. securis does not play any role as vector for introducing allochthonous pathogens in the Ria de Vigo. Nevertheless, depending on its invasiveness potential, the mussel could be a key host favouring spreading and epizootic outbreaks of marteliosis which is known to be harmful for local bivalve populations. Phylogenetic analyses of the COI gene placed all the resulting sequences in a clade within the genus Xenostrobus and its phylogeny congruent with an Australian/Pacific origin. The COI tree suggests two historical introductions in European waters. One of these invasions seems to have started in Galicia, moving from there towards Italy and France, while the geographical spread of the second invasion cannot be deciphered, although the Australian/Pacific origin of this invasion seems very possible. The 18S network is congruent with one invasion starting in Galicia or in Italy, as the Australian haplotype is closely related to the haplotype found in these areas. Several hypotheses accounting for the colonization history of this species in Galician waters are discussed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The transfer and introduction of alien species is considered by scientists and policy-makers one of the major threats to both structural and functional biodiversity of marine ecosystems (Mack et al. 2000; Sousa et al. 2009). The first vector in marine alien species introductions is canals, following by shipping and aquaculture (Streftaris et al. 2005; DAISIE 2009).
Recipient (and probably also donor) areas for animal invasions in the Galician waters are mainly located in the Rias, which have been traditionally considered by naturalists as “hot spots”—even invasion corridors—at high risk for alien introductions from all over the world. In fact, today ca. 100 aliens have been recorded in Galician waters (Quilez-Badia 2009), most of which have been able to establish harmful reproducing populations, especially in estuaries supporting highly productive shellfish farming (Bañón et al. 2007). Garci et al. (2007) first reported the establishment of the alien mussel Xenostrobus securis (Lamarck, 1819) in the Ria de Vigo colonizing inner estuarine waters where it exhibits a wide ecological tolerance. This black-pigmy mussel is considered an important threat as bioinvasor with a negative impact on the introduced ecosystem (Darrigran 2002). It is endemic to the brackish waters of New-Zealand and Australia (www.ciesm.org). It also has been recorded as invading species inhabiting the Central Tyrrhenian Sea (Giusti et al. 2008), Italian coastal lagoons (Sabelli and Speranza 1994; Lazzari and Rinaldi 1994), delta of the river Po (Russo 2001), French Mediterranean lagoons and Japan (Gofas and Zenetos 2003; Kimura et al. 1999; Kohama et al. 2001).
The aim of this paper was recording the size, population abundance, geographic distribution, pathological condition, and sediment composition of substrata type of the non-indigenous invasive species X. securis at the Ria de Vigo, as well as to provide some insight into the invasion history of this species in European waters.
Materials and methods
Study site, samples and sediment characterization
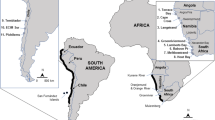
Sampling was carried out at five stations in two transects parallel to the coast, by scuba diving between 1 and 20 m depths, once during three consecutive summers (2006–2008) in the Ria de Vigo (42°18′N–8°37′W) (Fig. 1): near by the mouth of the Verdugo river at the inner inlet of Ria de Vigo (Station 1), Alvedosas (Station 2), Arcade (Station 3); Punta Cabalo (Station 4) and San Simón (Station 5). Where colonies of pigmy mussels were observed, three 20 × 20 cm replicate squares were sampled at each station. Specimens collected (Sample A) were transported to the laboratory, counted and measured for shell length (L), height (H) and width (W) using a digital calliper (Aerospace, 0.01 mm accuracy). Data obtained were extrapolated to account for the number of individual per m2 in colonies. Sediment samples were also collected at each station with a rectangular dredge standardized to 20 l. Granulometric fractions of the sediments, mineralogical and element composition analyses were undertaken by the standard dry-sieve for sand fraction and settling method for mud fraction, X-ray diffraction and X-ray fluorescence.

Location of sampling stations at the Ria de Vigo (NW Spain). 1. Pontesampaio; 2. Alvedosas; 3. Arcade; 4. Punta Cabalo; 5. San Simón
Two samples of 15 mussels were taken from stations 1 and 3 on June 2006 and another sample of 30 individuals was harvested from station 3 on June 2007. These 60 specimens (Sample B) were used for histopathological analyses.
Additional samples of mussels from stations 1 and 3 in 2008 (Sample C) were fixed in ethanol for genetic study. We also obtained samples from endemic Australian populations, and also from other European localities (France at Grau-du-Roi Camargue and Italy-Venezia).
Molecular characterization of the pigmy mussel
Genomic DNA extraction, PCR amplification and sequencing
DNA extractions were performed employing the DNAzol reagent® (Invitrogen Life Technologies™) according to the manufacture’s instructions. The primers LCO1490 and HCO2198 (Folmer et al. 1994) (Table 1) were used to amplify approximately 700 bp of the cytochrome oxidase c subunit I (COI) mitochondrial gene. For amplification of approximately 1,800 bp of the small subunit (18S) ribosomal RNA gene, the primers 16S-A and 16S-B (Medlin et al. 1988) were employed.
PCR reactions were performed in a total volume of 25 μl containing 1 μl of genomic DNA (150–200 ng), PCR buffer at 1× concentration, 1.5 mM MgCl2, 0.2 mM nucleotides (Roche Applied Science), 0.3 μM primers and 0.625 U Taq DNA polymerase (Roche Applied Science). The cycling protocol for the 18S rRNA gene was 2 min at 94°C, 35 cycles of 30 s at 94°C, 1 min at 55°C and 2 min at 72°C, followed by 7 min at 72°C. The cycling protocol for the COI gene was 2 min at 94°C, 35 cycles of 30 s at 94°C, 30 s at 50°C and 1 min at 72°C, followed by 7 min at 72°C. All PCRs were carried out in a TGradient thermocycler (Biometra) and a negative control (without DNA) was included for each set of PCRs. PCR products were separated on a 2% agarose (in 1× Tris–acetic EDTA buffer) gel, stained with ethidium bromide and scanned in a GelDoc XR documentation system (Bio-Rad Laboratories).
PCR products were purified for sequencing using the commercial kit Rapid PCR Purification system (Marligen Biosciences, Inc.) according to the manufacture’s instructions. The sequence reactions were performed on an ABI PRISM™ 3100 (Applied Biosystems) using a BigDye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems). All amplified fragments were sequenced with forward and reverse primers and the chromatograms were analysed using ChromasPro version 1.32 Technelysium Pty Ltd A. To obtain the entire sequences of the 18S rRNA gene, specific primers Xen1, Xen2 and Xen3 (Table 1) were designed using Primer-3 (Rozen and Skaletsky 2000).
Phylogenetic analysis
Six additional COI sequences were downloaded from GenBank: two sequences from X. atratus (AB298598, AB298599) (Kimura et al. 2007 submitted), one sequence from X. pulex (DQ917582) (Wood et al. 2007), and three sequences from Mytilus edulis, Mytilus galloprovincialis and Modiolus areolatus (DQ917606, DQ917605 and DQ917604, respectively) (Wood et al. 2007). Three 18S sequences from the latter species were also included in the analysis (L33448, L33451 and AF124210, respectively) (Wood et al. 2007). Sequences were aligned using M-COFFEE (Wallace et al. 2006) and corrected by eye. The final alignments include 21 COI sequences 567 bp long and 15 18S sequences 1,660 bp long. Best-fit nucleotide substitution models were selected by the AIC (Akaike 1974) criterion with the ModelTest web server (Posada 2006) and PAUP* (Swofford 2000). Bayesian phylogenetic analyses were implemented with the program Mr. Bayes 3.1.2 (Huelsenbeck and Ronquist 2001). Two independent runs with four MCMC chains each (three heated and one cold) were carried out for 11 million generations, with trees being sampled every 1,000 generations. Runs were checked for convergence, and the first million trees were discarded as “burnin”. A 50% majority rule consensus tree was computed from the last 10 million trees sampled. In addition, and in order to recover the relationships between closely related sequences, we built COI and 18S haplotype networks using statistical parsimony (Templeton et al. 1992) as implemented in the program TCS v.1.23 (Clement et al. 2000).
Pathological condition
Histopathology
Mussels from sample B were shucked and the soft parts were examined for lesions and gross abnormalities. An approximately 5 mm thick section of meat containing mantle, gills, foot and visceral mass was fixed in Davidson’s solution and embedded in paraffin; 5 µm thick sections were stained with Harris’ hematoxylin and eosin (Howard and Smith 1983) and examined under light microscopy for histopathological analysis.
Genomic DNA extraction, PCR amplification, cloning and sequencing of Marteilia
Molecular methods were applied for the identification of Marteilia cells infecting the pigmy mussels. Two distinct sources of DNA were employed: (a) two paraffin blocks in whose histological sections infection by Marteilia sp. was diagnosed (accordingly with the methodology by López-Flores et al. (2008); and (b)) tissues of pygmy mussels from sample B. The universal primers used are indicated in Table 1. Primers 16S-A and 16S-B (Medlin et al. 1988) were employed as amplification controls. Three PCR assays for Marteilia were carried out: (a) a specific PCR for M. sidneyi using the primers PRO2/LEG1 (Kleeman and Adlard 2000); (b) a specific PCR for M. refringens using the primers SAS2/SS2 (Le Roux et al. 1999); and (c) a nested PCR using the primer pairs MT1/MT2 for the first-round of PCR and MT1B/MT2B for the second-round of PCR (López-Flores et al. 2004). PCR conditions followed Kleeman and Adlard (2000), Le Roux et al. (1999) and López-Flores et al. (2004). Amplified products, showing the expected size, were ligated into cloning vector pCR2.1 at 14°C overnight and transformed into E. coli One Shot Top 10F′ Chemically Competent (Invitrogen Life Technologies™). Transformed cells were screened by PCR using the vector’s M13 forward and reverse primers. One positive clone from each sample was selected for sequencing as previously described.
Results
Biological and habitat characterization
Table 2 shows the range of shell morphometric parameters for the X. securis samples. Figure 2a–d illustrates a typical specimen from the Ria de Vigo. Pigmy mussels showed different size depending on the sampling station (Table 2), always located forming patches settled on arrays of rocks and soft-sediments (Fig. 2e, f). The patch density of X. securis varied among sampling sites (Table 2), although not between years. During the sampling operations in stations 1 and 3, where a vertical wall existed, it was observed that the distribution of the pigmy mussels was uniform across different substrata. Two interesting features were also noted. First, large patches were found at large rocks as well as at medium or small rocks. This pattern was also observed at soft bottoms. Second, pigmy mussels also settled on natural beds of exploited local populations of mussels (M. galloprovincialis) and oysters (Ostrea edulis) (Fig. 2g, h).

Shell morphology of Xenostrobus securis (a, b, c and d) from the Ria de Vigo. Typical patches of X. securis in rocky (e) and muddy (f) substrata as seen by scuba diving. The exploited bivalves Mytilus galloprovincialis (g) and Ostrea edulis (h) are used as substrata for X. securis settlement in stations 1 and 3
Sediment characteristics of sampling sites are presented in Table 3. Individuals were found in all types of sandy and muddy fine sediments in the brackish waters of the inner part of the Ria de Vigo. The highest density occurred in those places that showed the highest values of organic matter (Table 3). Elemental analysis of sediments revealed high concentrations of toxic elements (lead, nickel and vanadium) mostly in stations 1 and 5.
Genetic characterization
The 18S Bayesian consensus phylogeny was very uniformative, and all the ingroup sequences formed a completely unresolved single clade (data not shown). The 18S haplotype network indicated that Galician and Italian samples share the same haplotype. Morover, the Australian haplotype is closest to this Galician/Italian haplotype (Fig. 3a). On the other hand, the COI Bayesian tree was well resolved (Fig. 3b). In this tree, the Xenostrobus sequences formed a highly supported monophyletic ingroup, with high posterior probability (0.98). Within this group, the three species (X. securis, X. atratus and X. pulex) formed distinct and well-supported clades, although the relationships between them remained ambiguous. Within X. securis four main lineages could be distinguished (clades I–V, Fig. 3b). Two of these lineages (III and IV) are Australian, while the other two (I and II) include sequences from Italy, France and Galicia. The relationships between these four lineages were not well resolved. No single haplotype network could be reconstructed for COI due to the higher level of divergence. The TCS analysis resulted in SIX different subnetworks that could not be connected under the 95% parsimony limit (data not shown). Nucleotide sequence data reported for X. securis are available in the GenBank under the accession numbers FJ949108-FJ949122 for COI gene and FJ949123–FJ949134 for 18S rRNA gene.

Phylogenetic analyses. a 18S haplotype network. Each circle corresponds to a different haplotype. Circle sizes are proportional to haplotype frequency. The connections between the circles indicate 1 nucleotidic change. Xenostrobus securis sequences are identified by the clone number followed by a country code (AUS: Australia, FR: France, GAL: Galicia, IT: Italy. b Bayesian COI 50% majority rule consensus tree. Xenostrobus securis sequences are identified by the clone number followed by a country code (AUS: Australia, FR: France, GAL: Galicia, IT: Italy). Numbers above branches are posterior probabilities
Pathology
Histopathological analysis revealed some pathological conditions (Table 4). Marteilia sp. was detected in two of the three samples, with the highest prevalence (27%) at station 3 on June 2007. In this sample, six individuals showed light infections characterised by primary cells, up to 15 µm long, enclosing various secondary cells, located in the epithelia of the stomach and digestive primary ducts (Fig. 4a). Moreover, two additional mussels had more advanced infections, with sporulation stages located in the epithelium of the digestive diverticula (Fig. 4b). Mature sporulated stages consisted of primary cells, up to 30 μm long, including up to eight sporonts, which enclosed up to four spores (Fig. 4b).

Histological sections of Xenostrobus securis infected by Marteilia refringens. a Primary cells of the parasite (arrows) enclosing various secondary cells located in the epithelia of the stomach. b Sporulation stages of the parasite (arrows) in the epithelium of the digestive diverticula
Intracellular colonies consisting of Rickettsia-like organisms, up to 50 μm in diameter, were occasionally found in digestive epithelia. Most of them were located in the stomach epithelium and some in the epithelium of secondary ducts and digestive tubules. The infection intensity was low in every case.
A neoplastic condition affecting gonad was detected in two mussels at station 1 (Table 4), characterised by a mass proliferation of germ cells, and which did not give rise to ripe gametes. One of the affected mussels showed gonad areas with normal aspect (i.e. with gonad follicles enclosing ovocytes, some of which were atresic), intermixed with gonad areas with follicles filled with high numbers of undifferentiated basophilic cells and abundant mitotic figures (Fig. 5a). Some of these undifferentiated cells were seen in haemolymph sinuses out of the follicles. Normal follicles were not observed in the other affected mussel. Every gonad follicle was partially filled with abnormal cells enclosing a very large (up to 18 µm long) lobed nucleus with peripheral chromatin and up to 4 nucleoli (Fig. 5b). Mitotic figures were also abundant. Some cells inside the follicles showed numerous filaments that could correspond to prophase chromosomes (Fig. 5c). Ripe ovocytes were observed in some of the follicles together with the abnormal cells.

Histological sections of Xenostrobus securis affected by gonad neoplasia. a Mantle areas with gonad follicles filled with high numbers of undifferentiated very basophilic cells. b Mass of abnormal cella (arrows) with very large nucleus enclosing various nucleoli inside a gonad follicle. c Cells showing numerous filaments (arrows) that could correspond to prophase chromosomes inside a gonad follicle. (M mitotic figure)
Two mussels from the station 1 showed an abnormal condition characterised by mineral deposits forming concentric layers in an area of the mantle (Table 4), which probably correspond to pearls. In fact, two pearls (2 mm in diameter) were found in one of those mussels when macroscopic examination of the meat was performed.
Negative PCR results were observed for DNA isolated from two paraffin blocks using the different Marteilia primers, whereas the internal control of amplification was positive for both DNAs. Negative PCR results were also obtained for DNA isolated from fixed tissues with the primers PRO2/LEG1 and SAS2/SS2. However, positive results were obtained for five samples (5/30) by nested PCR. BLAST searches showed closest identity with IGS gene sequence of Marteilia refringens infecting Mytilus galloprovincialis (AJ629366; AJ629361; AJ629358) (López-Flores et al. 2004) and from other intermediate crustacean hosts as Brachyura zoeae (AM504144), Acartia discaudata (AM504142), A. clausi (AM504139), Oithona sp. (AM504138) and Euterpina acutifrons (AM504137) (Carrasco et al. 2007). Nucleotide sequence data reported for M. refringens are available in the GenBank under the accession numbers EU854303–EU854307.
Discussion
Colonization history
The COI phylogeny confirms the adscription of the individuals sampled from the Ria de Vigo as Xenostrobus securis. Although the relationships among the four main X. securis lineages could not be resolved, the inferred phylogeny is congruent with an Australian origin. The fact that the European clade II is more closely related, and with high support, to an Australian sequence than to other European group (clade I) is remarkable. However, the exact origin can not be confirmed in the absence of more samples from the source area. For example, it would have been very informative to have a sample from Japan, where most of the aquaculture trade originates from-particularly Ruditapes philippinarum—which could have vectored the first invasion to Europe. In any case, the fact that the European samples come from two distinct clades implies two distinct colonisations. The basal position of the Galician sample within clade I suggests that one of these invasions could have started in Galicia, moving from there towards Italy and France, while the geographical spread of the second invasion (clade II) cannot be deciphered, although the Australian origin of this invasion seem very likely in the light of the available data. The 18S network is congruent with one invasion starting in Galicia or in Italy, as the Australian haplotype is most closely related to the haplotype found in these areas. As mentioned above, inferences are contingent on the available sequences. To confirm these hypotheses it would be necessary to obtain more sequences from these and other geographical locations in Asia and Europe.
Xenostrobus securis is registered in Galicia at least from 1995 (Bañón et al. 2008; J.E. Trigo, personal communication). Although ship ballast usually has been considered the source of introduction to many invasive species (Carlton 1985), this vector is anyway a most unlikely way for this species, which settles mostly in close lagoons and shallow depths. Furthermore, it is doubtful that X. securis can attach to a hull were it rather nestles in the muddy lining of the bottom (either soft or hard). In fact, the presence of this species in the Adriatic Sea since 1992 was related with intense shellfish farming in the area (Occhipinti-Ambrogi 2000; Zenetos et al. 2004)). Although local laws are quite restrictive controlling the origin, transport purification or transformation and, definitively, granting authorizations for sale or reallocation of any marine live species in Galician waters since 1993, it is not possible to reject an uncontrolled seed introduction of some bivalve molluscs from Australia-New Zealand-Pacific as the origin of X. securis in the Ría de Vigo.
On the other hand, although our 18S network and COI tree (clade 2) show the Australian origin of this invasion, they do not allow us to indicate whether or not the European introduction went through the Adriatic or Galicia, via or not from the French lagoons. An aquaculture connection from other areas to Galicia is more likely the potential vector since there is a consistent trade of spat, which may result in multiple invasions.
Biological and habitat characterization
Although significant size differences were observed among individuals from different sampling sites, no significant differences were found in shell size at each station in different years. It is likely that these differences may be due to phenotypical response to different environmental and/or ecological conditions existing in the five sampling sites, providing inter-population preferential settlement of mussels, typical of in invertebrate larvae (Snelgrove et al. 1993; Eckman et al. 1994; Turner et al. 1994). In fact, the relationship between molluscan density and sediment characteristics in estuarine ecosystems is well-known (Sousa et al. 2005, 2006), as well as the advection and deposition of larvae (passive processes) having a profound impact on settlement and thus on population abundance and distribution patterns (Eckman 1983; Snelgrove 1994).
Our observations on substrata type and size/density of individuals within patches suggest that the settlement of X. securis in the Ria de Vigo does not result from an active larval behavioural preference of substrate utilization. Instead, it is likely that pigmy mussels accumulate in rocks or sediments of different types leading to an aggregated distribution mainly because they are passively suspended and retained in these locations. Nevertheless, post-settlement movements along the substrate could also contribute to the final spatial aggregated pattern observed as indicated by Crisp (1974). A lack of microhabitat selection is supported by our observations during sampling operations. The presence of X. securis in all types of substrata in stations influenced by the tidal cycle (stations 1 and 3) suggests that larvae could be transported during different portions of the tidal cycle at heights varying from soft-bottom when tide is low, up to 2 m (hard rock) when the tide is high. This observation is also supported by the fact that schools of mussels at both soft and rocky bottoms samples are very similar in size, reflecting synchronicity in larval release, dispersal and settlement.
Pathology
Tissues stored as paraffin blocks are a potential source of DNA. Nevertheless, the degradation of DNA strands, in fragments of hundreds bps, is the most significant problem in analyzing DNA extracted from paraffin blocks. In our study the internal control of amplification was positive, which indicate the integrity of the DNA samples. False negative PCR results obtained from paraffin blocks can be due to the low intensity of infection by Marteilia-like parasite. Thus, the identification of Marteilia species infecting X. securis was only possible using ethanol-fixed material and after amplification by nested-PCR. The sequences obtained support the presence of Marteilia refringens infecting X. securis in the Ria of Vigo. Surveys performed in late 1980s showed that this parasite infects mussels Mytiuls galloprovincialis in the Galician Rias, including Ria de Vigo, causing death or sublethal detrimental effect to the host (Villalba et al. 1993a, b; Fuentes et al. 1995; Pérez-Camacho et al. 1997). Therefore, the transmission of M. refringens from local M. galloprovincialis to X. securis is a plausible hypothesis. In case that the prevalence of M. refringens in the X. securis population of the Ría de Vigo reached high values, it could contribute to increase the prevalence of this disease in cultured mussels M. galloprovincialis. The lack of a thorough epizootiological study prevents from confirming if the prevalence increase in station 3 from June 2006 to 2007 corresponds to an actual increasing trend.
The occurrence of rickettsia-like organisms in digestive epithelia of cultured M. galloprovincialis is common in Galician rías (Villalba et al. 1997), although their colonies are usually located in the digestive tubules and are smaller than those observed in X. securis. Generally, no serious pathogenic effect is attributed to these prokaryotes when they infect digestive epithelia, although it could depend on the infection intensity. An exception might be the rickettsia Candidatus Xenohaliotis californiensis, responsible for the withering syndrome associated with high abalone mortalities (Moore et al. 2001).
Gonadal neoplasia has been also recorded in M. galloprovincialis collected from a raft in the Ría de Vigo, with a mean prevalence of 6% (Alonso et al. 2001). The neoplasia detected in X. securis is similar to that observed in a case of germinoma found in a mussel from the Ría do Burgo (Villalba 2006). The detection of two cases of germinoma only at station 1 might suggest an association with location. Two main causes have been proposed for cancer in molluscs: viral infection and pollution (Barber 2004).
Several analyses undertaken in the sediments of the San Simón inlet showed a very strong pollution by lead in this area (resulting values up 286 times the natural concentration for some samples), and also that signs of pollution were previously reported in the area, which indicate the ceramic factory constitutes the maximum input of lead to the Bay (Álvarez-Iglesias et al. 1999). More recently, Prego et al. (2008) showed that lead contamination in the inner part of the Ría de Vigo still remained high. On the other hand, it has been observed that there is a net Ni oceanic input (net dissolved budget is 0.5 and particulate 0.3 mmol s−1) and is accumulated inside the Ría (Prego et al. 2006). Although Ni and V biogeochemistry is still not well understood, high concentrations could be attributed to different sources, among them the influence of industrial and urban inputs from the neighbouring Vigo Ría (Santos-Echeandía et al. 2005). High loads of some contaminants (as lead, nickel and vanadium) in station 1 (Table 4) may results in the neoplasic disorders observed in X. securis. These tree elements have been reported as citogenics and mutagenics in M. galloprovincialis in the Ria de Vigo (Alonso et al. 2001).
As a whole, the pathologic conditions described in X. securis had been previously found in M. galloprovincialis from the Ría de Vigo. Remarkably, some symbionts (in the original sense of the word) that are frequent in the mussels M. galloprovincialis cultured in the Ría de Vigo (Villalba et al. 1997) and in many other bivalve molluscs, such as ciliates, copepods (Mytilicola intestinalis), trematodes, gregarines (Nematopsis spp.), turbellarians (Urastoma cyprinae, Paravortex sp.), were not detected in X. securis. The scarcity of parasite species in comparison with autochtonous mussels is consistent with the invasive character of the black pigmy mussel and could contribute to its spreading. In addition, the histological evidence of gamete production and release is consistent with the spreading of X. securis through the Ría de Vigo.
Further studies on the ecological impact of the invasive X. securis are desirables for clarifying the de facto threat this NIS pose as a pest to the Galician waters.
References
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Automat Contr 19:716–723
Alonso A, Suárez P, Álvarez C, San Juan F, Molist P (2001) Structural study of a possible neoplasia detected in Mytilus galloprovincialis collected from the Ría of Vigo (NW Spain). Dis Aquat Organ 47:73–79
Álvarez-Iglesias P, Rubio B, Vilas F (1999) A case study of pollution by lead in intertidal sediments of San Simón Bay (Inner Ria de Vigo, NW Spain). Contribution to XUGA (PGIDT99MARAE1) and CICYT (MAR 97-0626) projects. www.scientificjournals.com/sj/espr_special/Pdf/aId/3657
Bañón R, Rolán E, García-Tasende M (2008) First record of the purple dye murex Bulinus brandaris (Gastropoda: Muricidae) and a revised list of non native molluscs from Galician waters (Spain, NE Atlantic). Aquat Invasions 3(3):331–334
Barber BJ (2004) Neoplastic diseases of commercially important marine bivalves. Aquat Living Resour 17:449–466
Carlton JT (1985) Transoceanic and interoceanic dispersal of coastal marine organisms: the biology of ballast waters. Oceanogr Mar Biol Annu Rev 23:313–371
Carrasco N, López-Flores I, Alcaraz M, Furones MD, Berthe FC, Arzul I (2007) Dynamics of the parasite Marteilia refringens (Paramyxea) in Mytilus galloprovincialis and zooplankton populations in Alfacs Bay (Catalonia, Spain). Parassitology 134(Pt 11):1541–1550
Clement M, Posada D, Crandall K (2000) TCS: a computer program to estimate gene genealogies. Mol Ecol 9(10):1657–1660
Crisp DJ (1974) Factors influencing the settlement of marine invertebrate larvae. In: Grant PT, Mackie AM (eds) Chemoreception in marine organisms. Academic Press, New York, pp 177–265
DAISIE (2009) Delivering alien invasive species inventories for Europe. http://www.europe-aliens.org/
Darrigran G (2002) Potential impact of filter-feeding invaders on temperate inland freshwater environments. Biol Invasions 4:145–156
Eckman JE (1983) Hydrodynamic processes affecting benthic recruitment. Limnol Oceanogr 28:241–257
Eckman JE, Werner FE, Gross TF (1994) Modelling some effects of behaviour on larval settlement in a turbulent boundary layer. Deep-Sea Res II 41:185–208
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol 3(5):294–299
Fuentes JM, Villalba A, Zapata C, Álvarez G (1995) Effects of stock and culture environment on the infections by Marteilia refringens and Mytilicola intestinalis in the mussel Mytilus galloprovincialis cultured in Galicia (NW Spain). Dis Aquat Organ 21:221–226
Garci ME, Trigo JE, Pascual S, González AF, Rocha F, Guerra A (2007) Xenostrobus securis (Lamarck, 1819) (Mollusca: Bivalvia): first report of an introduced species in Galician waters. Aquac Int 15:19–24
Giusti F, Dell’Angelo B, Sosso M, Schiaparelli S (2008) First record of the invasive species Xenostrobus securis (Lamarck, 1819) (Bivalvia: Mytilidae) from Central Tyrrhenian Sea (Western Mediterranean). Boll Malacol 44:11–14
Gofas S, Zenetos A (2003) Exotic molluscs in the Mediterranean Basin: current status and perspectives. Ocean Mar Biol Annu Rev 41:237–277
Howard AW, Smith CS (1983) Histological techniques for marine bivalve molluscs. NOAA Technical Memorandum NMFS-F/NEC 25
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogeny. Bioinformatics 17:754–755
Kimura T, Tabe M, Shikano Y (1999) Limnoperna fortunei kikuchii Habe 1981 (Bivalvia: Mytilidae) is a synonym of Xenostrobus securis (Lamarck, 1819): introduction into Japan from Australia and/or New Zealand. Venus 58:101–117
Kleeman SN, Adlard RD (2000) Molecular detection of Marteilia sydneyi, pathogen of Sydney rock oysters. Dis Aquat Organ 40(2):137–146
Kohama T, Montani S, Kajiwara Y, Yamada M (2001) Population dynamics of sessile bivalves Mytilus galloprovincialis and Xenostrobus securis in hyper eutrophicated bay, Japan. Bull Jpn Soc Fish Sci 67:664–671
Lazzari G, Rinaldi E (1994) Alcune considerazioni sulla presenza di specie extra Mediterranee nelle lagune salmastre di Ravenna. Boll Malacol 30:195–202
Le Roux F, Audemard C, Barnaud A, Berthe F (1999) DNA Probes as potential tools for the detection of Marteilia refringens. Mar Biotechnol 1(6):588–597
López-Flores I, de la Herrán R, Garrido-Ramos MA, Navas JI, Ruiz-Rejón C, Ruiz-Rejón M (2004) The molecular diagnosis of Marteilia refringens and differentiation between Marteilia strains infecting oysters and mussels based on the rDNA IGS sequence. Parasitol 129(Pt 4):411–419
López-Flores J, Robles F, Vlencia JM, Grau A, Villalba A, De la Herrán R, Garrido-Ramos MA, Ruiz-Rejón C, Ruiz-Rejón M, Navas JI (2008) Detection of Marteilia refringens using nested PCR and in situ hybridisation in Chamelea gallina from the Balearic Islands (Spain). Dis Aquat Organ 82:79–87
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz F (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl 10(3):689–710
Medlin L, Elwood HL, Stickel S, Sogin ML (1988) The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 71:491–499
Moore JD, Robbins TT, Hedrick RP, Friedman CS (2001) Transmission of the ricketsiales-like prokaryote “Candidatus Xenohaliotis californiensis” and its role in withering syndrome of California abalone, Haliotis spp. J Shellfish Res 20:867–874
Occhipinti-Ambrogi A (2000) Biotic invasions in a Mediterranean Lagoon. Biol Invasions 2:165–176
Pérez-Camacho A, Villalba A, Beiras R, Labarta U (1997) Absorption efficiency and condition of cultured mussels (Mytilus edulis galloprovincialis Linnaeus) of Galicia (NW Spain) infected by parasites Marteilia refringens Grizel et al. and Mytilicola intestinalis Steuer. J Shellfish Res 16:77–82
Posada D (2006) ModelTest Server: a web-based tool for the statistical selection of models of nucleotide substitution online. Nucleic Acids Res 34:W700–W703
Prego R, Cotté MH, Cobelo-García A, Martin JM (2006) Trace metals in the water column of the Vigo Ria: Offshore exchange in mid-winter conditions. Estuar Coast Shelf Sci 68:289–296
Prego R, Ferro P, Trujillo C (2008) Lead and Zinc contamination of surface sediments in the main harbours of the Galician Rias. J Iber Geol 34(2):243–252
Quilez-Badia G (2009) ICES WGITMO report: 91–115. Washington DC, USA
Rozen S, Skaletsky HJ (2000) Primer3 on the WWW for general users and for biologist programmers. In: Krawetz S, Misener S (eds) Bioinformatics methods and protocols: methods in molecular biology. Humana Press, Totowa, pp 365–386
Russo P (2001) Great concentration of Xenostrobus securis (Lamarck, 1819) in different areas of the Po River Delta. La Conchiglia 298:49–50
Sabelli B, Speranza S (1994) Rinvenimento di Xenostrobus sp. (Bivalvia: Mytilidae) nella laguna di Venezia. Boll Malacol 29(9–12):311–318
Santos-Echeandía J, Prego R, Cobelo-García A (2005) Copper, nickel, and vanadium in the Western Galician shelf in early spring after the prestige catastrophe: is there seawater contamination? Anal Bioanal Chem 382(2):360–365
Snelgrove PVR (1994) Hydrodynamic enhancement of invertebrate larval settlement in microdepositional environments: colonization tray experiments in a muddy habitat. J Exp Mar Biol Ecol 176:149–166
Snelgrove PVR, Butman CA, Grassle JP (1993) Hydrodynamic enhancement of larval settlement in the bivalve Mulinia lateralis (Say) and the polychaete Capitelia sp. I In the microdepositional environments. J Exp Mar Biol Ecol 168:71–109
Sousa R, Guilhermino L, Antunes C (2005) Molluscan fauna in the freshwater tidal area of the River Minho estuary, NW of Iberina Peninsula. Int J Limnol 41:141–147
Sousa R, Antunes C, Guilhermino L (2006) Factors influencing the occurrence and distribution of Corbicula fluminea (Müller, 1774) in the River Lima estuary. Int J Limnol 42:165–171
Sousa R, Gutierrez JL, Aldridge DC (2009) Non-indigenous invasive bivalves as ecosystem engineers. Biol Invasions. doi 10.1007/s10530-009-9422-7
Streftaris N, Zenetos A, Papathanassiou E (2005) Globalisation in marine ecosystems: the story of non-indigenous marine species across European seas. Oceanogr Mar Biol 43:419–453
Swofford DL (2000) PAUP*: phylogenetic analysis using parsimony (*and other methods), version 4. Sinauer Associates, Sunderland
Templeton AR, Crandall KA, Sing CF (1992) A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 132:619–633
Turner EJ, Zimmer-Faust RK, Palmer MA, Lukenbach M, Pentcheff ND (1994) Settlement of oyster (Crassostrea virginica) larvae: effects of water flow and a water-soluble chemical clue. Limnol Oceanogr 39:1579–1593
Villalba A (2006) Different types of neoplasia in Crassostrea gigas, Venus verrucosa and Mytilus galloprovincialis from the Spanish coast. J Shellfish Res 25:786–787
Villalba A, Mourelle SG, Carballal MJ, López MC (1993a) Effects of infection by the protistan parasite Marteilia refringens on the reproduction of cultured mussels Mytilus galloprovincialis in Galicia (NW Spain). Dis Aquat Organ 17:205–213
Villalba A, Mourelle SG, López MC, Carballal MJ, Azevedo C (1993b) Marteiliasis affecting cultured mussels Mytilus galloprovincialis of Galicia (NW Spain). I. Etiology, phases of the infection, and temporal and spatial variability in prevalence. Dis Aquat Organ 16:61–72
Villalba A, Mourelle SG, Carballal SG, López C (1997) Symbionts and diseases of farmed mussels Mytilus galloprovincialis throughout the culture process in the Rías of Galicia (NW Spain). Dis Aquat Organ 31:127–139
Wallace IM, O’Sullivan O, Higgins DG, Notredame C (2006) M-Coffee: combining multiple sequence alignment methods with T-Coffee. Nucleic Acids Res 34(6):1692–1699
Wood AR, Smita A, MacAvoy ES, Gardner JPA (2007) A molecular phylogeny of the marine mussel genus Perna (Bivalvia: Mytilidae) based on nuclear (ITS1&2) and mitochondrial (COI) DNA sequences. Mol Phylogenet Evol 44:685–698
Zenetos A, Gofas S, Russo G, Templado J (2004) CIESM atlas of exotic species in the Mediterranean. In: Briand F (ed) Molluscs, vol. 3. CIESM Publishers, Monaco, 376 pp
Acknowledgments
Authors thank S. Gofas, J. Trigo, E. Rolán and E. Pulpeiro for their helpful comments on the first draft of the paper, and to JC Harshbarger for his helpful comments on gonadal neoplasia. Thanks are also due to Iain Suthers (University of New South Wales, Australia); Jacques Pelorce (Hydrologue membre de l’association du Grau-du-Roi, Camargue littoral environment, France) and G. Arcangeli (Istituto Zooprofilattico Sperimentale delle Venezie, Italy) for providing samples. J.A. Durán (IIM-CSIC), M. Meléndez, E. Penas and A. Ramilo (CIMA-Xunta de Galicia) assisted in technical analyses. Thank you to two anonymous reviewers for their excellent comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pascual, S., Villalba, A., Abollo, E. et al. The mussel Xenostrobus securis: a well-established alien invader in the Ria de Vigo (Spain, NE Atlantic). Biol Invasions 12, 2091–2103 (2010). https://doi.org/10.1007/s10530-009-9611-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-009-9611-4