
Abstract
Post-dispersal seed predation can severely limit plant recruitment, but its ultimate impact could be modulated by environmental factors and by the composition of the granivore guild. Here, we analyze the relative impact of the non-native wild boar and native rodents on seed survival and seedling establishment of the mast conifer Araucaria araucana. Predation, seed survival and seedling establishment were measured at different microsites and distances from 11 isolated trees in Lanín National Park (Argentina) over a period of marked fluctuation in seed production. Wild boar consumed between 10 and 30% of available seeds on a 13-day period, threefold less than rodents. Wild boar predation was mainly affected by forest canopy composition, while microsite conditions influenced both kind of predators, with high chronic rodent predation underneath dense vegetation and moderate (but interannually variable) wild boar predation at poorly vegetated microsites. Predation by rodents was spatially clustered at the microsite scale, particularly during non-mast years; while predation by wild boar was spatially structured at a coarser scale and less modified by masting. The exclusion of wild boar increased significantly the amount of surviving seeds, resulting in higher seedling establishment in intermediate production years, but not affecting it during the mast year. At tree level, seedling establishment was negatively correlated with predation; while at stand level, cone production accounted for most of the seedling establishment variability. The current wild boar population may not be affecting the seedling establishment at population scale, probably due to minimization of its impact by the Araucaria masting strategy. However, if wild boar population numbers continue to increase, their impact may shift from individual tree scale to stand scale, threatening Araucaria forest regeneration.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Exotic species are considered one of the main factors of biodiversity alteration (Vitousek et al. 1997) by modifying community’s composition, food webs and ecological processes (Mooney et al. 2005; Ruscoe et al. 2006). The invasion of exotic herbivore species may alter the native granivory assemblage and their interactions with plants (Moles and Drake 1999; Choquenot and Ruscoe 2000; Wilson et al. 2007) and may ultimately modify the relative importance of granivory and/or herbivory compared with the other forces that control seedling establishment (Allen et al. 1994; Campbell and Atkinson 2002; Wilson et al. 2003).
At a local scale, the population size of a species with large edible seeds may be limited either by habitat carrying capacity or by intrinsic ability of populations to grow. In the former case, the limitation is the microsite and population size would change only if the habitat structure changed by disturbance, independent of the amount of seeds produced or surviving. In the latter case, the population size may be regulated by seed availability (seed limitation) and/or by germination success and seedling survival (recruitment limitation; Clark et al. 1999). Therefore, the ultimate effect of an exotic species will depend on which stage of the plant demography it affects and the strength and extent of this effect in terms of the spatial and temporal scale from the plant perspective.
The impact of granivory by exotic species on seed survivorship on long-lived trees may be minimized, if the plant is a mast seeding (intermittent synchronous output of seeds) species and if this evolutionary adaptation is associated with predator satiation, allowing a higher probability of seedling recruitment after the peak seed production (Predator Satiation Hypothesis: Janzen 1971; Silvertown 1980; Ostfeld et al. 1996). On the other hand, the environment may greatly influence the effect of the exotic predator on plant demography, if herbivory occurs mainly on microsites where plant reproductive fitness is normally maximized due to higher seed or seedling survivorship and establishment rates. For example, the exotic species may affect the abundance and/or distribution of recruits away from the parent if it attacks mainly the sites that are “safe” against native predators (Gómez et al. 2003; Iida 2004). Environmental influence can also be seen if microsites favor predation in the area of higher primary seed dispersion, where the exotic species may ultimately modify the initial seed rain pattern produced by wind dispersal (Nathan and Casagrandi 2004). Few studies considered these two aspects of plant ecology in the context of biological invasions and the consequences on plant recruitment (Choquenot and Ruscoe 2000; Sweetapple 2003; Wilson et al. 2007).
Related to the worldwide European cultural expansion, wild boar (Sus scrofa) has established in every continent and in several oceanic islands (Lever 1985). Because it is an r-strategist with high ecological plasticity, high opportunistic feeding behavior and the highest reproductive potential of all ungulates in relation to body mass (Rosell et al. 2001), this introduction followed by invasion has become of great concern among conservation biologists and is now considered one of biggest threats to biodiversity conservation (Singer et al. 1981; Tisdell 1984; Lowe et al. 2004; Cruz et al. 2005; Silva and Saavedra 2008).
In all the places where wild boar has been introduced (Australia, New Zealand, United States, Hawaii, Galapagos islands, Chile and Argentina) negative impacts to the ecosystem like changes on species composition, local extinctions of plants, reduction of species diversity and promotion of exotic plant invasion by soil disturbance were reported (Howard 1964; Bratton 1975; Challies 1975; Wood and Roark 1980; Howe et al. 1981; Singer et al. 1981; Campbell and Rudge 1984; Tisdell 1984; Coblentz and Baber 1987; Vtorov 1993; Sweitzer and Van Vuren 2002; Merino and Carpinetti 2003; Cruz et al. 2005; Skewes et al. 2007).
In its natural range in Europe and Asia, wild boar is associated with large-seeded species (Quercus spp. and dipterocarp forests) where the ungulate predates on seeds and seedlings (Groot Bruinderink and Hazebroek 1996; Ickes et al. 2001; Rosell et al. 2001; Pulido 2002). In this scenario, the ultimate effect of wild boar on seedling establishment and recruitment depends on the masting characteristics of the trees, on the understory vegetation structure and on the animal numerical response to food pulses occurrences and to snowy winters or summer droughts (Massei et al. 1997; Curran and Leighton 2000; Curran and Webb 2000; Kuiters and Slim 2002; Maillard and Fournier 2004; Bieber and Ruf 2005; Melis et al. 2006).
In oak and dipterocarp forests, the rodent assemblage consumes large amounts of seeds and is influenced by masting events and climate conditions (Jensen 1982; Wolff 1996), but compared with wild boar, the interaction probably differs in terms of the spatial and temporal scale due to differences in movement capacities and home range sizes (McShea 2000; Sodeikat and Pohlmeyer 2001). Resident and less mobile species, like rodents, might increase seed predation rates by changing their diet and/or through reproduction within the seed fall season (Hulme and Hunt 1999). In contrast, large nomadic vertebrates like wild boar, acting at a different spatial scale, may perceive the seed as a patchy-distributed resource that is seasonally available, so the expected response could be a change in their diet while they migrate (Curran and Leighton 2000; Silman et al. 2003).
We studied the temporal and spatial patterns of post-dispersal seed predation by rodents and non-native wild boar (S. scrofa) on a mast species, the large-seeded conifer Araucaria araucana. In addition, we assessed the relative effects of the different granivores on seedling establishment considering the influence of masting, the distance from the source and vegetation heterogeneity. In this context, we discuss the influence of wild boar predation on an ecosystem without this type of herbivore and its relative importance compared with the other factors that limit A. araucana regeneration. To conduct this study, we posed the following questions: (a) do rodents and wild boars have similar quantitative, temporal and spatial seed predation patterns?; (b) does seed predation by rodents and wild boars differ according to masting, vegetation cover or distance to the source?; (c) is there evidence of seedling establishment limitation by wild boar seed predation?
Materials and methods
Study species
Araucaria araucana (Araucariaceae) is a long-lived (>1,200 years), large seeded dioecious emergent conifer found in the temperate forest of South America (Veblen et al. 1995). Female cones contain 100–200 large nuts (piñones) weighting ~3.5 g, which are dispersed by gravity only up to 13 m from the parent tree (Muñoz Ibañez 1984). Seeds start to fall in March until June and exceptionally, September. A. araucana is a masting species with a low average fecundity (960 seeds per tree) and a moderate temporal variation in cone production that fluctuates annually between 2 and 45 mean cones per tree with a highly synchronized production among trees within population and among populations at a regional scale (Sanguinetti and Kitzberger 2008). Main native post-dispersal seed predators are the greater long-clawed mouse (Chelemys macronyx), the long haired mouse (Abrotrix longipilis), the long-tailed mouse (Oligoryzomis longicaudatus) and the arboreal mouse (Irenomys tarsalis) (Shepherd and Ditgen 2005). According to Shepherd and Ditgen (2008), A. longipilis may play a relevant role as a disperser of Araucaria seeds to favorable microsites for seedling germination. An exotic predator, the wild boar, invaded the forests in our study area during the 1970s, from east to west (Jaksic et al. 2002). A dietary study shows that, in autumn, wild boar feces contain more than 90% of Araucaria seeds (Izquierdo et al. 2001).
Study area
This study was conducted from 2004 to 2008 in a 250 ha A. araucana and Nothofagus pumilio or Nothofagus antarctica mixed patch in Lanín National Park, Neuquén Province, Argentina (39°34′44″S/71°27′39″W) with an average winter temperature of 7–8°C (De Fina 1972). The selected patch was the largest and densest, and was 2–19 km apart from the other parches. In xeric sites N. antarctica shrubs (<5 m height) dominate the understory and gaps within the forest. Tall grasses are the predominant vegetation in moderate shade to sunny sites. Beneath A. araucana trees, there is less vegetation cover, such as short forbs (<0.5 m in height), Araucaria seedling clumps, and dead leaves or bare soil, depending on soil type and amount of sunlight. Forest gaps in humid places are occupied by dense high pastures or dense bamboo Chusquea culeou thickets.
Experimental studies
Seed predation and the influence of environmental factors
Within 30 ha of the selected patch, we randomly chose and marked 11 female A. araucana trees distant at least 30 m from the nearest female tree. We studied seed predation by rodents and wild boar and the effects of masting, habitat, microsites and distance from parent tree from 2004 to 2008 (Table 1). Seeds were placed along two radial transects per marked tree, in groups of 20 seeds, at 5, 10, 15, 20 and 25 m from the tree base. Each of the trees and the seed groups were geographically located with GPS for spatial analyses. Seed predation was estimated by considering the proportion of seeds removed on three consecutives nights from the total seeds per group or per marked tree, depending on the analysis performed (Hulme and Hunt 1999). In 2004, 2006, 2007 and 2008, between March and June, we also performed the same study but over periods of 13 consecutive nights (hereafter called “fortnight”), repeating it consecutively after replacing the predated seeds, when possible. In 2004 and 2007, we performed six consecutive fortnight samplings. In 2006, only two fortnight samples were obtained between April and May and in 2008, we measured seed predation only once during May–June, when the highest seed consumption usually occurred, because of the great scarcity of seeds. In 2005, it was impossible to do the fortnight sampling altogether, because of the extremely poor cone productivity (mean value of two cones per tree) which hindered us to obtain sufficient seeds for the experiment. Habitats were defined according to canopy composition: A. araucana and N. pumilio mixed forest and A. araucana and N. antarctica mixed forest. Microsites were classified as either: bare soil (N = 15), leaf litter (N = 20), forbs or Araucaria seedling clumps (N = 18), tall grasses (N = 23), shrubs or bamboo thickets (N = 34), representing an increasing gradient of vegetation cover. Levels of vegetation cover were defined as “low” (pooling bare soil and leaf litter, N = 35), “medium” (forbs and tall grasses, N = 41) and “high” (shrubs and bamboo thickets, N = 34). The groups of marked seeds at the fixed distances were placed randomly within microsite types.
In order to differentiate the predation produced by each type of predator, we obtained information from colleagues that concurrently monitored the predator’s activities with automatic cameras in 2004 and 2005 at different microsites during the whole seed fall season (Shepherd and Ditgen 2005; 2008). Using 10 cameras at different locations within the Araucaria study forest, more than 1,700 photos of granivores were used to identify seed predator species and the way they fed and damaged the seeds. Rodents, wild boar and, less frequently, hare fed on untouched seeds on the ground. Rodents may act as scatter-hoarders and mainly remove the marked seeds, but when they feed in situ they leave teeth marks on the seed. In contrast, wild boar finds the seeds during rooting activity and destroys them completely; leaving flattened empty seed covers.
Masting behavior was assessed using cone counts per seeding tree obtained from our related study on cone production at tree and forest level in the same trees and study area where seed predation was evaluated (Sanguinetti and Kitzberger 2008). Mean cone production per tree was 23, 1, 12, 35 and 7 for 2004, 2005, 2006, 2007 and 2008 year, respectively.
Variation of predation within the seed fall season
Data obtained between March and June in 2004 and 2007 with the fortnight seed predation repeated measures were used to analyze seed consumption variation within the seed fall season and the predator response during a moderate and high cone production year, respectively. In 2004, 2006 and 2007, we monitored in November the fate and abundance of marked seeds that remained after the last fortnight sampling of the season (Table 1).
Seed predation and seedling establishment
Between 2004 and 2008, we measured Araucaria seedling establishment around the 11 selected trees where seed predation experiments were performed (Table 1). Using eight radial transects of 20 m per tree, we estimated the abundance of seedlings per square meter. The year of establishment was estimated through seedling height, due to the strong relationship between this variable and age (F (1,70) = 517.2; R 2 = 0.881; P < 0.0001; Sanguinetti, unpublished data). The seedling establishment accumulation over the period 2004–2008 was compared with the estimated seed predation rates of rodents, wild boar or both at tree level.
Effect of wild boar exclusion on seed survival and seedling establishment
In 2004, we constructed two 2,500 m2 exclosures against large granivores, each encircling five seeding trees. Between 2004 and 2007, we used eight radial transects of 15 m per tree in the exclosures to estimate abundance of seeds and seedlings per square meter, and we used 5 of the 11 initially marked trees as control for each exclosure (Table 1). At the same time, we counted the number of cone scales to obtain an estimation of the total seed fallen per plot, considering the strong correlation between seeds and scales inside the cone (F (1,18) = 17.1; R 2 = 0.487; P < 0.001; Sanguinetti, unpublished data). This allowed us to estimate the seed survivorship variation with distance from the source, inside and outside the exclosure. We also estimated the year of establishment of seedlings over a period of 3 years before and 3 years after exclosures construction.
Changes in predator relative abundance
Between 2003 and 2007, the relative abundance and the forest use by wild boar were monitored by recording the proportion of ground uprooted by boars (Hone 1995). Six parallel, permanent transects (300 m long and 4 m wide), separated from one another by 100 m, were established to cover the entire area where the seed predation was estimated. The annual mean value of the number of rootings and their size per transect were used as an index of changes in wild boar relative abundance and forest use. Mean rodent relative abundance data from 2003 to 2007 were obtained from a related study carried out in the same forest. An increment between 1.8- and 3.5-fold was observed from February to April in rodent relative abundance for all the years during the period 2004–2007. The years with the highest and lowest rodent numerical responses during the fall seed season were 2004 and 2007, respectively. In April, the mean rodent relative abundances (captures per 100 trap nights; N = 500 trap nights) were 1.38, 0.77, 0.75 and 1.04 for 2004, 2005, 2006 and 2007, respectively (Shepherd and Ditgen, unpublished data, 2008).
Data analyses
Seed predation and the influence of environmental factors
Seed predation rates were analyzed using general linear mixed models using Statistica 6.0 software. The proportion of eaten and/or removed seeds was root arcsine transformed and tested for normality using Kolmogorov Smirnov test before analysis with ANOVA and MANOVA repeated measures were performed. Seed predation by rodents and wild boar at 3 and 13 days, the dependent variables, were performed separated or together by univariate or multivariate analysis, respectively. In the 13-day experiment, each seed group was considered the subject and fortnights and years as the repeated measures (two within-subject levels), distance from parent tree as covariate fixed effect, and habitat, microsites or vegetation cover were considered the between-subject random factors (Sokal and Rohlf 1994). Six repeated measures were used when we compared years 2004 and 2007 and only data from the 3-day experiment was used to analyze the among years variation on seed predation during 2004 to 2008. For this analysis, the natural logarithm of the number of cones per tree was used as a covariable to evaluate the masting effect on seed predation. The effect of distance from the parent on seed survivorship was explored by regressions with the Cox’s Proportional Hazard semiparametric model, using the maximum partial likelihood as estimation method (Scheiner and Gurevitch 2001).
Within seed fall season variation of predation
Seed predation variation within seed fall season was evaluated using general linear models (GLM) with repeated measures. Data from 2004 and 2007 were used from six repeated measures covering almost the entire seed fall season.
Spatial variation of seed predation
To verify potential variations on the spatial pattern of seed predation by rodents and wild boar, Moran’s I spatial autocorrelation statistic was used (Moran 1950). Point-referenced seed groups (x and y coordinates) and seed exploitation data by predator identity were used to estimate the Moran Index and the correlogram with different lag distances. Mean I values for each predator were obtained from the largest data set performed by the repeated measures sampling design during 2004 and 2007. Significance test was performed for complete spatial randomness (Moran 1950). Significantly positive Moran’s I values at a certain lag distance mean that seed predation rates from points separated by that distance are more similar than those farther apart, denoting a “contagious” or “aggregated” effect. On the other hand, significantly negative values for a particular lag distance mean that seeds separated by that distance suffered different seed predation rates denoting the existence of a “repulsive” effect at that spatial scale. To test for different spatial patterns of predation among predators, we performed a partial-Mantel test using the distance matrices for seed exploitation by each predator and a third matrix, holding constant, of distances between seed locations. Significance was determined by a permutation test and a t-test.
Seed predation and seedling establishment
Annual estimates of seed predation rates by each type of predator and seedling establishment per tree were used in regressions analysis. Seed predation by each type of granivore or the overall predation rates were considered as predictor variables, and the density of seedling established per tree and year were used as the dependent variable.
Effect of wild boar exclusion on seed survival and seedling establishment
The effect of wild boar exclusion on seed survival was analyzed by generalized linear models (GLMs) using the number of seeds per plot and transect of 10 seeding trees inside and outside the exclosures as the dependent variable; the treatment as a factor; and the distance to the tree as a covariate, using a log function as a link function. On the other hand, the ratio seeds:scales per plot was used as a seed survivorship index and a Homogeneity-slope model test was applied to verify significant differences on seed survival with distance between trees inside and outside the exclosures.
Seedling establishment variation was analyzed using Kruskal Wallis test considering the number of seedlings per tree as the dependent variable and the treatment as a factor. For each year, a Mann–Whitney test was performed to verify the significance level of seedling abundance per tree affected by the treatment.
Changes in predator relative abundance
An analysis of variance was performed to evaluate the existence of significant differences between years of seed predator relative abundance and forest use. Before the analysis was done, the number and size of rooting per transect and the proportion of the ground covered by rooting were natural log and root arcsine transformed, respectively. Information about within season variation on relative abundance of rodents provided by Shepherd and Ditgen was used to interpret the temporal variation on seed predation rates.
Results
Seed predation and the influence of environmental factors
Masting effect
In general, wild boar consumed 3–6 times less seeds than rodents, accounting for 10–30% of overall granivory (Fig. 1). Seed predation by native and exotic species had a significant different temporal pattern among years (Table 1; MANOVA test) where both kinds of granivores increased seed consumption during non-mast years, but rodents had a strong temporal variation (Fig. 1). Only native species had a significantly negative relationship between seed predation and seed production [ANCOVA effect: F (1,50) = 8.43; P = 0.005 (native) vs. F (1,50) = 0.52; P = 0.47 (exotic)].

Mean (±SE, N = 11) A. araucana seed predation by rodent, wild boar or both predators per tree during the period 2004–2008 estimated from 3-day seed disappearance during May. For each data set different letters means significant differences (P < 0.01). Arrows represents masting condition: inter-mast (continue) and mast (discontinue)
Habitat and microsite effects
Habitat type was an important variable producing different ratios of seed predation by exotic and native granivores (MANOVA: Habitat: F (2,104) = 11.09; P < 0.0001) principally due to the strong habitat selectivity of wild boar, which positively selects mixed Araucaria–N. pumilio over Araucaria–N. antarctica forests (Tables 1, 2). Only seed predation by native species exhibited temporal variation on habitat effect (Table 1) due to a stronger decrease in N. pumilio habitat during mast year.
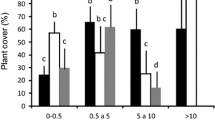
Microsites explained much of the variation in seed predation by native and exotic species and their overall predation intensity (Tables 1, 2). Seeds in C. culeou bamboo thickets had a significantly higher predation than all other microsites (post hoc Bonferroni test, P < 0.0001). Predation underneath shrubs was significantly higher than in leaf litter and bare soil microsites (P < 0.05). In bamboo and shrubs, rodents were the main predators whereas wild boar consumed significantly more seeds in high pastures, dead leaves and short shrubs microsites (P < 0.0001). Therefore, vegetation cover effect explained the differences on seed predation, where seed predation was significantly higher at dense vegetation microsites (F (2,106) = 28.18; P < 0.0001); no differences were found among microsites with medium and low cover (P = 0.163). Wild boar concentrated their predation on low or moderate vegetation cover while rodents consumed more seed in high cover microsites.
Species-specific differences in predation and survival rates were explained by predation at high vegetation cover irrespective of the year or at low vegetation cover only during the non-mast year also (e.g., 2004)(Fig. 2, MANOVA test; Table 2). During mast years, low cover microsites received similar low predation levels by both granivore types. These differences among native and exotic species between masting and no masting years were mostly explained by an intra-seasonal behavior predation within high cover patches (Fig. 2; MANOVA: Year × Fortnight × Cover: F (20,192) = 2.53; P < 0.001).

Vegetation cover and within year effects on mean seed predation (proportion) by rodent, wild boar and both (overall) predators estimated by fortnight repeated measures: a 2004 year; b 2007 year. Bars denoted confidence intervals with α = 0.05
Distance effects
Distance to seed source was a poor linear predictor of Araucaria seed predation rates. Only when microsites were considered, did distance explain a significant part of the variation in seed predation rates. A consistent pattern of maximum seed survivorship at intermediate distances (15 m) emerged when high cover microsites were excluded from the analyses; regardless of year and time within the seed fall season. A significant interaction was observed between predator identity and distance, where seed survival from rodents showed a positive relationship with distance (proportional Cox-regression analysis: χ2 = 67.9, P < 0.0001, b = 0.023 ± 0.002) and seed survival from wild boar showed a negative relationship with distance (proportional Cox-regression analysis: χ2 = 64.4; P < 0.0001, b = −0.030 ± 0.003).
Within seed fall season predation
Intra-seasonal patterns of predation changed between native and exotic granivores and according to the masting condition. During the non-mast year 2004, predation by rodents increased early within the seed fall season and remained high during the mid-late season, whereas predation by wild boar increased toward the end of the seed fall season. In contrast, during the masting year 2007, predation by rodents increased steadily while wild boar predation decreased slightly with time (Fig. 2; MANOVA: Fortnight × Year: F (10,96) = 4.03; P < 0.0001). Therefore, within-season seed predation by both exotic and native almost doubled within the seed fall season during non-mast year of 2004, but this trend was dampened from May to June during strong mast year 2007 (Year × Fortnight, Table 2). In consequence, between March and June a higher proportion of seeds survived to overall seed predation during 2007. However, during winter and early spring in years with enough production of seeds, predation by wild boar doubled the rate by rodents; and ate 65% of the remaining seeds.
Spatial variation in seed predation
Spatial patterns of seed predation showed marked differences between native and exotic species and years, with exceptions at very short distances (5 m) and very long distances (>250 m). At short distances, both predators showed high positive spatial autocorrelation, but at long distances both granivores showed lack of autocorrelation, irrespective of the year of observation. Predation by rodents showed a significant positive autocorrelation within trees contrasting with spatially independent predation patterns in wild boar, and an opposite pattern at distance between neighboring trees where the exotic showed a significant negative autocorrelation (Fig. 3). The clumping in rodent granivory was enhanced during the non-mast year of 2004, compared to the mast year of 2007. In contrast, wild boar did not substantially change the spatial pattern of seed predation between these years. However, partial-Mantel tests show that both predators had a significantly high correlation value between seed predation patterns (0.784) (t = 19.93; P < 0.001) that significantly departed from randomness (P = 0.001).

Correlograms of rodent and wild boar seed predation during 2004 and 2007. The mean Moran’s I Index from the repeated measures sampling experiments are shown. Distance is in log scale to clarify the graph and the different scales levels are shown below. Asterisk shows significantly autocorrelation values at α = 0.05
Seed predation and seedling establishment
During 2004–2008, a negative but not significant relationship was observed between seedling establishment and seed predation rates at tree level (Fig. 4). This negative relationship held true when seed predation was partitioned into native and exotic species, but was stronger for wild boar (Fig. 4; Table 4). At tree level, no significant influence of cone production on seedling establishment was observed. However, at stand level, cone production accounted for most of the seedling establishment variability, while overall predation and wild boar predation became marginally significant (Table 4).

Relationship between seedling establishment and mean seed predation rates by wild boar, rodents and both predators per tree during the period 2004–2008
Effect of wild boar exclusion on seed survival and seedling establishment
Wild boar exclusion produced a significant variation on seed survivorship only in the spring of mast year 2007, when a fourfold increase was observed (Wald χ2 = 135.5, P < 0.0001). No seeds were detected in 2005 and no significant differences were observed between treatment conditions in 2006 (Wald χ2 = 0.045, P = 0.831; Fig. 5a). The seed survivorship index (ratio seeds:scales) significantly changed in mast year 2007 where wild boar was excluded, and a clear negative relationship between seed survivorship and distance was observed (Fig. 5b). At tree level, seedling establishment accumulation of 4 years significantly increased twofold inside the wild boar exclosures (0.23 ± 0.06 vs. 0.11 ± 0.03 seedlings m−2; K–W test: H (1,18) = 14.06, P = 0.0002) compared to control plots. The effect of wild boar exclusion on seedling establishment was stronger during inter-mast years, when higher differences were observed between treatment conditions (Fig. 5c), but was non significant during the mast year 2007 (Mann–Whitney test: U (1,9) = 27.0, P = 0.233).

Wild boar exclusion effects on seed survivorship and seedling establishment at tree level (N = 9): a seed density (±SE); b survivorship index variation around trees and; c seedling density establishment (±SE) before and after exclosure building and inside and outside them. * P < 0.05, ** P < 0.01. Dashed and continuous arrows indicate date of exclosure building and mast year, respectively
Changes in predator relative abundance
Wild boar rooting numbers, size and percent cover on the forest floor changed significantly among years (F (4,20) = 4,94, P = 0.006, F (4,20) = 9,3, P = 0.0002, F (4,20) = 3,02, P = 0.04; respectively). Wild boar rooting activity was higher during 2007, 2004 and 2003 with 9.1 (±1.9 SE), 9.0 (±1.4 SE) and 8.9 (±3.8 SE) rootings per transect. In contrast, rooting numbers in 2005 and 2006 were 5.5 (±1.4 SE) and 3.2 (±0.8 SE), respectively, suggesting a lower wild boar activity in the forest during this period. Similar results where observed with percent cover of rooting in the forest floor and with rooting sizes: on average, sizes varied between 22 and 102 m2 during the higher use years and only 6.2 m2 in 2006. Rodent relative abundance increased significantly in mid-April during 2004, 2006 and 2007 (Shepherd and Ditgen, personal communication), coinciding with the fortnight period when seed predation by rodents increased.
Discussion
Native and exotic species had different relative impacts on post-dispersal seed predation in A. araucana forest, according to the amount of seeds consumed, their temporal and spatial variation and their responses to vegetation heterogeneity. In our study area, although rodents were the main seed eaters, introduced S. scrofa consumed an important proportion of the total seeds, especially outside dense understory vegetation and close to seeding trees, where wild boar predation was higher than 30%, equivalent to that of rodents.
Wild boar had a lower predation fluctuation between years and a lower response to strong inter-mast and mast events and showed a broader predation activity within season, extending their seed consumption during winter and spring becoming at that moment the main predator. In our study area, wild boar reduced significantly the amount of surviving seeds during mast year but this effect did not alter seedling establishment, suggesting that other factors related with recruitment limitation may be acting on this conifer. However, this recruitment limitation is not related with this exotic species because, contrary to what happens in Quercus spp. forest, wild boars do not eat or destroy Araucaria seedlings during rooting activities. These results suggest that during inter-mast years A. araucana seedling establishment is controlled by seed availability (seed production and/or surviving limitation) where a reduction on seed predation pressure by wild boar exclusion may allow a higher seedling establishment. This non-native ungulate may only be affecting A. araucana seed survivorship and seedling regeneration at the individual level but may not affect the tree population as a whole. Consequently, the masting pattern of this conifer, characterized by the high synchrony between trees and populations, may alternatively result in the starvation and the satiation of native and exotic predators in order to guarantee that a minimal threshold of seeds survive to overcome the next filter to population recruitment that affects seedling survival and growth. However, although wild boar affected seed survivorship at the study site, considering the small sample size these results cannot yet be generalized and more studies are needed to determine the real extent of wild boar impact on Araucaria forests.
Temporal variation and masting effect
The significant inverse relationship between cone productivity and seed predation levels during the period 2004–2007 at the population level, together with important increases in seed survival and seedling establishment during mast year, support the Predator Satiation Hypothesis (Janzen 1971; Silvertown 1980). Similar results were observed for mast seeding tree species that interact with resident and mobile predators and where the high spatial synchrony between trees and populations was considered an important factor for seed satiation (Jensen 1982; McShea 2000). Seed predation in A. araucana may depend not only on the crop size during a particular year, but also on the size of the previous crop. The two highest seed predation rates were observed during 2005 and 2008, after years with moderate to high crop sizes. On the other hand, during 2006 and 2007 we observed the lowest predation rates, after years with poor crop sizes. This interannual predation pattern was observed on mast large-seeded species with wild boar and rodents as the main predators (Singer et al. 1981; Ickes et al. 2001; Rosell et al. 2001).
Our estimates of seed predation by wild boar is 15-fold greater than the values reported for the species on oaks (Quercus spp.) in Europe, where the species is native and consumes one seed for every 99 eaten by rodents (Gómez et al. 2003). According to Uzal and Nores (2004), wild boar populations are controlled by a complex negative feedback system with a primary factor acting with a one year delay (the unstable acorn and beech food supply caused by mast fluctuation) and a secondary factor with 2 years of delay related with weather conditions or with the time lag of a cohort to reach reproductive status. Several empirical datasets reveal the importance of food pulses provided by large-seeded trees on wild boar birth synchrony (Maillard and Fournier 2004) and on population dynamics (Jedrezejewska et al. 1997; Bieber and Ruf 2005). Other studies concluded that the severity of the winter was the main force controlling wild boar population dynamics (Melis et al. 2006). In our system, the severe winter and low seed production of 2005 could have determined both the low Araucaria forest use by wild boar in 2006 and the lack of an increase in seed predation during the mast event in 2007. This relationship between forest use and seed consumption variation by the introduced wild boar and masting was observed in Quercus spp. forests in USA, where wild boar decrease significantly the forest use during inter-mast years and where the volume of the stomach filled by acorns changed from <10 to 84% from inter-mast to mast years (Singer et al. 1981).
Rodents were the main seed predators from March to June, and, according to trap data (Shepherd and Ditgen, unpublished data, 2008), increased seed predation during the fall seed seasons was caused by numerical responses of the rodent population. Lower rodent densities were observed during the lowest inter-mast years but this does not prevent the occurence of the highest seed predation rates. During mast years, trap data suggest that rodents may not build-up their numerical response enough to increase seed predation. Wild boars became more preponderant from June onward and the fact that they started consuming A. araucana seeds at the peak of their availability, together with the difference in use intensity among years, may suggest that they could be maximizing food intake and minimizing travel and search costs by using the highest quality habitat during peak seed availability as was observed in USA (Singer et al. 1981). This preliminary conclusion is supported by the fact that the studied Araucaria forest seems to be the patch most used by wild boars in autumn. During 2008, no seed predation by wild boar was detected on the neighboring patches (Sanguinetti, unpublished data, 2008). However, more studies are necessary to understand how wild boars respond to spatial heterogeneity in terms of habitat and seasonal variations in food availability. One of the issues to consider in the future should be seed predation during spring, when wild boar could be feeding on seeds dispersed by rodents, as was observed in Europe with Quercus spp. (Focardi et al. 2000). If this was the case, wild boar could affect the potential function of A. longipilis as secondary disperser.
Habitat and microsite effects
Seed predation by rodents and wild boar was strongly affected by vegetation heterogeneity at coarse and fine spatial scales, but these interactions changed within the seed fall season and among years. Our results suggest that wild boar feeding decisions may occur first at a coarse vegetation scale defined by canopy composition (Singer et al. 1981) and, within this habitat, predation would be regulated by a finer scale associated with open understory with few vegetation cover. On the other hand, rodents are more regulated by fine scale heterogeneity related with high understory vegetation cover (Caccia et al. 2006). Wild boar seems to be consuming seeds at “safe” places against rodent predation, especially during mast years, when rodents leave mostly vacant that type of environment, due to the enough resource available under denser vegetation. The existence of high seed predation by rodents in areas with bamboo, regardless the year, suggest that A. araucana seedling regeneration may be strongly hampered by the indirect effect of bamboo on seed survivorship by providing optimum habitat for their main predators, as was observed elsewhere (Wada 1993).
Distance effects
Rodents and wild boar showed an opposite feeding pattern. Rodents tended to consume seeds farther away from the parent trees, where vegetation cover is denser, while wild boars preferred lower cover microsites, more commonly found next to the parent tree. This explains the raise in seed survivorship at intermediate distances. The strong change in the distribution of surviving seeds around the seeding tree with wild boar exclusion seems to confirm the distance-dependent seed predation by this exotic species.
Seed predation spatial pattern by rodents and wild boar
The significant differences on seed predation pattern between rodents and wild boar may be related to their differences in feeding strategies, movement limitations and home range sizes. Wild boars usually move seasonally along 3.6–12.3 km2 of their home range for food (Singer et al. 1981; Rosell et al. 2001). Therefore, they are able to select particular places for feeding according to food abundance. In our study area, wild boars use the Araucaria forest during the seed fall season like was observed in North American oak forests (Singer et al. 1981). Within the forest, the significant negative autocorrelation pattern in wild boar seed predation at neighboring trees scale may be related to the selection of certain mother trees located on humid sites with a specific vegetation cover. The existence of a positive autocorrelation only at very short distances within trees may be the consequence, according to the distance to the source pattern observed, of a higher seed predation occurring in a small area close to the selected seeding tree where wild boars probably spend more time searching for food. On the other hand, rodents have a small home range, usually less than 1800 m2 (Redford and Eisenberg 1992) and often with a size similar to the seed rain area of the A. araucana tree (530–700 m2); therefore, their predation pattern will be more related with their ability to increase caching and/or to produce a numerical response at the tree scale. The spatial autocorrelation of rodents with a significant increment within tree scale during the inter-mast year may suggest that seed predation is controlled by the abundance of seeds within the vegetated sites and by the fact that rodents take more risk venturing on poor vegetated sites during low cone production years. The spatial autocorrelation analysis together with the microsite effects on seed predation coincide and support the conclusion that during inter-mast years rodents move all around the seed rain area searching for seeds at all types of microsites, while on a mast year they restrict their movements to vegetated sites with enough food. These differences between rodents and wild boar on seed predation spatial patterns may affect the spatial pattern of seedling establishment. The results of the regression analysis performed at tree and stand scales suggest that seedling establishment is related with seed predation by rodents at tree scale and by wild boar at tree and stand scales.
Short and long term effects of seed predation on A. araucana regeneration
Our estimates of predation rates and seed survivorship, the results of the multiple regression analysis over seedling density per tree for the period 2000–2007 and the effect of wild boar exclusion suggest that rodents and wild boar strongly limit the availability of seed for germination during inter-mast years, but not during mast events, where neither the native nor the exotic species apparently inhibit seedling establishment. This conclusion is supported by the fact that at stand scale, seedling establishment depends mostly on the number of seeds produced. During the mast year 2007, similar number of seedlings were counted inside and outside the exclosure regardless the significantly higher number of seeds surviving without wild boar. This unexpected result may indicate that more seeds survive outside the exclosure between November and February and/or germination is facilitated by wild boar disturbance, both possibly due to the covering of seeds with soil during rooting activity. Although wild boar seed predation is high and occurs mainly on microsites that are “safe” against rodent predation and favorable for seedling establishment, the rooting disturbance may compensate this negative effect allowing the surviving seeds to reach the seedling stage.
In general, we may conclude that the spatial and temporal pattern of seedling establishment in A. araucana is probably determined by seed, microsite and recruitment limitation factors acting simultaneously. Temporally, seedling establishment is limited by cone production and modulated by seed predation and microsites. In this context, the current wild boar population is affecting the favorable microsites for Araucaria seed germination and seedling establishment, but without depleting all the available seeds; moreover, it is not affecting seedling growth and survival, due to Araucaria large roots and tough and unpalatable leaves. These could be one of the main differences between Quercus spp. and Araucaria forest since in the former wild boar not only consumes a lot of seeds but also eats and uproots many seedlings (Groot Bruinderink and Hazebroek 1996; Sweitzer and Van Vuren 2002; Gómez et al. 2003).
Therefore, for our study area, we preliminarily predict the existence of pulses of seedling establishment during the summer after the occurrence of mast years. However, seed predation by wild boar may produce an important impact on the individual fitness of mother trees by reducing the abundance of seedlings and/or modifying their spatial distribution. We predict that trees surrounded by a high proportion of open vegetation in humid places will be more affected by wild boar predation, and their seedlings should establish at intermediate distances from the parent tree; whereas trees with a low seed production potential and surrounded by a high proportion of dense vegetation will be more affected in their regeneration success by rodent predation, and their rare seedlings should be expected to occur near the parent. Highest levels of seedling regeneration are expected to occur in places with a high density of mother trees without bamboo or shrub understory, and beneath the biggest trees, that can compensate for the higher seed predation rates through a disproportional cone output during a mast year.
This study suggests that the current wild boar population, which invaded the Araucaria study area some 30 years ago, may not be affecting the seedling establishment, probably due to minimization of its impact by the Araucaria masting strategy. However, wild boar can extraordinarily increase its population numbers, as was seen in Europe due to surplus food and milder winters (Rosell et al. 2001). If in north Patagonia wild boar numbers continue to increase, which is very likely (Peris et al. 2007), their impact may shift from individual tree scale to stand scale, threatening Araucaria forest regeneration. Although this threat could take several centuries to be noticed considering Araucaria longevity, climate change may favor wild boar fitness and may also cause a higher frequency of forest fires, which may turn into a much more threatening scenario for Araucaria sooner than expected.
References
Allen R et al (1994) Regeneration in indigenous forest after eradication of Norway rats, Breaksea Island, New Zealand. N Z J Bot 32:429–439
Bieber C, Ruf T (2005) Population dynamics in wild boar Sus scrofa: ecology, elasticity of growth rate and implications for the management of pulsed resource consumers. J Appl Ecol 42:1203–1213. doi:10.1111/j.1365-2664.2005.01094.x
Bratton SP (1975) The effect of the European wild boar, Sus scrofa, on gray beech forest in the Great Smoky Mountains. Ecology 56:1356–1366. doi:10.2307/1934702
Caccia FD, Chaneton EJ, Kitzberger T (2006) Trophic and non-trophic pathways mediate apparent competition through post-dispersal seed predation in a Patagonian mixed forest. Oikos 113:469–480. doi:10.1111/j.2006.0030-1299.14390.x
Campbell DJ, Atkinson IAE (2002) Depression of tree recruitment by the Pacific rat (Rattus exulans Peale) on New Zealand’s northern offshore islands. Biol Conserv 107:19–35. doi:10.1016/S0006-3207(02)00039-3
Campbell DJ, Rudge MR (1984) Vegetation changes induced over ten years by goats and pigs at Port Ross, Auckland Islands (Subantartic). N Z J Ecol 7:103–118
Challies CN (1975) Feral pigs (Sus scrofa) on Auckland Island: status and effects on vegetation and nesting sea birds. NZ J Zool 2:198–206
Choquenot D, Ruscoe WA (2000) Mouse population eruptions in New Zealand forests: the role of population density and seedfall. J Anim Ecol 69:1058–1070. doi:10.1046/j.1365-2656.2000.00462.x
Clark JS, Beckage B, Camill P, Cleveland B, Hillrislambers J, Lichter J, McLachlan J, Mohan J, Wyckoff P (1999) Interpreting recruitment limitation in forests. J Am Bot 86:1–16. doi:10.2307/2656950
Coblentz BE, Baber DW (1987) Biology and control of feral pigs on Isla Santiago, Galapagos, Ecuador. J Appl Ecol 24:403–418. doi:10.2307/2403883
Cruz FC, Donlan J, Campbell K, Carrion V (2005) Conservation action, Galapagos: feral pigs (Sus scrofa) eradication from Santiago Island. Biol Conserv 121:473–478. doi:10.1016/j.biocon.2004.05.018
Curran LM, Leighton M (2000) Vertebrate response to spatiotemporal variation in seed production of mast-fruiting Dipterocarpaceae. Ecol Monogr 70:101–128
Curran LM, Webb CO (2000) Experimental tests of the spatio-temporal scale of seed predation in mast-fruiting Dipterocarpaceae. Ecol Monogr 70:129–148
De Fina AL (1972) El clima de la región de los bosques andino-patagónicos argentinos. In: Por Dimitri MJ (ed) La región de los bosques andino-patagónicos. INTA, Buenos Aires, pp 35–53
Focardi S, Capizzi D, Monetti D (2000) Competition for acorns among wild boar (Sus scrofa) and small mammals in a Mediterranean woodland. J Zool 250:329–334. doi:10.1111/j.1469-7998.2000.tb00777.x
Gómez JM, García D, Zamora R (2003) Impact of vertebrate acorn- and seedling-predators on a Mediterranean Quercus pyrenaica forest. For Ecol Manag 180:125–134. doi:10.1016/S0378-1127(02)00608-4
Groot Bruinderink GWTA, Hazebroek E (1996) Wild boar (Sus scrofa L.) rooting and forest regeneration on podzolic soils in the Netherlands. For Ecol Manag 88:71–80. doi:10.1016/S0378-1127(96)03811-X
Hone J (1995) Spatial and temporal aspects of vertebrate pest damage with emphasis on feral pigs. J Appl Ecol 32:311–319. doi:10.2307/2405098
Howard WE (1964) Introduced browsing mammals and habitat stability in New Zealand. J Wildl Manag 28:421–429. doi:10.2307/3798193
Howe TD, Singer FJ, Ackerman BB (1981) Forage relationships of European wild boar invading northern hardwood forests. J Wildl Manag 45:748–754. doi:10.2307/3808713
Hulme PE, Hunt MK (1999) Rodent post-dispersal seed predation in deciduous woodland: predator response to absolute and relative abundance of prey. J Anim Ecol 68:417–428. doi:10.1046/j.1365-2656.1999.00294.x
Ickes K, Dewalt SJ, Appanah S (2001) Effects of native pigs (Sus scrofa) on woody understory vegetation in a Malaysian lowland rain forest. J Trop Ecol 17:191–206. doi:10.1017/S0266467401001134
Iida S (2004) Indirect negative influence of dwarf bamboo on survival of Quercus acorn by hoarding behavior of wood mice. For Ecol Manag 202:257–263. doi:10.1016/j.foreco.2004.07.022
Izquierdo F, Pelliza A, Siffredi G, Gallo L, Strizler N (2001) Araucaria araucana (Molina) K. Koch seed diet in wild and domestic herbivores. International Araucariaceae Symposium, 14–17 March, Auckland, New Zealand
Jaksic FM, Iriarte JA, Jimenez JE, Martinez DR (2002) Invaders without frontiers: cross-border invasions of exotics mammals. Biol Invasions 4:157–173. doi:10.1023/A:1020576709964
Janzen DH (1971) Seed predation by animals. Annu Rev Ecol Syst 2:465–492. doi:10.1146/annurev.es.02.110171.002341
Jedrezejewska B, Jedrzejewski W, Bunevich AN, Milkowski L, Krasinski A (1997) Factors shaping population densities and increased rates of ungulates in Bialowieza Primeval Forest (Poland and Belarus) in the 19th and 20th centuries. Acta Theriol (Warsz) 42:399–451
Jensen TS (1982) Seed production and outbreaks of non-cycling rodent populations in deciduous forests. Oecologia 54:184–192. doi:10.1007/BF00378391
Kuiters AT, Slim PA (2002) Regeneration of mixed deciduous forest in a Dutch forest-heathland, following a reduction of ungulate densities. Biol Conserv 105:65–74. doi:10.1016/S0006-3207(01)00204-X
Lever C (1985) Naturalized mammals of the world. Longman Press, London 354 pp
Lowe S, Browne M, Boudjelas S, Poorter D (2004) 100 of the world’s worst invasive alien species. A selection from the global invasive species database. Invasive Species Specialist Group published. The World Conservation Union (UICN), 10 pp
Maillard D, Fournier P (2004) Timing and synchrony of births in the wild boar (Sus scrofa LINNAEUS, 1758) in a Mediterranean habitat: the effect of food availability. Galemys 16:67–74
Massei G, Genov PV, Staines BW, Gorman ML (1997) Factors influencing home range and activity of wild boar (Sus scrofa) in Mediterranean coastal area. J Zool 242:411–423
McShea WJ (2000) The influence of acorn crops on annual variation in rodent and bird population. Ecology 81:228–238
Melis C, Szafranska PA, Jedrzejewska B, Barton K (2006) Biogeographic variation in wild boar (Sus scrofa L.) density in western Eurasia. J Biogeogr 33:803–811. doi:10.1111/j.1365-2699.2006.01434.x
Merino ML, Carpinetti BN (2003) Feral ping Sus scrofa population estimates in Bahia Samborombon Conservation Area, Buenos Aires Province, Argentina. J Neotrop Mammal 10:269–275
Moles AT, Drake DR (1999) Post-dispersal seed predation on eleven large-seeded species from the New Zealand flora: a preliminary study in secondary forest. N Z J Bot 37:679–685
Mooney HA, Mack RN, McNeel JA, Neville LE, Schei PJ, Waage JK (2005) Invasive alien species: a new synthesis. Island Press, Washington, DC 368 pp
Moran PAP (1950) Notes on continuous stochastic phenomena. Biometrika 37:17–23
Muñoz Ibañez R (1984) Análisis de la productividad de semillas de Araucaria araucana (Mol.)C. Koch, en el Area de Lonquimay—IX Región. Tesis para optar al Titulo de Ingeniero Forestal. Facultad de Ciencias Agrarias, Veterinarias y Forestales. Escuela de Ciencias Forestales, Universidad de Chile, Chile 98 pp
Nathan R, Casagrandi R (2004) A simple mechanistic model seed dispersal, predation and plant establishment: Janzen-Connell and beyond. J Ecol 92:733–746. doi:10.1111/j.0022-0477.2004.00914.x
Ostfeld RS, Jones CG, Wolff JO (1996) Ecological connections in eastern deciduous forests. Bioscience 46:323–330. doi:10.2307/1312946
Peris SJ, Pescador M, Sanguinetti J, Pastore H (2007) Colonización por el jabalí (Sus scrofa) en la región andina de la Patagonia argentina. VII Jornadas de la Sociedad Española para la Conservación y Estudio de los Mamíferos (SECEM), Huelva, España
Pulido FJ (2002) Biología reproductiva y conservación: el caso de la regeneración de bosques templados y subtropicales de robles (Quercus spp.). Rev Chil Hist Nat 75:5–15
Redford KH, Eisenberg JF (1992) Mammals of the Neotropics, vol 2 the southern cone. University of Chicago Press, 430 pp
Rosell C, Fernández-Llario P, Herrero J (2001) El Jabalí (Sus scrofa Linnaeus, 1758). Galemys 13:1–25
Ruscoe WA, Norbury G, Choquenot D (2006) Trophic interactions among native and introduced animal species. In: Allen RB, Lee WG (eds) Biological invasion in New Zealand, Chap. 16. Ecological Studies vol 1986
Sanguinetti J, Kitzberger T (2008) Patterns and mechanisms of masting in the large-seeded southern hemisphere conifer Araucaria araucana. Austral Ecol 33:78–87
Scheiner SM, Gurevitch J (2001) Design and analysis of ecological experiments, 2nd edn. Oxford University Press, 415 pp
Shepherd JD, Ditgen RS (2008) Piñones y mamiferos pequeños: predadores y dispersores en los bosques de Araucaria araucana. XXII Jornadas Argentinas de Mastozoología. Sociedad Argentina para el Estudio de los Mamíferos (SAREM). Presentación oral. Del 10 al 13 de Noviembre, Villa Giardino-Córdoba, Argentina
Shepherd JD, Ditgen RS (2005) Human use and small mammals communities of Araucaria forests in Neuquén, Argentina. Mastozool Neotrop 12:217
Silman MR, Terborgh JW, Kiltie RA (2003) Population regulation of a dominant rain forest tree by a mayor seed predator. Ecology 84:431–438. doi:10.1890/0012-9658(2003)084[0431:PROADR]2.0.CO;2
Silva CA, Saavedra B (2008) Knowing for controlling: ecological effects of invasive vertebrate in Tierra del Fuego. Rev Chil Hist Nat 81:123–136
Silvertown JW (1980) The evolutionary ecology of mast seeding in trees. Biol J Linn Soc Lond 14:235–250. doi:10.1111/j.1095-8312.1980.tb00107.x
Singer FJ, Otto DK, Tipton AR, Hable CP (1981) Home ranges, movements, and habitat use of European wild boar in Tennessee. J Wildl Manag 45:343–353. doi:10.2307/3807917
Skewes O, Rodríguez R, Jaksic F (2007) Ecología trófica del jabalí europeo (Sus scrofa) silvestre en Chile. Rev Chil Hist Nat 80:295–307
Sodeikat G, Pohlmeyer K (2001) Temporary home range modifications of wild boar family groups (Sus scrofa L.) caused by drive hunts in Lower Saxony (Germany). Eur J Wildl Res 48:161–166
Sokal RR, Rohlf FJ (1994) Biometry: The principles and practice of statistics in biological research, 3rd edn. W. H. Freeman and Company, New York 887 pp
Sweetapple PJ (2003) Possum (Trichosurus vulpecula) diet in a mast and non-mast seed year in a New Zealand Nothofagus forest. N Z J Ecol 24:123–137
Sweitzer RA, Van Vuren DH (2002) Rooting and foraging effects of wild pigs on tree regeneration and acorn survival in California’s Oak Woodland Ecosystems. USDA Forest Service Gen. Tech. Rep. PSW-GTR-184
Tisdell CA (1984) Feral pigs threaten native wildlife in Australia. Tigerpaper 2:13–18
Uzal A, Nores C (2004) Endogenous and exogenous constraints in the population changes of wild boar (Sus scrofa). Galemys 16:83–98
Veblen TT, Burns RB, Kitzberger T, Lara A, Villalba R (1995) The ecology of the conifers of southern South America. In: Enright N, Hill R (eds) Ecology of the Southern Conifers. Melbourne University Press, Parkville, Australia
Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997) Human domination of Earths’s ecosystems. Science 277:494–499. doi:10.1126/science.277.5325.494
Vtorov IP (1993) Feral pig removal:effects on soil microarthropods in Hawaiian rain forest. J Wildl Manag 57:875–880. doi:10.2307/3809092
Wada N (1993) Dwarf bamboos affect the regeneration of zoochorous trees by providing habitats to acorn-feeding rodents. Oecologia 94:403–407. doi:10.1007/BF00317116
Wilson DJ, Wright EF, Canham CD, Ruscoe WA (2003) Effects of possums and rats on seedling establishment at two forest sites in New Zealand. N Z J Ecol 27:147–155
Wilson DJ, Wright EF, Canham CD, Ruscoe WA (2007) Neighbourhood analyses of tree seed predation by introduced rodents in a New Zealand temperate rainforest. Ecography 30:105–119
Wolff JO (1996) Population fluctuations of mast-eating rodents are correlated with production. J Mammal 77:850–856. doi:10.2307/1382690
Wood GW, Roark DN (1980) Food habits of feral hogs in coastal South Carolina. J Wildl Manag 44:506–511. doi:10.2307/3807990
Acknowledgments
The authors thank Santiago Quiroga and the summer students and volunteers from Global Vision International (GVI) for their help during the field work. We thank Rebecca Ditgen and John Shepherd for sharing the information provided by the camera trapping field work. We thank John Shepherd and Richard Sage for improvement of the manuscript. This work is part of the Ph.D. thesis of JS supported by the German Government (GMZ Minister) and the International Plant Genetic Resources Institute (IPGRI), by the Russell Train Scholarship (WWF), by the Agencia Española de Coop. Internacional (AECI) with the project CGL2004-0176-Feder (Salvador J. Peris coord.) and by the Argentina National Park Administration (Lanín National Park).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sanguinetti, J., Kitzberger, T. Factors controlling seed predation by rodents and non-native Sus scrofa in Araucaria araucana forests: potential effects on seedling establishment. Biol Invasions 12, 689–706 (2010). https://doi.org/10.1007/s10530-009-9474-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-009-9474-8