
Abstract
Research on the impacts of house mice Mus musculus introduced to islands is patchy across most of the species’ global range, except on islands of the Southern Ocean. Here we review mouse impacts on Southern Ocean islands’ plants, invertebrates, land birds and seabirds, and describe the kinds of effects that can be expected elsewhere. A key finding is that where mice occur as part of a complex of invasive mammals, especially other rodents, their densities appear to be suppressed and rat-like impacts have not been reported. Where mice are the only introduced mammal, a greater range of native biota is impacted and the impacts are most severe, and include the only examples of predation on seabird eggs and chicks. Thus mice can have devastating, irreversible and ecosystem-changing effects on islands, impacts typically associated with introduced rats Rattus spp. Island restoration projects should routinely include mouse eradication or manage mouse impacts.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The impacts of invasive alien mammals are most profound on insular fauna, flora and ecosystems (Diamond 1989; Williamson 1996). In particular, the predatory impacts of rats Rattus rattus, R. norvegicus and R. exulans, cats Felis catus and pigs Sus scrofa on insular ecology and charismatic fauna such as seabirds are such that their eradication from islands has progressed apace (Veitch and Clout 2002; Courchamp et al. 2003; Parkes and Murphy 2003; Towns and Broome 2003; Nogales et al. 2004; Howald et al. 2007; Jones et al. 2008). House mice Mus musculus are one of the most widespread invasive mammals on earth, and amongst vertebrates the breadth of their global distribution is second only to that of humans (Bronson 1979; Brooke and Hilton 2002). Despite this, there has been little island conservation action devoted to mice, relative to other introduced mammals (Wanless et al. 2007; Howald et al. 2007). Why has one of the most commonly introduced mammals on islands been so widely ignored by island conservation programmes? Two reasons are suggested. First is the relative paucity of described impacts on charismatic fauna (cf. Fritts and Rodda 1998), or lack of devastating herbivory, e.g. by goats Capra hircus (Campbell and Donlan 2005), especially relative to the impacts of other introduced mammals. Second, a higher percentage of mouse eradications has failed compared to Rattus spp. eradications (38 vs. 5–10%, Howald et al. 2007; MacKay et al. 2008), although the reasons for the higher rate of failed mouse campaigns remain unclear (MacKay et al. 2008). These factors have probably contributed to the slow development of technology for successful mouse eradications, and the patchy coverage of eradication attempts, relative to rat eradications.
Purpose of this review
The impacts of introduced mice on seabirds have only recently been assessed critically, in contrast to studies of their impacts on other biota from Southern Ocean islands; here we review all their described impacts. The impacts of mice preying on island-endemic invertebrates and causing extinctions, e.g. on Antipodes Island (Marris 2000) and Marion Island (Chown et al. 2002) or precipitating potentially irreversible changes to ecosystem functioning, e.g. Marion Island (Smith et al. 2002), have, to the best of our knowledge, failed to generate wide scientific, popular or conservation interest. By contrast, descriptions of conclusive proof that mice were preying on Gough Island’s seabirds (Angel et al. 2005; Wanless et al. 2005) led to substantial media interest (e.g. Marris 2005; Dangerfield 2006; Pearce 2006), an unsolicited offer of support for eradication from the New Zealand Department of Conservation (G. M. Hilton, personal communication) and the establishment of an advisory group to raise funds for and oversee a process aimed at eradicating mice (Angel and Cooper 2006). The subsequent publication describing those impacts in a peer-reviewed, international journal (Wanless et al. 2007) led to renewed media interest (e.g. Millius 2007). The relative importance of conserving endemic invertebrates or plants is, in theory, the same as conserving charismatic fauna such as albatrosses. However, the evidence suggests that in reality, less visible fauna and flora tend not to generate sympathetic responses or to drive island conservation actions. It is our belief that negative impacts of mice on any insular systems, either direct or indirect (such as through changing nutrient cycles, synergy with other invasive species or changes to other major ecosystem processes) are sufficient grounds to merit remedial action as a high conservation priority. Demonstrating negative impacts on charismatic species such as seabirds should not be a pre-requisite for planning or securing funding for island conservation and restoration projects.
Here we summarise the impacts of mice on the plants, invertebrates and native birds of the islands of the Southern Ocean. The regional focus is chosen because various national Antarctic research programmes have facilitated the publishing of a substantive body of relevant research within the biogeographic setting. Special focus is given to those islands where mice are currently the only invasive mammal, some of which have been particularly well studied, and these studies describe the kinds of impacts that could be expected elsewhere, in more temperate and tropical systems where such studies are lacking. The aim of this review is to describe the distribution of introduced house mice on islands in the Southern Ocean and to draw attention to the importance of mouse impacts on island ecosystems, impacts that may be overlooked or (more probably) suppressed on islands where mice occur together with other introduced mammals. The long-term consequences of the impacts, both direct and indirect, of invasive house mice are as important to consider as the impacts of more widely studied invasive mammals on islands. Finally, we point to the importance of including the management of mouse impacts in island restoration programmes.
Regional distribution of mice
The house mouse is one of the most widely introduced mammals on oceanic islands, occurring on >200 islands where Red-listed birds are also found (Brooke and Hilton 2002; Brooke et al. 2007; G. M. Hilton, personal communication). However, it is seldom the only introduced species (Berry et al. 1978; Bronson 1979; Brooke et al. 2007). Thus, across most of their non-native range, the impacts of mice on island biota appear to be difficult to separate from those of other introduced mammal species.
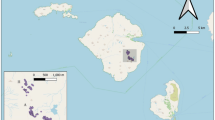
For the purposes of this review, we have defined the Southern Ocean to include islands slightly to the north of the Subtropical Convergence (such as Tristan da Cunha, Amsterdam and St Paul) and islands well south of the Antarctic Convergence (Heard and McDonald) (Fig. 1). Only oceanic islands are considered. Antipodes and Gough islands have mice as the only introduced mammal (King 1990; Jones et al. 2003a; Angel and Cooper 2006). On Marion Island, mice have been present since the 1800s (Jansen van Vuuren and Chown 2007), whereas the domestic cat was introduced in the 1950s and eradicated in the early 1990s (Bester et al. 2000). On the French islands of St Paul and Australie (Kerguelen group), mice remain after successful eradications of ship rats Rattus rattus and rabbits Oryctolagus cuniculus, in both cases the only other introduced mammals (Micol and Jouventin 2002, T. Micol, in litt.). All other Southern Ocean islands/island groups either lack mice or currently have other mammals in addition to mice (Table 1).

Islands of the Southern Ocean. Islands with house mice as the only introduced mammal currently present are indicated by arrows
Impacts on flora
On Antipodes Island, sedge species, in particular the native Carex appressa, attract large concentrations of mice that feed extensively on the inflorescences and seeds (Godley 1989). A more recent diet study found a large percentage of mouse diet on the Antipodes to consist of native sedges and plants in general (McIntosh 2001). On Gough Island at least 15 species of endemic or native as well as seven introduced plant species are consumed by mice (Jones et al. 2003a). The flower heads and fruiting bodies of the native tussock grasses Spartina arundinacea and Parodiochloa flabellata are chewed extensively by mice as are the seeds of the herbaceous Acaena sarmentosa, and endemic sedges Carex spp. and Scirpus spp. (Wace 1986; Jones et al. 2003a). On Macquarie Island mice consume a range of native plant species, some of which are available throughout the year (Copson 1986). Comparative studies between mouse-free Prince Edward Island and Marion Island are to date the most comprehensive with regards to the impacts that mice have on Southern Ocean island flora. The mice on Marion Island feed on the seeds and young shoots of at least five native and one introduced plant species. In the late 1960s the native sedge Uncinia compacta was abundant in mire vegetation (Huntley 1971) but has now almost been extirpated from this habitat on Marion due to seed predation by mice (Smith and Steenkamp 1990). Mice remove up to 75% of all the seed heads of this native plant, significantly reducing the population compared to the neighbouring mouse-free island of Prince Edward (Chown and Smith 1993). Given the long time that mice have been present on Marion Island and the relatively rapid decrease observed in this sedge species, it is possible that diet switches by mice caused similar impacts (or even extinctions) on other native/endemic species, before plant distributions and densities were mapped. Mice are also thought to be having a similar impact on the herbaceous Acaena magellanica (Avenant 1999) and damage the peat-forming Azorella selago (a disturbance-sensitive cushion plant) by burrowing into it (Chown and Cooper 1995). On Guillou Island (Kerguelen group), the seeds and flowers of the indigenous Acaena magellanica were among the main plant items consumed during summer (Le Roux et al. 2002). These studies all suggest a preference for native plants over introduced species, possibly related to the lack of defensive traits such as secondary compounds, which may be lost among native island plants (Bowen and van Vuren 1997). There is, however, no evidence of plant species having gone extinct on any of these islands due to mice (Wace and Dickson 1965; Godley 1989; Copson and Whinam 1998). Excluding Marion Island, there are no studies on the possible population-level impacts that mouse predation has on the plants on any of these islands, possibly reflecting a paucity of data rather than a lack of effect. The importance of plant material compared to invertebrates in mouse diet on Marion Island has increased over time, from occurring on average in 36% of stomachs in the late 1970s to 59% in the 1990s, particularly during mid to late-summer (Avenant 1999; Smith et al. 2002). The seasonal pattern may reflect prey-switching from invertebrates to seeds as invertebrate densities decrease (Avenant and Smith 2004), but could also be due to optimal foraging in the presence of abundant seed loads. Similar patterns of seasonal changes in the importance of dietary groups have been found on Guillou and Gough islands (Le Roux et al. 2002; Jones et al. 2003b).
Impacts on invertebrate fauna
Although mice are omnivorous they can also be highly selective and may prefer invertebrate prey. This has been documented for Southern Ocean islands such as Marion, Macquarie, Antipodes and Guillou (Gleeson 1981; Copson 1986; McIntosh 2001; Le Roux et al. 2002). Comparative studies of mouse-free and mouse-invaded island pairs (Bollons-Antipodes and Prince Edward-Marion) strongly suggest that mice have a major impact on invertebrates at three levels: species composition, relative abundances and size distributions. Predation by mice is considered responsible for local extinctions of several invertebrate species on Antipodes Island (Marris 2000) and also for the absence of the flightless moth Pringleophaga kerguelensis on Marion (Vári 1971). Differences in invertebrate abundances between Bollons Island and Antipodes Island are dramatic and have been attributed to mouse predation on Antipodes (Marris 2000). Mice on Marion are strongly size-selective feeders, preferring the larger individuals of moth larvae and weevils (Crafford and Scholtz 1987). This has resulted in quantitative differences in invertebrate body size and biomass between Marion and Prince Edward islands (Crafford and Scholtz 1987; Chown and Smith 1993). Further quantitative and robust analyses of population trends of moth larvae and weevil larvae on Marion Island showed that densities decreased significantly, by approximately an order of magnitude, between 1976/77 and 1996/97 (Chown et al. 2002). Flightless island species are particularly vulnerable to introduced predators (Carlquist 1965; Hobbs 1989; Quammen 1996), and the flightless Lepidoptera on Antipodes and Gough islands are heavily impacted by mouse predation (Patrick 1994; Jones et al. 2002); indigenous moths are major prey items wherever mouse diets have been studied in the Subantarctic (Gleeson and van Rensburg 1982; Copson 1986; Le Roux et al. 2002). Inter-decadal comparisons from Marion Island showed that the relative importance of the main invertebrate species in mouse diet has changed since the late 1970s (Chown and Smith 1993). By the 1990s, endemic moth larvae had been replaced by native weevil adults, in particular Ectemorrhinus similis and Bothrometopus randi, as the main prey. This prey shift suggests a marked impact on their previously preferred prey (Chown and Smith 1993).
A strong preference by mice for large, slow-growing invertebrate species has serious consequences, given the life-history traits of some species (Peters 1983; Brown et al. 1993). For example, adults of the flightless moth Pringleophaga marioni are short-lived (10–14 days) and have very low dispersal abilities (Rowe-Rowe et al. 1989) and larvae take 2–3 years to mature (Crafford 1990). The decreased abundance of this species in mouse diets on Marion suggests over-harvesting, and unless remedial action is taken, local extinction is possible. P. marioni plays a critical role in Marion Island’s nutrient cycle; where invertebrate species are also keystone species, predation can have deleterious effects for the whole ecosystem (Smith 1978; Smith and Steenkamp 1992; Smith and Steenkamp 1993). House mice could have an indirect negative effect on invertebrates, by reducing the abundance of important resources, such as a particular native plant that is important for a stage of an invertebrate’s life-cycle.
Impacts on terrestrial birds
Relatively few reliable studies have shown any direct impacts of mice on native terrestrial birds. On Gough Island, direct nest predation and competition are believed to explain the reduced abundance of the endemic Gough Bunting Rowettia goughensis in the lowlands, but there are few quantitative data to support this contention. During the 2000/01 breeding season, four of 15 monitored Gough Bunting nests (27%) were apparently depredated by mice, one during incubation, and three during the chick stage (Cuthbert and Hilton 2004). A study using artificial nests and eggs found evidence of mouse predation, although such studies are difficult to interpret (Faaborg 2004; Thompson and Burhans 2004).
Mouse predation on invertebrates may have an indirect influence on several island bird species. Lesser Sheathbill Chionis alba numbers on Prince Edward Island have remained relatively constant since the 1970s but decreased on Marion Island by 20% over the same period (Burger 1978; Huyser et al. 2000). Observations on Marion over the last 20 years indicate that Lesser Sheathbills now forage mostly on the coast during winter and not inland as they did previously and still do on Prince Edward (Huyser et al. 2000). The change in foraging behaviour is thought to be due to mouse predation reducing invertebrate densities (Crafford and Scholtz 1987; Rowe-Rowe et al. 1989). The sheathbill population relies heavily on invertebrates during the winter when other, preferred food resources in penguin or seal colonies are not available (Burger 1982). Now sheathbills on Marion commence breeding at a lower body mass and lay smaller clutches (Huyser et al. 2000). Kelp Gulls Larus dominicanus also forage more on invertebrates during winter (Burger 1978). Mouse predation on invertebrates may thus impact this species in a similar manner as it does the Lesser Sheathbill.
A comparative study of Antipodes Island Snipe Coenocorypha aucklandica meinertzhagena and Auckland Island Snipe C. aucklandica on Adams Island (free of introduced mammals) found that the former was much less abundant than expected, and they are probably affected by mice consuming invertebrates in a similar manner to the Lesser Sheathbills on Marion Island (Miskelly et al. 2006). Mice on Antipodes Island may also be in direct competition with the Antipodes Parakeet Cyanoramphus unicolor as scavengers of Subantarctic Skua Catharacta antarctica and Northern Giant-Petrel Macronectes halli kills (Imber et al. 2005).
Impacts on seabirds
In this section we draw on all available data, not just from Southern Ocean islands. The direct impact of house mice on seabird populations was until recently considered negligible (Moors and Atkinson 1984; Atkinson 2001, Witmer et al. 2007). Reported impacts were restricted to predation of eggs and young chicks of very small seabirds, such as storm-petrels (of which newly hatched chicks may weigh only 10 g). Only four credible examples are known to us, excluding Gough Island. Mice were suspected of depredating eggs of the Grey-backed Storm-Petrel Garrodia nereis on Antipodes Island (Burger and Gochfeld 1994), eggs and chicks of the White-faced Storm-Petrel Pelagodroma marina on Selvagem Grande Island, Madeiran Archipelago (Campos and Granadeiro 1999), eggs and small chicks of the Ashy Storm-Petrel Oceanodroma homochroa on the Farallon Islands (Ainley et al. 1990) and Blue Petrel Halobaena caerulea chicks (ca 30 g) on Marion Island (Fugler et al. 1987), where a chick was found with deep wounds on its back and neck. We have disregarded an unsupported statement of depredation of nests of the Polynesian Storm-Petrel Nesofregetta fuliginosa (BirdLife International 2004) and other unsubstantiated reports in the grey literature. Bird remains in mouse stomach samples are a possible indication of mouse predation, e.g. on Macquarie and Antipodes islands, although these could derive from scavenging (Copson 1986; McIntosh 2001). Also on Antipodes Island, the abundance of species such as Black-bellied Storm-Petrels Fregatta tropica on Bollons and other mouse-free offshore islets, but their extremely low density on Antipodes, has been speculatively attributed to predation by mice (Imber et al. 2005).
All of the above reported incidents of predation are incidental, inferred or based on post hoc observations of gnawed egg shells, wounded chicks and mice eating dead chicks. Without direct observations of mice depredating nests or killing live chicks, the possibility that mice were only scavenging abandoned eggs and moribund or dead chicks cannot be excluded. Research conducted on Gough Island during 2003/4 was the first to show conclusive evidence that large and previously healthy chicks of the burrowing Atlantic Petrel Pterodroma incerta (weighing 70–700 g) and surface-nesting Tristan Albatross Diomedea dabbenena chicks (weighing 0.3–8 kg) were being preyed upon and killed by mice (Wanless et al. 2007; see also Wanless 2007). Wanless et al. (2007) suggest that this behaviour may not be restricted to Gough Island and may have been overlooked on other islands, for example due to the practical difficulties in studying activity in burrowing petrel nests, and the more obvious impacts of rats and cats where present with mice. Recent events on Marion Island support this hypothesis. House mice on Marion became the sole introduced mammal following the eradication of cats Felis catus in the 1990s (Bester et al. 2000). Since 2004, several Wandering Albatross Diomedea exulans chicks have succumbed to wounds consistent with mouse attacks (P. G. Ryan, personal communication; RMW personal observation). These are the first records of wounded chicks in over 20 years of intensive study (P. G. Ryan, personal communication). A key observation is that to date, the only records of (probable or confirmed) mouse predation on seabird eggs and chicks are from islands on which mice are the only introduced mammal. On islands where the mice are part of a complex of invasive mammals, the effects of dominance, competition and predation by larger species may render them less of a threat to native vertebrates (Courchamp et al. 1999; Wanless et al. 2007, Harris and Macdonald 2007; Quillfeldt et al. 2008).
The dynamics of seabird-nutrient cycle interactions have been well studied in the Subantarctic (Smith 1978, 1979; Smith and Steenkamp 1990, 1992, 1993) and elsewhere (Croll et al. 2005; Fukami et al. 2006). The transport of nutrients from marine to terrestrial systems is a critical determinant of terrestrial productivity on Subantarctic and other seabird-dominated islands (Smith 1978; Smith 1979; Smith and Steenkamp 1993; Erskine et al. 1998; Croll et al. 2005; Fukami et al. 2006; Maron et al. 2006; Mulder et al. 2008). Significant reductions in seabird densities due to predation will ultimately reduce nitrogenous fertilisation, leading to lower primary productivity (e.g. Croll et al. 2005; Fukami et al. 2006). This could cause mice to rely more heavily on predation of seabird nests, setting up a positive feedback and precipitating rapid decreases in seabird numbers.
Effects of climate change on mouse impacts
Climatic change is expected to be marked in the Southern Ocean, and empirical evidence of change already has been reported from Macquarie, Kerguelen, Marion, Heard and South Georgia (Allison and Keague 1986; Bergstrom and Chown 1999; van Aarde and Jackson 2006). There is now abundant evidence that temperatures and hours of sunshine/day are increasing and rainfall regimes are changing at several islands (Allison and Keague 1986; Smith and Steenkamp 1990; Smith 2002). The actual causes for these changes are unknown, but they may reflect changing atmospheric circulation patterns, associated with changes in the oceanic circulation patterns (Smith and Steenkamp 1990).
The impacts of climate change may affect mouse populations directly in two ways. First, if mice are temperature-stressed and experience thermally induced die-offs during winter (e.g. at Marion Island, Crafford and Scholtz 1987; Crafford 1990), then ameliorating conditions could facilitate higher survivorship in winter, leading to higher densities at the start of the breeding season and potentially higher average or peak densities (Rowe et al. 1964; King 1982; Singleton et al. 2001, 2005; Ruscoe et al. 2005; Ferreira et al. 2006). Second, climate change has the potential to increase mouse densities by enhancing terrestrial productivity, through lengthened growing seasons for plants or longer breeding seasons for mice and invertebrates. A relaxation of the constraints of temperature and resource limitation could facilitate greater variability in annual density cycles, possibly leading to irruptions, such as has been observed in Australia and New Zealand (King 1982; Singleton et al. 2001, 2005). This would only add complexity and difficulty to mouse population management. However, these effects may be self-regulating or offset because of deleterious effects of higher mouse densities on ecosystem functioning. On Marion Island, an increasing mouse population is likely to place enhanced predation pressure on soil invertebrates. This will decrease rates of nutrient cycling (nutrient availability and mineralization) with a negative effect on primary productivity (Crafford 1990; Chown and Smith 1993; Smith et al. 2002). Similarly, on Gough Island it is possible that increased mouse densities in response to climate-linked variables could be offset by reduced terrestrial productivity due to lower seabird densities and concomitant reduction in nitrogenous fertilisation.
Global relevance
The impacts of introduced mice on island ecosystems and species are poorly described across most of their range (see reviews by Harris (2008) and Traveset et al. (2008) on impacts of introduced rodents on islands). However, the islands of the Southern Ocean represent an exception to this pattern. Mice are the only introduced mammal on at least five of these islands (Antipodes, Australie, Gough, Marion and St. Paul), and it is on three of these islands that their impacts appear to be the most significant of all mouse-invaded islands, although to some extent this may reflect relative research effort and/or time since eradication of other invasive species. Where mice co-occur on islands with other introduced mammals, their densities are suppressed (e.g. Miller and Miller 1995; Choquenot and Ruscoe 2000; Russell and Clout 2004; Witmer et al. 2007; Caut et al. 2008; Simberloff 2008); for example, the eradication of Rattus rattus from Buck Island (Caribbean) was followed by an irruption of house mice (Witmer et al. 2007). It is probable that their (visible?) impacts are diminished as a consequence. However, eradications of other introduced mammals from islands are leaving increasing numbers of islands with mice as the only introduced mammal, including the Southern Ocean islands of Marion, St. Paul and Australie (Kerguelen group) (Bester et al. 2000; Micol and Jouventin 2002; T. Micol, in litt.). Far more islands around the world have had rats Rattus spp. eradicated than mice (Howald et al. 2007), and the total island area cleared of rats globally dwarfs that cleared of mice (Fig. 2). Ironically, the largest island cleared of house mice to date is Enderby (710 ha, Auckland Island group), but the operation was targeted at rabbits and the successful eradication of mice was incidental (Torr 2002). To date, the simultaneous eradication of mice and rats has been achieved on nine islands, three of which involved R. rattus and six involved R. norvegicus. At least five rat eradications have failed to eradicate mice simultaneously (including the Southern Ocean islands of St. Paul and Australie) (Micol and Jouventin 2002; Howald et al. 2007; MacKay et al. 2008; T. Micol, in litt.).

Island area cleared of rats (Rattus spp.) and house mice (Mus musculus). All known efforts prior to 1986 are summed. Modified from Wanless et al. (2007)
The incidence and impacts of rat predation on island biota vary widely and depend on factors such as the physical environment of the island, seasonal availability of food, the size, behaviour and population density of the rats and the presence of other rat species and/or other predators or competitors (Woodward 1972; Atkinson 1978; Taylor 1979; Moors and Atkinson 1984). Although the effects of these factors have been studied principally for rats, there is evidence that the impacts of mice vary according to similar factors (Chown and Smith 1993; Smith et al. 2002; Wanless et al. 2007). The evidence of significant impacts by mice on birds of the Southern Ocean islands is limited. What little evidence we found was mostly indirect, due to competition for food. Wanless et al. (2007) have, however, shown significant effects of mouse predation on the chicks of seabirds as large as albatrosses. Outside the Southern Ocean, especially on islands where small passerines or ground-nesting birds breed that do not have strong nest-defence or predator-avoidance instincts, the impacts of mouse predation could be/become significant. Small rodents have been shown to be significantly more motivated to attack chicks and depredate eggs when denied food, in contrast to when provided with alternatives ad libitum (Bradley and Marzluff 2003). This finding is in agreement with the general pattern of mouse predation of seabirds on Gough Island, which is mostly restricted to the lean winter months (Wanless et al. 2007). We predict that where house mice are or become the only introduced mammal, including on temperate or tropical islands, nest predation is likely to occur during times of severe food-stress, such as winter seasons or dry monsoons.
Mouse impacts have probably been overlooked to some extent, but this review shows that rat-like behaviour and impacts can be expected, especially on islands where mice are the only introduced mammal. Predation by introduced mice on seeds, invertebrates and the eggs and chicks of birds is causing significant changes to species distributions, densities and persistence on islands in the Southern Ocean. Some of these impacts are likely to result in extinction of impacted taxa and irreversible effects on ecosystem functioning and similar effects are likely to occur on islands outside the Southern Ocean. There is thus a strong case for eradicating introduced mice from islands. It may be important to eradicate mice and other species simultaneously, or to give careful consideration to the best order of a staged, multi-species eradication programme, for three reasons. First, if toxic baits are applied to eradicate mice, primary or secondary poisoning of ‘non-target’ species could achieve eradication or substantial knockdown of numbers (e.g. of cats), making subsequent operations for those species cheaper and more efficient. Second, higher densities of mice following release from competition or predation could conceivably accelerate extinction of prey sources (e.g. endemic invertebrates) (Courchamp et al. 1999; Harris and Macdonald 2007; Caut et al. 2007, 2008); thus eradication of one invasive species could lead indirectly to the extinction of a native or endemic species (Zavaleta 2002). Third, a rebounding mouse population or rejuvenating island ecology (e.g. higher seed loads after release from herbivory) might compromise the susceptibility of mice to eradication for an unknown period of time, while exacerbating their negative biodiversity impacts.
Island ecosystems are beguilingly simple, but should nevertheless be treated as a whole (Poiani et al. 2000; Zavaleta et al. 2001; Caut et al. 2008). Where several invasive mammals coexist, eradicating one could have profound and perhaps unpredictable effects on the interactions between native species and other invasive species. We recommend that the management of mice be included in island restoration plans while taking due cognisance of the difficulties of eradicating multiple species from an island, such as is being planned for Subantarctic Macquarie Island (Anon. 2007). Reliable methods that result in the simultaneous eradication of mice and rats should be a high priority for island conservation research. In addition, changes in the diet and behaviour of mice on Marion, St. Paul and Australie should be studied, as these provide excellent and immediate ‘natural’ experiments on the effects of competitive release or meso-predator release and could give insights into causal mechanisms for the evolution, or the lack of evolution, of predatory behaviour in house mice.
References
Ainley DG, Henderson RP, Strong CS (1990) Leach’s and Ashy Storm-Petrel. In: Ainley DG, Boekelheide RJ (eds) Seabirds of the Farallon Islands. Stanford University Press, Palo Alto, pp 128–162
Allison F, Keague PL (1986) Recent changes in the glaciers of Heard Island. Polar Rec 23:255–271
Angel A, Cooper J (2006) A review of the impacts of introduced rodents on the islands of Tristan da Cunha and Gough. Research report no. 17. Royal Society for the Protection of Birds, Sandy
Angel A, Wanless RM, Hilton GM, Ryan PG (2005) Niche expansion, competitive release and the evolution of predation in the house mouse: lessons from Gough Island, South Atlantic. Paper presented at the Society for Conservation Biology Conference, Brasilia, Brazil, 15–19 July 2005
Anon. (2006) Draft Prince Edward Islands management plan. Department of Environmental Affairs and Tourism, Pretoria
Anon. (2007) Plan for the eradication of rabbits and rodents on Subantarctic Macquarie Island. Parks and Wildlife Service, Department of Tourism, Arts and the Environment, Tasmania & Biodiversity Conservation Branch, Department of Primary Industries and Water, Tasmania
Atkinson IAE (1978) Evidence for effects of rodents on the vertebrate wildlife of New Zealand Islands. In: Dingwall PR, Atkinson IAE, Hay C (eds) The ecology and control of rodents in New Zealand nature reserves. Information series No. 4. Department of Lands and Survey, Wellington, pp 7–31
Atkinson IAE (2001) Introduced mammals and models for restoration. Biol Conserv 99:81–96
Avenant NL (1999) The ecology and ecophysiology of Marion Island mice, Mus musculus L. PhD Thesis, Department of Zoology and Entomology. University of the Orange Free State, Bloemfontein, pp 160
Avenant NL, Smith VR (2004) Seasonal changes in age class structure and reproductive status of mice on Marion Island (Subantarctic). Polar Biol 27:99–111
Bergstrom DM, Chown SL (1999) Life at the front: history, ecology and change on Southern Ocean islands. Trends Ecol Evol 14:472–477
Berry RJ, Peters J, van Aarde RJ (1978) Subantarctic house mice colonization, survival and selection. J Zool (Lond) 184:127–141
Bester MN, Bloomer JP, Bartlett PA, Muller DD, van Rooyen M, Buechner H (2000) Final eradication of feral cats from Subantarctic Marion Island, southern Indian Ocean. S Afr J Wildl Res 30:53–57
BirdLife International (2004) Threatened birds of the world 2004 (CD-ROM). BirdLife International, Cambridge
Bowen L, van Vuren D (1997) Insular endemic plants lack defences against herbivores. Conserv Biol 11:1249–1254
Bradley J, Marzluff J (2003) Rodents as nest predators: influences on predatory behavior and consequences to nesting birds. Auk 120:1180–1187
Bronson FH (1979) The reproductive ecology of the house mouse. Q Rev Biol 54:265–299
Brooke MdL, Hilton GM (2002) Prioritising the world’s islands for vertebrate eradication programmes. Aliens 16:12–13
Brooke MdL, Hilton GM, Martins TLF (2007) Prioritizing the world’s islands for vertebrate-eradication programmes. Anim Conserv 10:380–390
Brown JH, Marquet PA, Taper ML (1993) Evolution of body size: consequences of an energetic definition of fitness. Am Nat 142:573–584
Burger AE (1978) Terrestrial invertebrates: a food resource for birds at Marion Island. S Afr J Antarct Res 8:87–100
Burger AE (1982) Foraging behaviour of lesser sheathbills Chionis minor exploiting invertebrates on a Subantarctic island. Oecologia 52:236–245
Burger J, Gochfeld M (1994) Predation and effects of humans on island-nesting seabirds. In: Nettleship DN, Burger J, Gochfeld M (eds) Seabirds on islands: threats, case studies and action plans. BirdLife International, Cambridge, pp 36–67
Campbell K, Donlan CJ (2005) Feral goat eradications on islands. Conserv Biol 19:1362–1374
Campos JL, Granadeiro JP (1999) Breeding biology of the White-faced Storm-Petrel on Salvagem Grande Island, North-East Atlantic. Waterbirds 22:199–206
Carlquist S (1965) Island life: a natural history of the islands of the world. American Museum of Natural History, New York
Caut S, Casanovas JG, Virgos E, Lozano J, Witmer GW, Courchamp F (2007) Rats dying for mice: modelling the competitor release effect. Austral Ecol 32:858–868
Caut S, Angulo A, Courchamp F (2008) Avoiding surprise effects on Surprise Island: alien species control in a multi-trophic level perspective. Biol Inv. doi:10.1007/s10530-008--9397-9
Chapuis J-L, Bouss P, Barnaud G (1994) Alien mammals, impact and management in the French Subantarctic islands. Biol Conserv 67:97–192
Choquenot D, Ruscoe WA (2000) Mouse population eruptions in New Zealand forests: the role of population density and seedfall. J Anim Ecol 69:1058–1070
Chown SL, Cooper J (1995) The impact of feral house mice at Subantarctic Marion island and the desirability of eradication. Report on a workshop held at the University of Pretoria, 16–17 February 1995. Directorate: Antarctica & Islands, Department of Environmental Affairs and Tourism, Pretoria
Chown SL, Smith VR (1993) Climate change and the short-term impact of feral house mice at the sub-Antarctic Prince Edward Islands. Oecologia 96:508–516
Chown SL, McGeoch MA, Marshall DJ (2002) Diversity and conservation of invertebrates on the Subantarctic Prince Edward Islands. Afr Entomol 10:67–82
Clark M, Dingwall P (1985) Conservation of islands in the Southern Ocean: a review of the protected areas of Insulantarctica. IUCN, Gland
Copson GR (1986) The diet of the introduced rodents Mus musculus L. and Rattus rattus L. on Subantarctic Macquarie Island. Aust J Wildl Res 13:441–445
Copson GR, Whinam J (1998) Response of vegetation on Subantarctic Macquarie Island to reduced rabbit grazing. Aust J Bot 46:15–24
Courchamp F, Langlais M, Sugihara G (1999) Cats protecting birds: modelling the mesopredator release effect. J Anim Ecol 68:282–292
Courchamp F, Chapuis J-L, Pascal M (2003) Mammal invaders on islands: impact, control and control impact. Biol Rev 78:347–383
Crafford JE (1990) The role of feral house mice in ecosystem functioning on Marion Island. In: Kerry KR, Hempel G (eds) Antarctic ecosystems. Ecological change and conservation. Springer-Verlag, Berlin, pp 359–364
Crafford JE, Scholtz CH (1987) Quantitative differences between the insect faunas of Subantarctic Marion and Prince Edward islands: a result of human intervention? Biol Conserv 40:255–262
Croll DA, Maron JL, Estes JA, Danner EM, Byrd GV (2005) Introduced predators transform subarctic islands from grassland to tundra. Science 307:1959–1961
Cuthbert R, Hilton G (2004) Introduced house mice Mus musculus: a significant predator of endangered and endemic birds on Gough Island, South Atlantic Ocean? Biol Conserv 117:483–489
Dangerfield W (2006) Killer mice. Natl Geogr 210:26
Diamond J (1989) The present, past and future of human-caused extinctions. Philos Trans R Soc London B 325:469–476
Erskine PD, Bergstrom DM, Schmidt S, Stewart GR, Tweedie CE, Shaw JD (1998) Subantarctic Macquarie Island—a model ecosystem for studying animal-derived nitrogen sources using 15N natural abundance. Oecologia 117:187–193
Faaborg J (2004) Truly artificial nest studies. Conserv Biol 18:369–370
Ferreira S, van Aarde R, Wassenaar T (2006) Demographic responses of house mice to density and temperature on Subantarctic Marion Island. Polar Biol 30:83–94
Frenot Y, Chown SL, Whinam J, Selkirk PM, Convey P, Skotnicki M, Bergstrom DM (2004) Biological invasions in the Antarctic: extent, impacts and implications. Biol Rev 79:1–28
Fritts TH, Rodda GH (1998) The role of introduced species in the degradation of island ecosystems: a case history of Guam. Annu Rev Ecol Syst 29:113–140
Fugler SR, Hunter S, Newton IP, Steele WK (1987) Breeding biology of Blue Petrels Halobaena caerulea at the Prince Edwards Islands. Emu 87:103–110
Fukami T, Wardle DA, Bellingham PJ, Mulder CPH, Towns DR, Yeates GW, Bonner KI, Durrett MS, Grant-Hoffman MN, Williamson WM (2006) Above- and below-ground impacts of introduced predators in seabird-dominated island ecosystems. Ecol Lett 9:1299–1307
Gleeson JP (1981) The ecology of the House mouse, Mus musculus Linnaeus, on Marion Island. MSc Thesis, Faculty of Science. University of Pretoria, Pretoria, South Africa, p 112
Gleeson JP, van Rensburg PJJ (1982) Feeding ecology of the house mouse Mus musculus on Marion Island. S Afr J Antarct Res 12:34–39
Godley EJ (1989) The flora of Antipodes Island. N Z J Bot 27:531–563
Harris DB (2008) Review of negative effects of introduced rodents on small mammals on islands. Biol Inv. doi:10.1007/s10530-008-9393-0
Harris DB, Macdonald DW (2007) Interference competition between introduced black rats and endemic Galápagos rice rats. Ecology 88:2330–2344
Hobbs RJ (1989) The nature and effects of disturbance relative to invasions. In: Drake JA, Mooney HA, di Castri F, Groves RH, Rejmanek FJ, Williamson M (eds) Biological invasions: a global perspective. Wiley, Chichester, pp 389–405
Holdgate MW (1969) Biological work on Tristan da Cunha Island, Nightingale Island, Inaccessible Island and Gough Island, 1968. Polar Rec 14:661–662
Howald G, Donlan CJ, Galvan JP, Russell JC, Parkes J, Samaniego A, Wang Y, Veitch D, Genovesi P, Pascal M, Saunders A, Tershy B (2007) Invasive rodent eradication on islands. Conserv Biol 21:1258–1268
Huntley BJ (1971) Vegetation. In: van Zinderen Bakker Sr EM, Winterbottom JM, Dyer RA (eds) Marion and Prince Edward Islands. Report on the South African biological and geological expedition/1965–1966. A.A. Balkema, Cape Town, pp 98–160
Huyser O, Ryan PG, Cooper J (2000) Changes in population size, habitat use and breeding biology of lesser sheathbills (Chionis minor) at Marion Island: impacts of cats, mice and climate change? Biol Conserv 92:299–310
Imber MJ, Bell BD, Bell EA (2005) Antipodes Island birds in Autumn 2001. Notornis 52:125–132
Jansen van Vuuren B, Chown SL (2007) Genetic evidence confirms the origin of the house mouse on Subantarctic Marion Island. Polar Biol 30:327–332
Jones AG, Chown SL, Gaston KJ (2002) Terrestrial invertebrates of Gough Island: an assemblage under threat. Afr Entomol 10:83–91
Jones AG, Chown SL, Gaston KJ (2003a) Introduced house mouse as a conservation concern on Gough Island. Biol Conserv 12:2107–2119
Jones AG, Chown SL, Ryan PG, Gremmen NJM, Gaston KJ (2003b) A review of conservation threats on Gough Island: a case study for terrestrial conservation in the Southern Oceans. Biol Conserv 113:75–87
Jones HP, Tershy BR, Zavaleta ES, Croll DA, Keitt BS, Finkelstein ME, Howald GR (2008) Severity of the effects of invasive rats on seabirds: a global review. Conserv Biol 22:16–26
King CM (1982) Age structure and reproduction in feral New Zealand populations of the house mouse (Mus musculus) in relation to seedfall of southern beech. N Z J Zool 9:467–480
King CM (1990) The handbook of New Zealand mammals. Oxford University Press, Auckland
Le Roux V, Chapuis J-L, Frenot Y, Vernon P (2002) Diet of the house mouse (Mus musculus) on Guillou Island, Kerguelen Archipelago, Subantarctic. Polar Biol 25:49–57
MacKay JWB, Russell JC, Murphy EC (2008) Eradicating mice from islands: successes, failures and the way forward. In: Witmer GW, Pitt CW, Fagerstone KA (eds) Managing vertebrate invasive species: proceedings of an international symposium. USDA/APHIS/WS, National Wildlife Research Center, Fort Collins, pp 294–304
Maron JL, Estes JA, Croll DA, Danner EM, Elmendorf SC, Buckelew SL (2006) An introduced predator alters Aleutian island plant communities by thwarting nutrient subsidies. Ecol Monogr 76:3–24
Marris JWM (2000) The beetle (Coleoptera) fauna of the Antipodes Islands, with comments on the impact of mice; and an annotated checklist of the insect and arachnid fauna. J R Soc N Z 30:169–195
Marris E (2005) Mice gang up on endangered birds. Nature News. www.nature.com/news/2005/050718/full/050718–2.html
McIntosh AR (2001) The impact of mice on the Antipodes Islands. In: Southland Conservancy (ed) Antipodes Island expedition, October–November 1995. Department of Conservation, Dunedin, pp 52–57
Micol T, Jouventin P (2002) Eradication of rats and rabbits from Saint-Paul Island, French Southern Territories. In: Veitch CR, Clout MN (eds) Turning the tide: the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, IUCN, Gland, pp 199–205
Miller CJ, Miller TK (1995) Population dynamics and diet of rodents on Rangitoto Island, New Zealand, including the effect of a 1080 poison operation. N Z J Ecol 19:19–27
Millius S (2007) Killer mice hit seabird chicks. Sci News 171:269
Miskelly CM, Walker KJ, Elliott GP (2006) Breeding ecology of three Subantarctic snipes (genus Coenocorypha). Notornis 53:361–374
Moors PJ, Atkinson IAE (1984) Predation on seabirds by introduced animals and factors affecting its severity. In: Croxall JP, Evans PGH, Schreiber RW (eds) Status and conservation of the world’s seabirds. International Council for Bird Preservation, Technical Publication No.2, Cambridge, UK, pp 667–690
Mulder CPH, Grant-Hoffman MN, Towns DR, Bellingham PJ, Wardle DA, Durret MS, Fukami T, Bonner KI (2008) Direct and indirect effects of rats: does rat eradication restore ecosystem functioning of New Zealand seabird islands? Biol Inv. doi:10.1007/s10530--008-9396-x
Nogales M, Martin A, Tershy BR, Donlan CJ, Veitch D, Puerta N, Wood B, Alonso J (2004) A review of feral cat eradication on islands. Conserv Biol 18:310–319
Parkes JP, Murphy EJ (2003) Management of introduced mammals in New Zealand. N Z J Zool 30:335–359
Patrick B (1994) Antipodes Island Lepidoptera. J R Soc N Z 24:91–116
Pearce F (2006) Out of sight, out of luck. New Sci 192:10
Peters RH (1983) The ecological implications of body size. Cambridge University Press, Cambridge
Poiani KA, Richter BD, Anderson MG, Richter HE (2000) Biodiversity conservation at multiple scales: functional sites, landscapes, and networks. Bioscience 50:133–146
Pye T, Swain R, Seppelt RD (1999) Distribution and habitat use of the feral black rat (Rattus rattus) on Subantarctic Macquarie Island. J Zool Lond 247:238–429
Quammen D (1996) The song of the Dodo: island biogeography in an age of extinctions. Scribner, New York
Quillfeldt P, Schenk I, McGill RAR, Strange IJ, Masello JF, Gladbach A, Roesch V, Furness RW (2008) Introduced mammals coexist with seabirds at New Island, Falkland Islands: abundance, habitat preferences, and stable isotope analysis of diet. Polar Biol 31:333–349
Richardson ME (1984) Aspects of the ornithology of the Tristan da Cunha group and Gough Island. Cormorant 12:122–201
Rowe FP, Taylor EJ, Chudley AHJ (1964) The effect of crowding on the reproduction of the house-mouse (Mus musculus L.) living in corn-ricks. J Anim Ecol 33:477–483
Rowe-Rowe DT, Green B, Crafford JE (1989) Estimated impact of feral house mice on Subantarctic invertebrates at Marion Island. Polar Biol 9:457–460
Ruscoe WA, Elkinton JS, Choquenot D, Allen RB (2005) Predation of beech seed by mice: effects of numerical and functional responses. J Anim Ecol 74:1005–1019
Russell JC, Clout MN (2004) Modelling the distribution and interaction of introduced rodents on New Zealand offshore islands. Glob Ecol Biogeogr 13:497–507
Ryan PG (2005) Inaccessible Island seabird monitoring manual. RSPB research report no. 16. Royal Society for the Protection of Birds, Sandy
Ryan PG, Glass JP (2001) Inaccessible Island nature reserve management plan. Government of Tristan da Cunha, Edinburgh
Shirihai H (2002) A complete guide to Antarctic wildlife: the birds and marine mammals of the Antarctic Continent and Southern Ocean. Alula Press, Oy
Simberloff D (2008) Rats aren’t the only introduced rodents producing ecosystem impacts on islands. Biol Inv. doi:10.1007/s10530-008-9400-5
Singleton GR, Krebs CJ, Davis S, Chambers L, Brown P (2001) Reproductive changes in fluctuating house mouse populations in southeastern Australia. Proc R Soc Lond B 268:1741–1748
Singleton GR, Brown PR, Pech RP, Jacob J, Mutze GJ, Krebs CJ (2005) One hundred years of eruptions of house mice in Australia—a natural biological curio. Biol J Linn Soc 84:617–627
Smith VR (1978) Animal-plant-soil nutrient relationships on Marion Island (Subantarctic). Oecologia 32:239–253
Smith VR (1979) The influence of seabird manuring on the phosphorus status of Marion Island (Subantarctic) soils. Oecologia 41:123–126
Smith VR (2002) Climate change in the Subantarctic: an illustration from Marion Island. Clim Change 52:345–357
Smith VR, Steenkamp M (1990) Climatic change and its ecological implications at a Subantarctic island. Oecologia 85:14–24
Smith VR, Steenkamp M (1992) Soil macrofauna and nitrogen on a Subantarctic island. Oecologia 92:201–206
Smith VR, Steenkamp M (1993) Macro-invertebrates and peat nutrient mineralization on a Subantarctic island. S Afr J Bot 59:106–108
Smith VR, Avenant NL, Chown SL (2002) The diet and impact of house mice on a Subantarctic island. Polar Biol 25:703–715
Taylor RH (1979) How the Macquarie Island parakeet became extinct. N Z J Ecol 2:42–45
Thompson FR, Burhans DE (2004) Differences in predators of artificial and real songbird nests: evidence of bias in artificial nest studies. Conserv Biol 18:373–380
Torr N (2002) Eradication of rabbits and mice from Subantarctic Enderby and Rose islands. In: Veitch CR, Clout MN (eds) Turning the tide: the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, World Conservation Union, Gland, pp 319–328
Towns DR, Broome KG (2003) From small Maria to massive Campbell: forty years of rat eradications from New Zealand. N Z J Zool 30:377–398
Traveset A, Nogales M, Alcover JA, Delgado JD, López-Darias M, Igual JM, Godoy D, Bover P (2008) A review on the effects of alien rodents in the Balearic (western Mediterranean Sea) and Canary Islands (eastern Atlantic Ocean). Biol Inv. doi:10.1007/s10530-008-9395-y
van Aarde R, Jackson T (2006) Food, reproduction and survival in mice on Subantarctic Marion Island. Polar Biol 30:503–511
Vári L (1971) Lepidoptera (Heterocera: Tineidae, Hyponomeutidae). In: van Zinderen Bakker Sr EM, Winterbottom JM, Dyer RA (eds) Marion and Prince Edward Islands. Report on the South African biological and geological expedition/1965–1966. A.A. Balkema, Cape Town, pp 349–354
Veitch CR, Clout MN (2002) Turning the tide: the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, IUCN, Gland
Wace NM (1986) The arrival, establishment and control of alien plants on Gough Island. S Afr J Antarct Res 16:95–101
Wace NM, Dickson JH (1965) The terrestrial botany of the Tristan da Cunha Islands. Part II. The biological report of the Royal Society expedition to Tristan da Cunha, 1962. Philos Trans R Soc Lond B 249:273–360
Wanless RM (2007) Invaders of the last ark. Afr Birds Birding 12(4):55–59
Wanless RM, Angel A, Hilton GM, Ryan PG (2005) Cultural evolution in the introduced house mouse: evidence for the cultural transmission of a unique predatory behaviour on Gough Island? Paper presented at the Society for Conservation Biology Conference, Brasilia, Brazil, 15–19 July 2005
Wanless RM, Angel A, Cuthbert RJ, Hilton G, Ryan PG (2007) Can predation by invasive mice drive seabird extinctions? Biol Lett 3:241–244
Williamson M (1996) Biological invasions. Chapman & Hall, London
Witmer GW, Boyd F, Hillis-Starr Z (2007) The successful eradication of introduced roof rats (Rattus rattus) from Buck Island using diphacinone, followed by an irruption of house mice (Mus musculus). Wildl Res 34:108–115
Woodward PW (1972) The natural history of Kure Atoll, northwestern Hawaiian Islands. Atoll Res Bull 164:1–318
Zavaleta ES (2002) It’s often better to eradicate, but can we eradicate better? In: Veitch CR, Clout MN (eds) Turning the tide: the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, IUCN, Gland, pp 393–434
Zavaleta ES, Hobbs RJ, Mooney HA (2001) Viewing invasive species removal in a whole-ecosystem context. Trends Ecol Evol 16:454–459
Acknowledgments
John Parkes, Peter Ryan and two anonymous reviewers provided valuable comments on earlier versions of this manuscript. Part of the research and literature review was funded by the UK Overseas Territories Environment Programme. RMW is supported by the University of Cape Town, the (South African) National Research Foundation and the Royal Society for the Protection of Birds.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Angel, A., Wanless, R.M. & Cooper, J. Review of impacts of the introduced house mouse on islands in the Southern Ocean: are mice equivalent to rats?. Biol Invasions 11, 1743–1754 (2009). https://doi.org/10.1007/s10530-008-9401-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-008-9401-4