
Abstract
The diet of the Iberian otter (Lutra lutra) was determined by analysing 547 spraints collected at 28 sites within a wide area invaded by centrarchid fishes (pumpkinseed sunfish, Lepomis gibbosus and largemouth bass, Micropterus salmoides): the middle Guadiana basin (South-west Iberian Peninsula). Fish was the otters’ main prey, representing more than 60% of total individuals and more than 80% of total biomass. Otters preyed on most of the fish species captured in the field; however, the consumption of centrarchids was low compared to their abundance in the streams, and Jacobs’ index of preference showed a clear rejection of both species by the otter. Consumption of native fish genera (Squalius, Barbus and Chondrostoma) by otters increased in relation to their increase in the environment. In contrast, increasing numbers of L. gibbosus in the field was not reflected in otter consumption. The general decline of native freshwater fishes in Iberian rivers, the preferred prey of otters, together with the spread of exotic fish species (centrarchids and others) could put otter populations at risk.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The otter (Lutra lutra, Mustelidae) is a top predator in freshwater ecosystems. Fish are the otter’s main prey, but its opportunistic feeding behaviour allows it to feed on whatever prey is available (Mason and Macdonald 1986; Clavero et al. 2003). Exotic fish species are increasing their range in Iberian freshwater habitats (Clavero and Garcia-Berthou 2006), and among them, two centrarchids (pumpkinseed sunfish, Lepomis gibbosus, and largemouth bass, Micropterus salmoides) are among the most widespread species, specially in the Guadiana river basin (Bernardo et al. 2003). These fish species are potential threats to the Iberian native fish fauna and are the dominant fish species in many areas (Godinho and Ferreira 2000; Doadrio 2002; Blanco-Garrido 2006). Although the pumpkinseed sunfish was probably introduced into the Guadiana basin in the late 1970s (Almaça 1995), the first record of the species in the area was in 1984 (López and de la Cruz 1985). The introduction of largemouth bass into the basin predates that of pumpkinseed (Almaça 1965). The first record of the species in our study area is not available, but it was probably widespread at the time the pumpkinseed was introduced.
The question arises as to whether this new potential trophic resource is being consumed by native predators that largely depend on fish for their food. It is possible that otters from the Iberian Peninsula could benefit from the introduction of these exotic species, given apparent increased biomass available for food. This is what happened with the introduction and expansion of the red-swamp crayfish (Procambarus clarkii). Crayfish was introduced to Spain in 1973/1974, spread rapidly in most Iberian rivers (Hasburgo-Lorena 1983), and became important prey for otters and other predators (Adrián and Delibes 1987; Delibes and Adrián 1987; Correia 2001). Moreover, the recovery of the otter in Iberian Peninsula, after the decline suffered by the species in the 1970s, was apparently related in part to the introduction of crayfish (Ruiz-Olmo and Delibes 1998).
Information on consumption of centrarchids by otters from the Iberian Peninsula is very scarce and mainly refers to largemouth bass, which is less than 5% by frequency of occurrence in otter diets from running waters (López-Nieves and Casal 1984; Morales et al. 2004). Their contribution tends to be slightly higher in reservoirs (López-Nieves and Casal 1984; Pedroso and Santos-Reis 2006). However, no work addresses explicitly the relative importance of these invasive species in otter diets and the selectivity displayed by otters for them.
In this paper, we describe the otter diet in a wide area invaded by centrarchid fishes within the middle Guadiana basin. Our main objective was to evaluate the importance of both invasive species in otter diet. For this purpose, we analysed the selectivity displayed by otters for various fish species inhabiting the area.
Methods
Study area
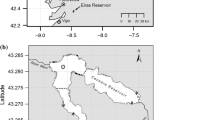
The field work was carried out in 28 localities distributed along the Guadiana basin, between Táliga and Malagón streams (Fig. 1). The area is characterised by a Mediterranean climate with seasonal flooding (autumn, winter and early spring) and severe droughts (summer) that vary markedly between years. The mean annual precipitation is about 600 mm. This climate regime strongly affects the hydrological network, causing high intra- and inter-annual variability in the flow regime. In summer, under average climate conditions, most tributaries become intermittent with a succession of pools of variable dimensions with little or no water flow. More permanent and stable hydrological conditions exist in downstream stretches or in the main river.

Map of the study area showing sampling sites in the middle Guadiana river basin
The Mediterranean climate regime produces high exploitation of water resources (Collares-Pereira et al. 2000). About 3 of the 5 tributaries we investigated are affected by the recent Alqueva Reservoir (dam closed at February 2002, Fig. 1), the largest European artificial lake. The Chanza river is flooded by two reservoirs, Chanza and the recent Andévalo Reservoir (dam closed at January 2004, Fig. 1).
In general, the human population density in the study area is moderate to low (mean 43 inhabitants/km2, range 3–320 inhabitants/km2). Farming is the main economic activity and large areas are still covered by pastures and natural forests.
Analysis of otter diet
The diet of the otter was determined by analysis of its faeces (referred to as ‘spraints’ hereafter), that were collected from April to June 2001 (prior to the filling of Alqueva and Andévalo dams). Spraints contain those parts of the food intake that cannot be digested (bones, scales, hair and feathers). We analysed a total of 547 spraints (Table 1). On average, 20 spraints were collected per site, and diet analyses followed standard methodology (Beja 1997). Prey remains were identified using published works (Webb 1980; Roselló 1986; Prenda et al. 2002) and our own collection for comparison. Each identified prey class was considered an ‘occurrence’. The relative frequency of occurrence (RFO) was the percentage of the total number of occurrences corresponding to a certain prey class (Mason and Macdonald 1986).
The minimum number of individuals of each prey class present in a spraint was estimated by the number and position (left–right) of diagnostic hard parts (mainly mouth bones for fish and endopodites/exopodites for the crayfish, Procambarus clarkii), which were measured with a calliper to the nearest 0.1 mm. In cases where these diagnostic pieces were not found, the remains of the prey item were considered to belong to a single individual.
We applied regression equations to estimate the original size and weight of the prey consumed by the otters. In the case of fish, regressions between the size of key bones and original length (Prenda et al. 2002), and between length and weight (Prenda et al. 2002; own unpublished data) were computed. The original size of ingested crayfish was estimated using a regression between the size of the endopodite/exopodite and the total length, computed from a crayfish sample (unpublished data). The weight of crayfish was estimated by regression between total length and crayfish weight without claws, which were rarely found in otter spraints.
The lengths and weights of fish and crayfish individuals without measured key hard parts were distributed following the frequencies of lengths and weights in the estimated samples of each prey class. Constant weights were assigned to the remaining prey classes: insects, 1 g; Rana perezi, 10 g; Bufo bufo, 50 g; unidentified amphibians, 10 g; Natrix sp., 50 g; Mauremys leprosa, 100 g; birds, 100 g; small mammals, 100 g (Beja 1996).
Prey selection
To search for possible preference for or avoidance of centrarchids (pumpkinseed sunfish and largemouth bass) by the otter, we studied otter prey selection focused on fish (Fig. 1). In each sampled site, the relative availability of fish prey was estimated by electrofishing. Fish sampling was done for 100–250 m stream length during 1–1.5 h. Given the high temporal variability in community composition exhibited by Iberian fishes (see Filipe et al. 2002), fish sampling and spraint collection were carried out simultaneously. All captured fish were identified and measured for total length, and then returned to the water. We applied regression equations between fish length and weight (Prenda et al. 2002, own unpublished data) to estimate the available fish biomass. Fish species that appeared rarely in the otter diet or in the environment were removed from the analysis of prey selection (Table 1). In many cases, it was difficult to assign fish remains to species that coexisted with congeneric species (Squalius, Barbus and Chondrostoma), due to the similarity of their bones, so congeneric fish species were pooled into a single prey item. Six fish prey categories were used in the analysis: Squalius, Barbus, Chondrostoma, Cobitis paludica, Lepomis gibbosus and Micropterus salmoides.
To obtain an absolute preference or avoidance value for each fish prey category, we used Jacobs’ (1974) selection index, calculated as:
where r is the proportion of the prey category in the diet and p is the proportion in the environment. The index ranges from −1 to +1, −1 being total avoidance, 0 no preference and +1 full preference. D was calculated for each prey category in each site. To interpret the index objectively, we compared alternative hypotheses, in which the null hypothesis corresponded with a Jacobs’ index equal to zero (the prey is used as available in the environment). Values were compared using Student’s t-test (Revilla et al. 2000).
Finally, Pearson correlation was carried out between fish prey categories in the environment and in otter diet. This analysis allowed us to describe changes in the otter diet that could be related to the variation of fish prey in the environment.
Results
Spraint analysis showed that fish was the otter’s main prey, representing more than 60% of total individuals and more than 80% of total consumed biomass (Table 1). Within fish, Barbus spp. were the main prey item, representing 29.5% of total individuals and 67.7% of total consumed biomass in the entire study area (Table 1). Crayfish was the second main prey item (21.5% of individuals and 8.2% of biomass, Table 1). Other categories can be considered as of minor importance.
Otters consumed most of the fish species captured in the field (Table 1). However, Anaecypris hispanica, Gambusia holbrooki and Australoheros facetus were captured during the field work but were absent from otter spraints. No eels (Anguilla anguilla) were captured during fish sampling, but one individual was detected in an otter spraint.
Predation on amphibians (mainly Rana perezi) was frequent, while birds, reptiles and mammals were rather rare in the otters’ diet (Table 1).
Using Jacob’s preference index for individuals, Barbus was the only preferred fish prey (mean D = 0.42), while centrarchids were avoided (pumpkinseed sunfish: mean D = −0.40; largemouth bass: mean D = −0.66) and the remainder were used as available (Fig. 2). Similar results were obtained for biomass; Barbus was again the only preferred prey (mean D = 0.34). Both centrarchids (pumpkinseed: mean D = −0.61; largemouth bass: mean D = −0.84) and Squalius (mean D = −0.43) were avoided (Fig. 2).

Jacobs’ index for each fish prey consumed by the otter. The index was applied to individuals and biomass consumed. Positive and negative values indicate preference and avoidance, respectively. Sq—Squalius sp., Ba—Barbus sp., Ch—Chondrostoma sp., Co—Cobitis paludica, Le—Lepomis gibbosus and Mi—Micropterus salmoides. Bars are the standard error. * mean is significantly different from zero (t-Student, P < 0.05)
There was a positive correlation between consumption of Squalius and Barbus and abundance of these prey in the environment (Fig. 3). This pattern was partially followed by Chondrostoma (Pearson correlation, r = 0.34, P = 0.08). However, there was no such change in the consumption of pumpkinseed sunfish; that is, increasing numbers of this species in the field were not reflected in diets (Fig. 3). The response of the otter to the increase of largemouth bass in the field could not be adequately addressed because the otter consumed this prey in only two locations.

Pearson correlation between the percentage of individuals of Squalius sp., Barbus sp., and pumpkinseed sunfish (Lepomis gibbosus) present in the environment and in otter diet
Discussion
Despite the recognized opportunistic trophic behaviour of otters, in our study area they showed a distinct preference for barbels (Barbus spp.) in their diets. Barbels accounted for the largest proportion of individuals and biomass consumed. This finding fits well with observations from other Iberian basins (Ruiz-Olmo et al. 2001). Both pumpkinseed sunfish and largemouth bass were clearly rejected, appearing in a significantly lower proportion in the diet than in the environment. Similarly, the consumption of centrarchids by otters was extremely low, as in other Iberian basins (Morales et al. 2004) and other areas (Lanski and Molnar 2003). Other fish predators, such as the grey heron (Ardea cinerea), have incorporated pumpkinseed in their diet (Peris et al. 1995), although consumption is low compared with other fish prey species.
The mechanisms involved in this avoidance pattern may be related to the anti-predator body structures of the two centrarchids. Pumpkinseed sunfish possesses a tough skin and sharp spines in the dorsal, pectoral and anal fins. Similarly, the largemouth bass has spines in its fins and a strong, spiny opercle. These morphological features impede an easy handling and capture. Heggberget and Moised (1994) found that, relative to their number in traps, spiny fishes with tough skin were rare in otter spraints. In general, otters prefer cyprinids to spiny-rayed percids (Mason and MacDonald 1986; Taastrøm and Jacobsen 1999; Jacobsen 2005). Cyprinids are also preferred over percids by other fish predators, such as pike (Esox lucius) (Eklöv and Hamrin 1989). Not only fish with hard structures may be rejected by otters, but also other prey with such features. Adrian and Delibes (1987) suggested that the Spanish ribbed newt (Pleurodeles waltl) was not consumed by otters because of its external sharp-pointed ribs. Anti-predator body structures do not exist in Iberian native freshwater fishes. Certainly, the absence of the cichlid Australoheros facetus in otter diet, another exotic species, also supports this hypothesis; it has the same body shape and features as pumpkinseed. Other invasive species, such as the red-swamp crayfish, that are consumed by otters do not possess these anti-predator structures.
Not surprisingly, the increase of pumpkinseed in the environment was not followed by a similar increase in otter diets. Pumpkinseed seem to be avoided or ignored by otters. It is possible that the time since the centrarchid invasion has not been long enough to allow otters to develop effective mechanisms to capture and handle them. In fact, otter species from areas where centrarchids are native consume them in a high proportion (Sheldon and Toll 1964). Although with our data we could not fit a clear functional response related to pumpkinseed consumption by otters, they could be displaying a type III functional response in its first steps, suggestive of a learning process. The type III response occurs in active predators which increase their search activity with increasing prey density (Holling 1959). This is typical of generalist carnivores for whom alternative prey are available (Revilla and Palomares 2002). Otters showed a wide trophic spectrum in the study area, consuming at least 13 fish species, aquatic invertebrates, amphibians, reptiles, birds and small mammals (Table 1). This great variety of alternative prey might allow otters to reject pumpkinseed and largemouth bass (Erlinge 1968).
In the Iberian Peninsula, there is a general decline of native fish fauna (Aparicio et al. 2000; Doadrio 2002), traditionally the preferred prey of otters (Prenda and Granado-Lorencio 1996), together with an increase of exotic fish species (Clavero et al. 2004). The increasing and dominance of exotic fish species, centrarchids and others, may force otters to consume them in a higher proportion than they do at present. In fact, Lutra lutra is known to prey highly on pumkinseed in reservoirs (Pedroso and Santos-Reis 2006), where exotic fishes are dominant (Clavero et al. 2004). However, the acclimation of Iberian otters to centrarchids does not necessarily mean that their populations can persist by feeding on them. Indeed, Kruuk and Goudswaard (1990) found that, though the spotted-necked otter (Lutra maculicollis) changed its diet following the introduction of exotic fish (Nile perch, Lates niloticus and others) in Lake Victoria, its population declined in most of the lake’s coasts.
If centrarchids and other potential exotic fish prey become dominant in the streams of the Iberian peninsula, the maintenance of healthy populations of Iberian otters could be at risk. The monitoring of fish communities and the analysis of the diet and habitat use/ occupancy by otters in areas affected by exotic fishes are needed to clarify the long term response of otters to changes in fish community composition.
References
Adrián MI, Delibes M (1987) Food habits of the otter (Lutra lutra) in two habitats of the Doñana National Park, SW Spain. J Zool Lond 212:399–406
Almaça C (1965) Contribution à la connaisance des poissons d’eaux interieures du Portugal. Arquiv Mus. Bocage 1:9–39
Almaça C (1995) Fish species and varieties introduced into Portuguese inland waters. Lisboa: Publ. Avuls Mus. Bocage
Aparicio E, Vargas MJ, Olmo JM, de Sostoa A (2000) Decline of native freshwater fishes in a Mediterranean watershed on the Iberian Peninsula: a quantitative assessment. Environ Biol Fishes 59:11–19
Beja PR (1996) An analysis of otter Lutra lutra predation on introduced American crayfish Procambarus clarkii in Iberian streams. J Appl Ecol 33:1156–1170
Beja PR (1997) Predation by marine-feeding otters (Lutra lutra) in South-west Portugal in relation to fluctuating food resources. J Zool Lond 242:503–518
Bernardo JM, Ilhéu M, Matono P, Costa AM (2003) Interannual variation of fish assemblage structure in a Mediterranean river: implications of stream flow on the dominance of native or exotic species. Riv Res Appl 19:521–532
Blanco-Garrido F (2006) Ecología, distribución y conservación de peces continentales en el cuadrante suroccidental ibérico. Ph D Thesis. Universidad de Huelva
Clavero M, Garcia-Berthou E (2006) Homogenization dynamics and introduction routes of invasive freshwater fish in the Iberian Peninsula. Ecol Appl 16:2313–2324
Clavero M, Blanco-Garrido F, Prenda J (2004) Fish fauna in Iberian Mediterranean river basins: biodiversity, introduced species and damming impacts. Aquat Conserv Mar Freshw Ecosyst 14:575–585
Clavero M, Prenda J, Delibes M (2003) Trophic diversity of the otter (Lutra lutra L) in temperate and Mediterranean freshwater habitats. J Biog 30:761–769
Collares-Pereira MJ, Cowx IG, Ribeiro F, Rodrigues JA, Rogado L (2000) Threats imposed by water resource development schemes on the conservation of endangered fish species in the Guadiana river Basin in Portugal. Fish Manag Ecol 7:167–178
Correia AM (2001) Seasonal and interspecific evaluation of predation by mammals and birds on the introduced red swamp crayfish Procambarus clarkii (Crustacea, Cambaridae) in a freshwater marsh (Portugal). J Zool Lond 255:533–541
Delibes M, Adrián MI (1987) Effects of crayfish introduction on otter Lutra lutra food in the Doñana National Park, SW Spain. Biol Conserv 42:153–159
Doadrio I (ed) (2002) Atlas y libro rojo de los peces continentales de España. Dirección General de Conservación de la Naturaleza. 2nd edn
Eklöv P, Hamrin SF (1989) Predatory efficiency and prey selection: interactions between pike Esox lucius, perch Perca fluviatilis and rudd Scardinius erythrophthalmus. Oikos 56:149–156
Erlinge S (1968) Food studies on captive otters (Lutra lutra L.). Oikos 19:259–270
Filipe AF, Cowx IG, Collares-Pereira MJ (2002) Spatial modelling of freshwater fish in semi-arid river systems: a tool for conservation. River Res Appl 18:123–136
Godinho FN, Ferreira MT (2000) Composition of endemic fish assemblages in relation to exotic species and river regulation in a temperate stream. Biol Invasions 2:231–244
Hasburgo-Lorena AS (1983) The status of the Procambarus clarkii population in Spain. Freshw Crayfish 6:131–133
Heggberget TM, Moised KE (1994) Prey selection in coastal Eurasian otters Lutra lutra. Ecography 17:331–338
Holling CS (1959) Some characteristics of simple types of predation and parasitism. Can Entomol 91:385–398
Jacobs J (1974) Quantitative measurement of food selection. A modification of the forage ratio and Ivlevs electivity index. Oecologia 14:413–417
Jacobsen L (2005) Otter (Lutra lutra) predation on stocked brown trout (Salmo trutta) in two Danish lowland rivers. Ecol Fresh Fish 14:59–68
Kruuk H, Goudswaard PC (1990) Effects of changes in fish populations in Lake Victoria on the food of otters (Lutra maculicollis Schinz and Aonyx capensis Lichtenstein). African J Ecol 28:322–329
Lanski J, Molnar T (2003) Diet of otters living in three different habitats in Hungary. Folia Zool 52:378–388
de López F, de la Cruz C (1985) Lepomis gibbous L. (Perciformes, Centrarchidae) nueva especie en la ictiofauna del Guadiana. Doñana Acta Vertebrata 12(1):165–166
López-Nieves P, Casal H (1984) Food habits of the otter in central Sierra Morena (Córdoba, Spain). Acta Theriol 29:383–401
Mason CF and Macdonald SM (1986) Otters: ecology and conservation. Cambridge University Press
Morales JJ, Lizana M, Acera F (2004) Ecología trófica de la nutria paleártica (Lutra lutra) en el río Francia (cuenca del Tajo, Salamanca). Galemys 16:57–77
Pedroso NM, Santos-Reis M (2006) Summer diet of Eurasian otters in large dams of south Portugal. Hystrix 17:117–128
Peris SJ, Briz FJ, Campos F (1995) Shifts in the diet of the grey heron (Ardea cinerea) in the Duero basin, central-west Spain, following the introduction of exotic fish species. Folia Zool 44:97–102
Prenda J, Granado-Lorencio C (1996) The relative influence of riparian habitat structure and fish availability on otter Lutra lutra L. sprainting activity in a small Mediterranean catchment. Biol Conserv 76:9–15
Prenda J, Arenas MP, Freitas D, Santos-Reis M, Collares-Pereira MJ (2002) Bone length of Iberian freshwater fish, as predictor of length and biomass of prey consumed by piscivorous. Limnética 21:15–24
Revilla E, Palomares F (2002) Does local feeding specialization exist in Eurasian badgers? Can J Zool 80:83–93
Revilla E, Palomares F, Delibes M (2000) Defining key habitats for low density populations of Eurasian badgers in Mediterranean environments. Biol Conserv 95:269–277
Roselló E (1986) Atlas osteológico de los teleósteos ibéricos. I. Mandíbula inferior (dentario y articular). Tesis de Licenciatura. UAM, Madrid
Ruiz-Olmo J and Delibes M (1998) La nutria en España ante el horizonte del año 2000. SECEM
Ruiz-Olmo J, López-Martín JM, Palazón S (2001) The influence of fish abundance on the otter (Lutra lutra) populations in Iberian Mediterranean habitats. J Zool Lond 254:325–336
Sheldon WG, Toll WG (1964) Feeding habits of the river otter in a reservoir in central Massachusetts. J Mammal 45:449–455
Taastrøm HM, Jacobsen L (1999) The diet of otters (Lutra lutra L.) in Danish freshwater habitats: comparisons of prey fish populations. J Zool Lond 248:1–13
Webb JB (1980) Otter spraint analysis. Occasional Publication, Mammal Society, London
Acknowledgements
We greatly acknowledge Miguel Clavero, Antonia Rebollo and Jerónimo Valle for field and lab assistance. Professor Peter B. Moyle provided very useful comments and suggestions on the edition of the manuscript. Dr. Drake and two anonymous referees made very valuable suggestions to early versions of the manuscript. This study was financially supported by the SECEM (Sociedad Española para la Conservación y Estudio de los Mamíferos) and the Spanish Government projects REN2002-03513/HID (Ministerio de Ciencia y Tecnología) and CGL2005-02699/HID (Ministerio de Educación y Ciencia).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Blanco-Garrido, F., Prenda, J. & Narvaez, M. Eurasian otter (Lutra lutra) diet and prey selection in Mediterranean streams invaded by centrarchid fishes. Biol Invasions 10, 641–648 (2008). https://doi.org/10.1007/s10530-007-9158-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-007-9158-1