
Abstract
Objective
The Δ9-tetrahydrocannabinolic acid synthase (THCAS) from Cannabis sativa was expressed intracellularly in different organisms to investigate the potential of a biotechnological production of Δ9-tetrahydrocannabinolic acid (THCA) using whole cells.
Results
Functional expression of THCAS was obtained in Saccharomyces cerevisiae and Pichia (Komagataella) pastoris using a signal peptide from the vacuolar protease, proteinase A. No functional expression was achieved in Escherichia coli. The highest volumetric activities obtained were 98 pkat ml−1 (intracellular) and 44 pkat ml−1 (extracellular) after 192 h of cultivation at 15 °C using P. pastoris cells. Low solubility of CBGA prevents the THCAS application in aqueous cell-free systems, thus whole cells were used for a bioconversion of cannabigerolic acid (CBGA) to THCA. Finally, 1 mM (0.36 g THCA l−1) THCA could be produced by 10.5 gCDW l−1 before enzyme activity was lost.
Conclusion
Whole cells of P. pastoris offer the capability of synthesizing pharmaceutical THCA production
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
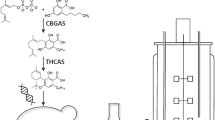
Since the discovery of the psychoactive cannabinoid, Δ9-tetrahydrocannabinol (THC), from Cannabis sativa in 1964, and the human endocannabinoid system, the effects and potential pharmaceutical applications of THC have been extensively studied. Nowadays, THC is widely used as therapeutic agent in treatment of chemotherapy-associated nausea and vomiting, AIDS-related loss of appetite as well as pain and muscle spasms in multiple sclerosis. Further applications are still under investigation and the demand of pharmaceutical grade THC is still increasing (Carlini 2004; Pertwee 2006). Nevertheless, legal regulations for the cultivation of C. sativa in most countries, chemical syntheses of THC with low yields or expensive, chiral precursors are drawbacks for a cost-effective THC production (Mechoulam 1970; Trost and Dogra 2007). Therefore a biotechnological approach transferring the enzymes of the plant biosynthetic pathway of Δ9-tetrahydrocannabinolic acid (THCA), the precursor of THC, into a microbial production host might present an appropriate alternative. The enzyme, Δ9-tetrahydrocannabinolic acid synthase (THCAS) catalyzing the last step of THCA biosynthesis (Fig. 1), the oxidative cyclization of cannabigerolic acid (CBGA), has been expressed in low amounts in recombinant tobacco hairy roots, insect cell cultures and secreting Pichia (Komagataella) pastoris cultures (Sirikantaramas et al. 2004; Taura et al. 2007). Furthermore, a crystal structure of the enzyme has indicated a covalently-bound FAD moiety (bound to His114 and Cys176), a disulfide bond (between Cys37 and Cys99) and six N-linked glycosylation sites (Shoyama et al. 2012). Additionally, a catalytic mechanism of the oxidation of CBGA to THCA has been proposed, including H2O2 formation for the regeneration of FAD during catalysis.

Biosynthetic pathway of Δ9-tetrahydrocannabinolic acid (THCA) biosynthesis in Cannabis sativa. Cannabigerolic acid (CBGA) and THCA are decarboxylated non-enzymatically by heat. Modified from Flores-Sanchez and Verpoorte (2008)
Since a scale-up approach with the isolated THCAS is not suitable due to: (i) the low water solubility of the substrate CBGA, (ii) the denaturing properties of H2O2, and (iii) the integral membrane enzyme CBGA synthase, that cannot be secreted and therefore not feasibly implemented into a cell-free production system, we focused in this study on the intracellular THCAS expression in Escherichia coli, Saccharomyces cerevisiae and P. pastoris cells and the possible application towards a whole cell production system of THCA.
Materials and methods
Chemicals
Δ9-THCA was purchased from THC Pharm (Frankfurt am Main, Germany). Cannabigerolic acid (CBGA) was purchased from Taros Chemicals (Dortmund, Germany).
Microorganisms, genes and plasmids
Cloning strategies and a detailed list of all strains and plasmids used in this study are given in the supplementary material (Supplementary Tables 1, 2). Synthetic coding sequences of THCAS (GenBank accession number AB057805) were codon optimized for expression in Saccharomyces cerevisiae and Pichia pastoris and purchased without signal peptide from GeneArt (Regensburg, Germany). Recombinant expression of THCAS was conducted with the following microorganisms: E. coli SHuffle T7 Express and SHuffle T7 Express lysY (NEB, Frankfurt am Main, Germany), containing modifications to enable disulfide bond formation in the cytosol, were used with pET28a(+) and pET32a(+) vectors (Merck, Darmstadt, Germany); Saccharomyces cerevisiae CEN.PK2-1CΔgal1 (Euroscarf, Frankfurt am Main, Germany) deficient in the β-galactokinase and CEN.PK2-1CΔgal1Δpep4 (this study) additionally deficient in the vacuolar Proteinase A; Pichia pastoris PichiaPink strains 1, 2 and 3 (Invitrogen, Darmstadt, Germany). All Pichia strains are adenine auxotrophs. Strain 2 and 3 contain additional knockouts of the vacuolar proteases Proteinase A (pep4) and Proteinase B (prb1), respectively. Pichia strains were transformed with linearized high copy (pPink_HC_THCAS) and low copy (pPink_LC_THCAS) vectors for integration into chromosomal TRP2 gene and Saccharomyces strains were transformed with pDionysos_THCAS vector, all containing a cDNA of THCAS with three additional histidines at the C-terminus and an additional N-terminal sequence coding for a 24 aa signal peptide from Proteinase A [UniProt accession number F2QUG8 in pPink vectors (Invitrogen, Darmstadt, Germany) and P07267 in pDionysos vector (Stehle et al. 2008)] for targeting into the cell vacuoles.
Culture conditions
Detailed media compositions are described in Supporting Information. If not stated otherwise, cells were cultivated as follows. Recombinant E. coli cells were grown in 1 l flasks, containing 100 ml LB-medium (50 µg kanamycin ml−1, 33 µg chloramphenicol ml−1, 100 µg spectinomycin ml−1) at 37 °C and 200 rpm to an OD600 of 0.6. THCAS expression was induced by addition of 1 mM IPTG and cells grown for 16 h at 20 °C. Recombinant S. cerevisiae cells were grown in minimal medium without leucine at 30 °C and 200 rpm for 24 h. Cells were used to inoculate 100 ml of 2 × YPAD medium at an OD600 of 0.5 and incubated after induction with 0.5 % (w/v) galactose at 20 °C and 200 rpm for 144 h. Recombinant P. pastoris cells were grown in BMGY at 30 °C and 200 rpm for 24 h. Afterwards, cells were harvested by centrifugation at 5000×g for 5 min and resuspended in modified BMMY (mBMMY) (Taura et al. 2007) to an OD600 of 20. Finally, Pichia cells were cultivated at 15 °C and 200 rpm until no increase in THCAS activity could be observed and supplemented with 0.5 % (v/v) methanol every 24 h for induction of protein expression.
Analytical methods
Cell density was measured from the OD600 value and cell dry mass (CDW) was calculated according to Tolner et al. (2006) with a correlation of CDW (g l−1) = 0.21 g l−1 × OD600. Protein concentrations were measured using Bradford assay (Ernst and Zor 2010). Methanol concentrations were determined by HPLC–UV analysis at 210 nm using Hi-Plex H 300 × 7.7 mm column by isocratic elution (5 mM H2SO4 in H2O) at 0.5 ml min−1 at 65 °C.
THCAS activity assay
A detailed protocol for cell lysis is described in the Supporting Information. Briefly, cells were harvested by centrifugation. Cell pellets were resuspended in 100 mM sodium citrate buffer pH 5.5 and the supernatant was diluted to 50 % (v/v) with 100 mM sodium citrate buffer pH 5.5. Yeast cells were lysed by glass beads and E. coli cells by sonication. After centrifugation of cell debris, lysate supernatants and diluted culture supernatants were used for determination of THCAS activity at 37 °C by addition of CBGA (final concentration 100 µM, 1 % (v/v) DMSO). Activity assays were stopped by addition of 0.3 assay volumes trifluoroacetic acid and 2.7 assay volumes acetonitrile (ACN) followed by incubation on ice for 15 min. Supernatants were analyzed after centrifugation (13,100×g, 4 °C, 30 min) by HPLC using a Nucleosil 100-5 C18 column. Isocratic elution [25 % (v/v) H2O with 0.1 % (v/v) TFA/75 % (v/v) ACN] was used at 0.7 ml min−1. Identification of CBGA and THCA was performed by HPLC–MS and quantification by HPLC–UV at 225 nm and 35 °C.
Results and discussion
Comparison of THCAS activities of recombinant Escherichia coli, Saccharomyces cerevisiae and Pichia pastoris strains
The functional expression of Δ9-tetrahydrocannabinolic acid synthase (THCAS) was investigated using different organisms (Table 1). X-ray structural analysis revealed a disulfide bond. Thus, achieving functional expression in a prokaryotic system, E. coli SHuffle T7 Express and SHuffle T7 Express lysY cells, that are able to establish disulfide bonds in the cytosol, were transformed with pET28a(+)_THCAS, containing the cDNA of THCAS without signal peptide. To rule out solubility issues during expression, cells were also transformed with pET32a(+)_THCAS containing an additional thioredoxin fusion tag for improved solubility. Unfortunately, no expression and activity of THCAS was obtained in E. coli cells. This indicates that functional expression of THCAS might require eukaryotic chaperones able to facilitate covalent binding of FAD to the THCAS or glycosylation of the protein. In both yeast expression systems, the THCAS was targeted into the vacuole of the cell using the signal peptides of the vacuolar proteinase A from S. cerevisiae or P. pastoris, respectively. As vacuolar proteases might degrade THCAS, wild type strains and protease knockout strains were compared with each other. Therefore the PEP4 gene was knocked out by homologous recombination in S. cerevisiae CEN.PK2-1C ∆gal1 (wt). PichiaPink strains 1 (wt), 2 (pep4) and 3 (prb1) were transformed with high and low copy vectors for genome integration. A screening of the different Pichia clones was conducted and is described in the supplementary section in detail. Data indicated that the strain containing the proteinase A knockout transformed with the high copy vector (PP2_HC) showed the highest THCAS activity among P. pastoris and S. cerevisiae strains. Therefore PP2_HC was chosen for subsequent optimization studies, since the obtained volumetric THCAS activity of PP2_HC cultures was only in the range of the latest published results (Taura et al. 2007).
Optimization of THCAS expression in P. pastoris
Procedures and medium composition were taken from Taura et al. (2007) as a starting point for expression optimization. Preliminary experiments indicated that methanol feeding at low cell densities needed optimization due to a slow metabolic rate of the yeast cultures. To ensure that methanol did not accumulate over time but was consumed before next methanol supplementation, expression cultures of PP2_HC were inoculated from overnight precultures at a higher cell density of 4.2 gCDW l−1 (OD600 of 20) in mBMMY. Methanol was added every 24 h at 0.5 % (v/v). 10, 15, 20 and 25 °C were tested for functional THCAS expression. The highest obtained activities during the cultivations are shown in Table 2. Detailed cultivation results of 10, 20 and 25 °C are given in the supplementary information.
The highest intracellular activity of THCAS (98 ± 5 pkat ml−1) was obtained at 15 °C for 192 h of cultivation (Fig. 2). THCAS activity was also found in the culture supernatant (44 ± 4 pkat ml−1) which might be due to misdirection into secretory vesicles upon overexpression (Rothman and Stevens 1986). Since microscopic analyses showed only intact cells and protein content was constant over the cultivation time in the supernatant (data not shown), cell lysis seems not responsible for supernatant activity. Considering the different assay temperatures used by Taura et al. (2007) (30 °C) and in this study (37 °C) (Fig. 3), the obtained volumetric THCAS activities were increased by 6350 % compared to the activity reported by Taura et al. (2007). Nevertheless, methanol was consumed at every feeding point at later stages of cultivation presenting a possibility for further optimization. Additionally, flocculation of cells could be observed in all PP2_HC cultivations. Together with the finding of increased functionally expressed THCAS at decreasing temperatures, it seems likely that at higher metabolic rates the correct folding of the THCAS might present a challenge for the cells, as it includes its oxidative folding in the ER together with an excessive production of reactive oxygen species (Delic et al. 2014).

Expression of THCAS using PP2_HC; Cultures were grown in 3-baffled shaking flasks at 200 rpm and 15 °C. Methanol was added every 24 h at a concentration of 0.5 % (v/v). Data points represent the means of three biological replicates with two technical replicates and error bars represent the standard deviation

Δ9-Tetrahydrocannabinolic acid synthase (THCAS) activity of cell lysate supernatant at different temperatures. Cells were lysed as described before and cell debris was centrifuged at 13,000×g at 4 °C for 5 min. 100 % activity at 52 °C confers to 623 ± 10 pkat g −1CDW . At 37 °C, 61 % maximum THCAS activity was observed (381 ± 8.3 pkat g −1CDW ). Data points represent the means of three biological replicates and error bars represent the standard deviation
Whole cell bioconversion of cannabigerolic acid (CBGA) to Δ9-tetrahydrocannabinolic acid (THCA) using P. pastoris cells
Preliminary experiments showed a low solubility of CBGA in aqueous solutions, e.g. around 200 µM in 100 mM sodium citrate buffer pH 5.5 (data not shown), impairing the production of higher amounts of THCA using cell-free aqueous systems or purified proteins. On the contrary, an immediate uptake of at least 14 mM CBGA into the cells could be observed (data not shown). Therefore, whole cell bioconversion of CBGA to THCA was investigated. A temperature dependency of THCAS activity in cell lysate supernatant is shown in Fig. 3.
Whole cell assays were performed with 10.5 gCDW l−1 at the lysate’s maximum activity (52 °C) and at a temperature where cells are still viable (37 °C) (Fig. 4a). Maximum activities of whole cells at 37 and 52 °C were 328 ± 6 and 601 ± 32 pkat g −1CDW , respectively (10.5 gCDW l−1 employed). During the whole assay time less than 1 % of the added CBGA (and THCA at later time points) is present in the supernatant. This indicates that CBGA and THCA are embedded into the cell membranes due to their hydrophobic character. The substrate concentration available for the enzyme seems non-limiting for high catalytic rates, as the maximum activities of whole cells are comparable to cell-free enzyme assays. Furthermore, since no enzyme activity could be determined in the supernatant after centrifugation, cell lysis does not occur during the timeframe of bioconversion.

Whole cell bioconversion of cannabigerolic acid (CBGA) to Δ9-tetrahydrocannabinolic acid (THCA) using PP2_HC cells at 37 °C and 52 °C (a) and cell viability during bioconversion (b). Cells were cultivated as described before, centrifuged, washed with 100 mM sodium citrate buffer pH 5.5 and finally resuspended in 400 µl 100 mM sodium citrate buffer pH 5.5–10.5 gCDW l−1 (OD600 of 50). Conversion was started by addition of 1 mM CBGA (0 h). At every subsequent time-point, cells were separated from the supernatant and resuspended in fresh 100 mM sodium citrate buffer pH 5.5 before new substrate (1 mM) was added [marked with arrows for 37 °C (black) and 52 °C (grey)]. CBGA and THCA concentrations in the supernatant as well as THCA synthase (THCAS) activity in the supernatant were measured at every time-point. CBGA and THCA concentrations in the supernatant were always below 10 µM and no THCAS activity could be measured in the supernatant (data not shown). 100 % cell viability at 37 and 52 °C confer to 340 ± 14 and 410 ± 89 colony forming units, respectively. Data points represent the means of three biological replicates and error bars represent the standard deviation
The amount of enzyme used in the bioconversion was able to convert 1 mM CBGA to THCA (0.36 g THCA l−1) before loss of activity arose. As this effect is not temperature-dependent (Fig. 4b), the inactivation of enzyme might be due to H2O2 production upon FAD regeneration. Nevertheless, the concentration of THCA was increased by 400 % compared to reactions with lysate and by 900 % compared to reports from Taura et al. (2007). Increasing the employed amount of cells could yield higher THCA levels. Furthermore, co-expression of a catalase or coupling THCA production to enzyme expression during cell growth could prolong enzymatic activity and thus increase THCA levels.
Conclusion
The expression of THCA synthase from Cannabis sativa l. was investigated using prokaryotic and eukaryotic expression systems. While no functional expression could be achieved in E. coli, the highest enzyme activity was obtained in P. pastoris cultures. Under optimized conditions, volumetric THCAS activity levels were increased by 6350 % compared to previous reports (Taura et al. 2007). The solubility issues in a biotechnological THCA production could be circumvented by employing P. pastoris whole cells. Finally, the whole cell bioconversion leads to the production of 1 mM THCA (0.36 g THCA l−1). Thus, in future whole cells might provide an alternative method for the production of pharmaceutical THC.
References
Carlini EA (2004) The good and the bad effects of (-) trans-delta-9-tetrahydrocannabinol (Delta 9-THC) on humans. Toxicon 44:461–467
Delic M, Graf A, Koellensperger G et al (2014) Overexpression of the transcription factor Yap1 modifies intracellular redox conditions and enhances recombinant protein secretion. Microb Cell 1:376–386
Ernst O, Zor T (2010) Linearization of the bradford protein assay. J Vis Exp 38:1–6
Mechoulam R (1970) Marihuana Chemistry. Science 168:1159–1165
Pertwee RG (2006) Cannabinoid pharmacology: the first 66 years. Br J Pharmacol 147:S163–S171
Rothman J, Stevens TH (1986) Protein sorting in yeast: mutants defective in vacuole biogenesis mislocalize vacuolar proteins into the late secretory pathway 47:1041–1051
Shoyama Y, Tamada T, Kurihara K et al (2012) Structure and function of ∆1-tetrahydrocannabinolic acid (THCA) synthase, the enzyme controlling the psychoactivity of Cannabis sativa. J Mol Biol 423:96–105
Sirikantaramas S, Morimoto S, Shoyama Y et al (2004) The gene controlling marijuana psychoactivity: molecular cloning and heterologous expression of Delta1-tetrahydrocannabinolic acid synthase from Cannabis sativa L. J Biol Chem 279:39767–39774
Stehle F, Stubbs MT, Strack D, Milkowski C (2008) Heterologous expression of a serine carboxypeptidase-like acyltransferase and characterization of the kinetic mechanism. FEBS J 275:775–787
Taura F, Dono E, Sirikantaramas S et al (2007) Production of Delta(1)-tetrahydrocannabinolic acid by the biosynthetic enzyme secreted from transgenic Pichia pastoris. Biochem Biophys Res Commun 361:675–680
Tolner B, Smith L, Begent RHJ, Chester KA (2006) Production of recombinant protein in Pichia pastoris by fermentation. Nat Protoc 1:1006–1021
Trost BM, Dogra K (2007) Synthesis of (-)-Delta9-trans-tetrahydrocannabinol: stereocontrol via Mo-catalyzed asymmetric allylic alkylation reaction. Org Lett 9:861–863
Acknowledgments
This study was financially supported by the Graduate Cluster Industrial Biotechnology (CLIB). The authors are thankful to the thesis students for their excellent help during the laboratory work: Dirk Münker, David Dannheisig and Madeleine Dorsch. We are also grateful to Parijat Kusari for critically reading this manuscript. Studies were conducted with the permission of No. 4584989 issued by the Federal Institute for Drugs and Medical Devices (BfArM), Germany.
Supporting information
Supplementary Table 1: List of microorganisms used for expression of THCAS.
Supplementary Table 2: List of plasmids.
Supplementary Fig. 1: Screening of P. pastoris clones—volumetric THCAS activity; cultures were inoculated at 0.105 gCDW l−1. Cultures were grown at 200 rpm and 20 °C. Methanol was added every 24 h at 0.5 % (v/v). Values are calculated from biological duplicates.
Supplementary Fig. 2: Screening of P. pastoris clones—specific THCAS activity; cultures were inoculated at 0.105 gCDW l−1. Cultures were grown at 200 rpm and 20 °C. Methanol was added every 24 h at 0.5 % (v/v). Values are calculated from biological duplicates.
Supplementary Fig. 3: Expression of THCAS using PP2_HC; Cultures were grown in 3-baffled shake-flasks at 200 rpm and 10 °C. Methanol was added every 24 h at 0.5 % (v/v). Data points represent the means of three biological replicates with two technical replicates and error bars represent the standard deviation.
Supplementary Fig. 4: Expression of THCAS using PP2_HC; Cultures were grown in 3-baffled shake-flasks at 200 rpm and 20 °C. Methanol was added every 24 h at 0.5 % (v/v). Data points represent the means of three biological replicates with two technical replicates and error bars represent the standard deviation.
Supplementary Fig. 5: Expression of THCAS using PP2_HC; Cultures were grown in 3-baffled shake-flasks at 200 rpm and 25 °C. Methanol was added every 24 h at 0.5 % (v/v). Data points represent the means of three biological replicates with two technical replicates and error bars represent the standard deviation.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Zirpel, B., Stehle, F. & Kayser, O. Production of Δ9-tetrahydrocannabinolic acid from cannabigerolic acid by whole cells of Pichia (Komagataella) pastoris expressing Δ9-tetrahydrocannabinolic acid synthase from Cannabis sativa l.. Biotechnol Lett 37, 1869–1875 (2015). https://doi.org/10.1007/s10529-015-1853-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10529-015-1853-x