
Abstract
Cry1Ie is encoded by cry1Ie, a gene that is silent in Bacillus thuringiensis strains but can be over-expressed in Escherichia coli. A protease-resistant core form of Cry1Ie with a molecular weight of approx. 55 kDa was found among the products of trypsin digestion. The oligomer and monomer of the protease-resistant core form were purified separately. The oligomer fraction contained a small quantity of dimer and a large amount of aggregates larger than tetramers. The 50% lethal concentrations (LC50) of the full-length Cry1Ie and both the monomer and oligomer fractions of the protease-resistant core form against Plutella xylostella were 14, 21 and 1400 μg/ml, respectively.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Crystal (Cry) proteins are produced by Bacillus thuringiensis (Bt) during sporulation. Many Cry proteins are synthesized as protoxins with a molecular mass of 130 kDa. Upon ingestion by an insect, the protoxin is activated in the midgut by gut proteases and is converted into a 65 kDa toxin that is derived mainly from the N-terminal region of the protoxin (Höfte and Whiteley 1989).
Oligomerization occurs in many bacterial pore-forming toxins. The association of these toxins with their respective receptors is a crucial step in the induction of proteolytic processing by membrane-associated proteases, triggering oligomerization and the subsequent insertion of the toxin into cellular membranes (Rosenberger et al. 2000). Many studies have focused on the oligomerization of Cry proteins (Puntheeranurak et al. 2005; Vie et al. 2001). Moreover, oligomers occur in solutions of Cry proteins (Feng and Becktel 1994; Walters et al. 1994; Guereca and Bravo 1999). Cry1Aa, Cry1Ac, Cry1C, Cry1D and Cry3A toxins exist in solution as a mixture of monomers and high-molecular-mass aggregates with apparent masses greater than 600 kDa, and native Cry toxins do not form oligomers of a defined size (Guereca and Bravo 1999). However, the level of insecticidal activity of oligomers in solution has not been reported.
Many cry1I-type genes are silent in Bacillus thuringiensis strains because they are often located downstream of the cry1 genes and a strong cry1 transcriptional terminator is present in the intervening sequence between the cry1 and cry1I genes (Gleave et al. 1993). Although the cry1Ie genes are silent in Bt strains, they have been over-expressed in Escherichia coli BL21(DE3), and the 84 kDa expressed product was toxic to Plutella xylostella, Ostrinia furnacalis and Leguminivora glycinivorella (Song et al. 2003).
In the present study, the protease-resistant core form of Cry1Ie protein was purified, monomers and oligomers were obtained, and the insecticidal activities of both were determined.
Materials and methods
Bacterial strains and plasmids
E. coli BL21(DE3) harboring the recombinant plasmid pET1Ie (Song et al. 2003) was obtained from the Institute of Plant Protection, Chinese Academy of Agricultural Sciences.
Expression and purification of Cry1Ie
E. coli BL21(DE3) harboring the recombinant plasmid pET1Ie was incubated at 37°C in liquid LB medium containing 100 μg ampicillin/ml. The expression of Cry1Ie in the cells was induced with 0.4 mM IPTG at 20°C for approx 15 h. The cells were collected by centrifugation at 5,000×g for 10 min and resuspended in lysis buffer (20 mM Tris/HCl, pH 8.0). The cells were lysed by ultrasonication at 200 W with a interval of 15 s on ice, and the inclusion bodies were separated from the soluble supernatant by centrifugation at 15,000×g for 10 min at 4°C. The inclusion bodies were washed four times with lysis buffer and three times with distilled water to remove the remaining soluble proteins and then solubilized in 0.5 mM Na2CO3 (pH 10.2). The insoluble components were separated by centrifugation at 15,000×g for 10 min and discarded. The supernatant was loaded onto a Q-Sepharose Fast Flow column pre-equilibrated with buffer containing 0.5 mM Na2CO3 at pH 10.2. The column was then washed with 2–3 column volumes of the Na2CO3 buffer and eluted by with a linear gradient of 0–1 M NaCl while the effluent was monitored at 260 and 280 nm. The peak fractions were collected and analyzed by SDS-PAGE.
Trypsin digestion and purification of the protease-resistant core form of Cry1Ie
The Cry1Ie protoxin was digested by treatment with trypsin (TPCK-treated, 1:20 w/w) in 50 mM Na2CO3 at 37°C for 1 h. The trypsin-digested products were loaded onto a Superdex-200 column HR (10/30) using a FPLC apparatus. The column was pre-equilibrated with 50 mM Na2CO3 at pH 10.2 and eluted at 0.6 ml/min; the eluate was monitored at 260 and 280 nm, and the peak fractions were collected.
N-terminal amino acid residue sequence determination
The purified protease-resistant core form of the protein was identified by SDS-PAGE, and it was subsequently transferred onto polyvinylidene difluoride membrane filters. The membrane-bound proteins, visualized by Coomassie Blue staining, were excised, and the N-terminal amino acid residue sequence was determined using the Procise 491 sequencing system from Applied Biosystems.
Glutaraldehyde crosslinking
Glutaraldehyde (25% w/v) was added to the protein solution to give 2% (v/v). After 2 min, the crosslinking reaction was quenched by the addition of solid NaBH4 to give a final NaBH4/glutaraldehyde molar ratio of 10:1. Then, 8 μl 25% deoxycholic acid solution and 25 μl 78% TCA solution were added (Lai et al. 1997). The samples were then centrifuged at 12,000×g for 10 min, and the precipitates were washed several times with distilled water and then redissolved in 30 μl of 1.5 M Tris/HCl buffer (pH 8.8) containing 1% (w/v) SDS and 50 mM DTT. Finally, the samples were boiled for 3 min and SDS-PAGE was performed at room temperature using the Tris/glycine buffer system described by Laemmli.
Protein quantification
The protein concentrations were determined by the method of Bradford using BSA as a standard.
Bioassay
Toxicity assays on the second instar larvae of Plutella xylostella were conducted on fresh leaf disks using the leaf-dip bioassay (Tabashnik et al. 1993). Disks cut from the leaves of cabbages grown in a greenhouse were used to grow P. xylostella. Different concentrations of toxin were brushed on leaves using a clean, autoclaved brush, and then the leaves were dried naturally at room temperature. 50 mM Na2CO3 (pH 10.2) was used as negative control. Twenty larvae were placed on a leaf disk and analyzed after 48 h. The 50% lethal concentrations (LC50) were calculated by probit analysis (Finney 1971) after repeating the bioassay at least four times.
Results
Expression and purification of Cry1Ie
Most of the Cry1Ie protein expressed in E. coli occurred as an insoluble form in inclusion bodies. The Cry1Ie protein, like many other Cry proteins, can be solubilized in 0.5 mM Na2CO3 at pH 10.2. The Cry1Ie protein was further purified by ion-change chromatography and analyzed by SDS-PAGE (Fig. 1, Lane 2).

SDS-PAGE analysis of purified full-length (protoxin) Cry1Ie protein. Lane 1, molecular mass marker; Lane 2, purified Cry1Ie protein
Trypsin digestion and purification of the protease-resistant core form of Cry1Ie
Cry1Ie was activated by treatment with trypsin from bovine pancreas. During activation, a 66 kDa protein was found which has been previously detected in the activation of many other Cry proteins. Moreover, among the digestion products, a protease-resistant core form with a molecular weight of approximately 55 kDa was observed (Fig. 2, Lane 3). The core form gradually became the main product as the concentration of trypsin was increased. The protease-resistant core form was purified using a Superdex-200 column, and its elution profile was recorded (Fig. 3a). Although peak 1 and peak 2 had different elution volumes, the proteins from both peaks had the same mobility in SDS-PAGE gels, implying that they had the same molecular mass (Fig. 3b).

Digestion of the full-length (protoxin) Cry1Ie with varying concentrations of trypsin at 37°C for 1 h. Lane 1, molecular mass marker; Lane 2, isolated Cry1Ie; Lanes 3–8, products of Cry1Ie digestion with trypsin at protein (weight/weight) ratios of 1:20, 1:30, 1:50, 1:80, 1:100 and 1:150

Purification of the protease-resistant core form of Cry1Ie protein. a Elution profile by size-exclusion chromatography of trypsin (1:20, w/w)-digested products. The solid line represents the absorbance at 280 nm, while the dashed line represents the absorbance at 260 nm. The void volume of the column was 8.8 ml as determined using Blue Dextran 2000. The flow rate was 0.6 ml/min. b Electrophoretic analysis of the peak fractions. Lane 1, molecular mass marker; Lanes 2–4, middle fractions of peak 1; Lanes 5–6, middle fractions of peak 2
This result indicated that both a monomer and an oligomer existed in the activated 55-kDa product. Glutaraldehyde was used as a crosslinking reagent to evaluate the oligomeric state of the protease-resistant core form of Cry1Ie (Fig. 4). A product with a molecular mass of 110 kDa was detected on SDS-PAGE after the crosslinking reaction, and a larger amount of protein was observed in a considerably higher oligomeric form, even larger than a tetramer. This result indicated that the oligomer of the core form included a small amount of dimer but was mainly composed of higher order aggregates larger than tetramers. The second peak consisted of the monomer of the core form of the Cry1Ie protein. Comparing the quantities of the collected fractions of the two peaks, we determined that about 40% of the trypsin-digested product existed in the oligomeric form.

SDS-PAGE analysis of glutaraldehyde crosslinked proteins of peaks 1 and 2. Lane 1, molecular mass marker; Lane 2, glutaraldehyde-crosslinked protein of peak 1; Lane 3, glutaraldehyde-crosslinked protein of peak 2; Lane 4, protein from peak 1; Lane 5, protein from peak 2; Lane 6, molecular mass marker
In order to determine whether these two forms of the Cry1Ie protease-resistant core are interconvertible, size-exclusion chromatography was used to detect the oligomeric states after incubation at 4°C for different periods of time (Fig. 5). The result shows that the oligomer did not convert into the monomer; however, while the monomer initially remained monomeric, it then partially converted into an oligomer with time. This implies that the oligomer is more stable than the monomer.

Elution profiles by size-exclusion chromatography of the oligomer and monomer of the protease-resistant core form of Cry1Ie incubated at 4°C for different periods of time. A Elution profiles of the oligomer. Curve a represents the newly collected oligomer, while Curve b represents the oligomer incubated at 4°C for 3 days. B Elution profiles of the monomer. Curve a represents the newly collected monomer, while Curve b represents the monomer incubated at 4°C for 3 days
The N-terminal amino acid residue sequence of the purified protease-resistant core form of the protein was determined to be ATSVV, which begins with amino acid residue 154 of full-length Cry1Ie.
Bioassay of the monomer and oligomer of the protease-resistant core form of Cry1Ie
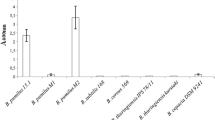
The LC50 of the full-length Cry1Ie protein and both the monomeric and oligomeric forms of the protease-resistant core of Cry1Ie were determined against P. xylostella (see Table 1). The LC50 of the monomer was slightly higher than that of full-length Cry1Ie; however, the 95% fiducial limits partially overlap. Since the molecular masses of the full-length Cry1Ie protein and the protease-resistant core form expressed in E. coli were 84 kDa (Song et al. 2003) and 55 kDa, respectively, the insecticidal activity of the core form of the Cry1Ie protein was about 35% lower than that of the full-length Cry1Ie protein. The LC50 of the oligomer was ~1,400 μg/ml, which is 70 times higher than that of the monomeric form (21 μg/ml), indicating that the insecticidal activity of the oligomer is dramatically lower.
Discussion
Like many other Cry proteins, Cry1Ie yields a 66-kDa protein during activation by trypsin. A protease-resistant core form also exists among the trypsin digestion products, with a molecular weight of approximately 55 kDa. In the core form, the N-terminus begins at amino acid 154 of full-length Cry1Ie; this is quite similar to the structure of the core form of Cry1Ia, in which the N-terminus starts with amino acid 156 (Sekar et al. 1997). The insecticidal toxicity of the core form of the Cry1Ie protein did not substantially differ from that of full-length Cry1Ie even though it lacks 153 amino acids of the N-terminus and some of the C-terminus. This result differed from the analysis of Cry1Ia, in which the core form of Cry1Ia exhibited only ~22% of the toxicity of the full-length protein (Sekar et al. 1997). This difference may in some way be related to the different target larvae used in the bioassay. We used P. xylostella larvae while Sekar et al. (1997) used Ostrinia nubilalis (European corn borer) larvae.
The crystal structure of the Cry1Aa toxin has been determined (Grochulski et al. 1995). Prediction of the secondary structure of Cry1Ie and its comparison with that of Cry1Aa indicated that the N-terminus of the Cry1Ie core is located at the beginning of helix α-4. The α-3 helix in domain I of Cry1Ab was found to contain sequences that could potentially form a coiled-coil structure important for oligomerization (Burkhard et al. 2001; Jimenez-Juarez et al. 2007). Site-directed mutagenesis in helix α-3 resulted in the formation of Cry1Ab mutants that still bound to the Bt-R1 receptor, similarly to the wild-type toxin; however, oligomerization of the mutants was affected, resulting in the complete loss of pore-forming activity and toxicity against Manduca sexta larvae (Jimenez-Juarez et al. 2007). Sequence alignment of Cry1Ab and Cry1Aa indicated that helix α-3 spans amino acids 90–119. Comparison of Cry1Ie with this Cry1Aa region indicated that helix α-3 spans amino acids 118–147 in Cry1Ie, which is outside the sequence of the protease-resistant core form of the Cry1Ie protein. Without helices α1–α3, the protease-resistant core form of Cry1Ie still has a considerable amount of the insecticidal activity like the full-length protein, suggesting that α1–α3 are not absolutely necessary for the process of pore-formation or transmembrane insertion into the midgut of the insect. In this regard, Cry1Ie appears quite different from other Cry proteins.
The insecticidal activity of the oligomer of the core form of Cry1Ie was determined and found to be substantially lower than that of the monomer. We found that the oligomer was formed in solution in absence of receptor. Conversion of oligomer into monomer did not occur to any measurable extent in vitro. The oligomers that form spontaneously in solution most likely differ from the oligomers that form on the membrane of the insect midgut. Aggregation in solution may partially mask the region of the protein that recognizes the membrane receptors. Consequently, the recognition and binding of the protein to its membrane receptors is hindered, and it is difficult for the oligomer that has already formed in solution to bind to the membrane. This may be the reason for the decrease in insecticidal activity of the oligomeric form.
References
Burkhard P, Stetefeld J, Strelkov SV (2001) Coiled coils: a highly versatile protein folding motif. Trends Cell Biol 11:82–88
Feng Q, Becktel WJ (1994) pH-induced conformational transitions of Cry IA(a), Cry IA(c), and Cry IIIA delta-endotoxins in Bacillus thuringiensis. Biochemistry 33:8521–8526
Finney DJ (1971) Probit analysis. Cambridge University Press, Cambridge, United Kingdom
Gleave AP, Williams R, Hedges RJ (1993) Screening by polymerase chain reaction of Bacillus thuringiensis serotypes for the presence of cryV-like insecticidal protein genes and characterization of a cryV gene cloned from B. thuringiensis subsp. kurstaki. Appl Environ Microbiol 59:1683–1687
Grochulski P, Masson L, Borisova S et al (1995) Bacillus thuringiensis Cry1Aa insecticidal toxin: crystal structure and channel formation. J Mol Biol 254:447–464
Guereca L, Bravo A (1999) The oligomeric state of Bacillus thuringiensis Cry toxins in solution. Biochim Biophys Acta 1429:342–350
Höfte H, Whiteley HR (1989) Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol Rev 53:242–255
Jimenez-Juarez N, Munoz-Garay C, Gomez I et al (2007) Bacillus thuringiensis Cry1Ab mutants affecting oligomer formation are non toxic to Manduca sexta larvae. J Biol Chem 282:21222–21229
Lai Z, McCulloch J, Lashuel HA et al (1997) Guanidine hydrochloride-induced denaturation and refolding of transthyretin exhibits a marked hysteresis: equilibria with high kinetic barriers. Biochemistry 36:10230–10239
Puntheeranurak T, Stroh C, Zhu R et al (2005) Structure and distribution of the Bacillus thuringiensis Cry4Ba toxin in lipid membranes. Ultramicroscopy 105:115–124
Rosenberger CM, Brumell JH, Finlay BB (2000) Microbial pathogenesis: lipid rafts as pathogen portals. Curr Biol 10:823–825
Sekar V, Held B, Tippett J et al (1997) Biochemical and molecular characterization of the insecticidal fragment of CryV. Appl Environ Microbiol 63:2798–2801
Song FP, Zhang J, Gu AX et al (2003) Identification of cry1I-type genes from Bacillus thuringiensis strains and characterization of a novel cry1I-type gene. Appl Environ Microbiol 69:5207–5211
Tabashnik B, Finson N, Chilcutt CF et al (1993) Increasing efficiency of bioassays: evaluation of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae). J Econ Entomol 86:635–644
Vie V, Van Mau N, Pomarede P et al (2001) Lipid-induced pore formation of the Bacillus thuringiensis Cry1Aa insecticidal toxin. J Membr Biol 180:195–203
Walters FS, Kulesza CA, Phillips AT et al (1994) A stable oligomer of Bacillus thuringiensis delta-endotoxin, Cry IIIA. Insect Biochem Mol Biol 24:963–968
Acknowledgments
This work was supported by grants from the Major State Basic Research Development Program of China (973 Program; No. 2009CB118902) and the National High Technology Research and Development Program of China (863 Program; No. 2006AA10A212 and No. 2006AA02Z189).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Guo, S., Zhang, Y., Song, F. et al. Protease-resistant core form of Bacillus thuringiensis Cry1Ie: monomeric and oligomeric forms in solution. Biotechnol Lett 31, 1769–1774 (2009). https://doi.org/10.1007/s10529-009-0078-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10529-009-0078-2