
Abstract
Plant growth-promoting endophytic fungi with gibberellin-producing ability were isolated from the roots of Carex kobomugi Ohwi, a common sand-dune plant, and bioassayed for plant growth-promotion. A new strain, Arthrinium phaeospermum KACC43901, promoted growth of waito-c rice and Atriplex gemelinii. Analysis of its culture filtrate showed the presence of bioactive GA1 (0.5 ng/ml), GA3 (8.8 ng/ml), GA4 (4.7 ng/ml) and GA7 (2.2 ng/ml) along with physiologically inactive GA5 (0.4 ng/ml), GA9 (0.6 ng/ml), GA12 (0.4 ng/ml), GA15 (0.4 ng/ml), GA19 (0.9 ng/ml) and GA24 (1.8 ng/ml). The fungal isolate was identified through sequence homology and phylogenetic analysis of 18S rDNA (internal transcribed region).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The internal tissues of apparently healthy roots host a variety of fungi (Vandenkoornhuyse et al. 2002). Fungal endophytes of these roots are a taxonomically diverse assemblage of ascomycetes inhabiting herbaceous and woody plants without forming mycorrhizal relationship or causing obvious disease symptoms in their hosts (Addy et al. 2005). These fungi can act as defenders against predators (Siegel and Bush 1997), growth-promoters (Bacon and White 2000) and competitors of microbial pathogens (Scannerini et al. 2001). Plant growth-promotion may be attributed to the secretion of gibberellins (GA) by the endophytic fungi in the rhizosphere (Choi et al. 2005). GA production had been reported in 12 different fungi (MacMillan 2002; Kawaide 2006). So far, there are no reports on GA production by any Arthrinium spp. Members of genus Arthrinium exist in diverse environmental conditions and can grow on a variety of substrates. Arthrinium phaeospermum forms various associations with healthy leaves, stems and roots of Arundo mauritanica, Bambusa spp., Brassica campestris, Carex spp., and Pinus officinalis. Although not very common, this fungus has world wide distribution (Agut and Calvo 2004).
Coastal regions of the world offer high economic returns and recreational opportunities and are subjected to more anthropogenic activities. The sand dunes in coastal areas are on verge of destruction due to excessive loss of native species as the efficiency of conservation and re-vegetation has been slowed by intensive human activities (Girard et al. 2002; Kim 2005). The sand-dune flora is subjected to abiotic stresss such as nutrient deficiency and high salinity. Under such adverse conditions, the role of symbiotic fungi in native plants growth and conservation cannot be overlooked. In the current study, the roots of sand-dune plants were screened for the presence of plant growth-promoting endophytic fungi. We were interested to study the possible role of these fungal symbionts in the survival of their host plants under harsh environmental conditions.
Materials and methods
Host plant, fungal strains, culture medium and growth conditions
Carex kobomugi Ohwi (Japanese sedge) was collected from the eastern coast of Korea. The screening and isolation of root endophytic fungi was carried out on Hagem minimal medium plates supplemented with 80 ppm streptomycin (Yamada et al.2001). Surface-sterilised roots were cut (0.5 cm) and placed aseptically on plates containing Hagem minimal media and incubated (25°C) until the emergence of fungi (Vazquez et al.2000). Purified fungi were grown in Czapek broth medium containing 1% glucose and peptone for GA production (Hasan 2002) at 30°C and 120 rpm for 7 days. Wild type Gibberella fujikuroi was used as control.
Screening and bioassay of fungal culture filtrates for plant growth-promoting activity
Surface-sterilized and uniconazol-treated (20 ppm) waito-c seeds were germinated on water/agar media (0.8% w/v), aseptically. Waito-c is a dwarf and GA-deficient rice cultivar. Ten μl of fungal culture filtrate suspension (30 ml of lyophilised culture filtrate suspended in 1 ml of autoclaved distilled water) were applied on apical meristems of two-leaved rice seedlings, and incubated for 7 days in a plant growth chamber. Shoot and whole seedling lengths were noted and compared with controls (G. fujikuroi culture filtrate and distilled water treated). The culture filtrate of the fungal isolate giving the best shoot elongation in the rice seedlings was bioassayed on two-leaved seedlings of a sand-dune plant, Atriplex gemelinii, as mentioned above.
Gibberellins extraction and quantification
Gibberellins were extracted from 30 ml culture filtrate of the fungal isolate CK-2-2 (Lee et al. 1998) and quantified by GC/MS in selected ion monitoring (SIM) mode. Retention time was determined by using the hydrocarbon standards to calculate the Kovats retention indices (KRI) value (Gaskin and MacMillan 1991), while the GAs quantification was based on peak area ratios of non-deuterated (extracted) GAs to deuterated (added internal standards) GAs.
Genomic DNA isolation and sequencing
Genomic DNA was isolated (see supplementary information) and its purity and quality checked by agarose gel electrophoresis. For identification, universal primers for 18S rDNA internal transcribed region (ITS) 1 and 4 were used, and the resultant PCR product was cloned and sequenced.
Phylogenetic analysis
The sequenced ITS region of the fungal isolate CK-2-2 was searched for homologous sequences using BLAST. All sequences were aligned using ClustalW. Neighbour joining tree with bootstrap consensus (500 replications) for the statistical support of tree nodes was constructed using MEGA 4 (Tamura et al. 2007).
Results and discussion
Screening and bioassay of isolated fungal culture filtrates for plant growth promoting activity
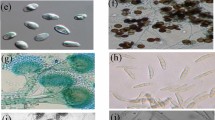
Two-leaved waito-c rice seedlings were used for preliminary screening experiment of fungal secondary metabolites. Since uniconazol blocks gibberellin synthesis during seed germination and the rice cultivar itself is a low GA producer, shoot elongation of seedlings is associated with fungal metabolite activity. Nine endophytic fungi were isolated from the roots of Carex kobomugi Ohwi. Five fungal isolates promoted shoot elongation compared to controls after 7 days (Fig. 1). Of these, the fungal isolate CK-2-2 gave maximum lengths and was thus selected for further study.

Effect of fungal culture filtrates (10 μl) on the length of waito-c rice seedlings after 7 days of incubation. Each sample was assayed in triplicate. Results are expressed as means (bars) with SD (error bar). Whole plant lengths as well as shoot lengths were measured after 7 days of treatment. The culture filtrate treated seedlings showed increased shoot and whole plant lengths. The highest length (shoot/plant) was induced by CK-2-2 (8.5 cm/20.4 cm) which was approximately similar to length (shoot/plant) shown by G. fujikuroi (8.7 cm/19.1 cm). The non-culture filtrate treated seedlings showed much lower length as seen in the figure
Plant growth-promoting activity of culture filtrate (CK-2-2) was verified by conducting a bioassay on two-leaved seedlings of the sand dune plant Atriplex gemelinii. The average shoot length affected by the culture filtrates of the fungal isolate CK-2-2 was 3.2 cm, while that by the culture filtrates of G. fujikuroi was 3.1 cm. Both treatments gave increased lengths as compared to non-culture filtrate treated seedlings (2 cm) after 15 days. Since the fungal isolate CK-2-2 was isolated from a sand-dune plant, the effect of its culture filtrate metabolites was analyzed on Atriplex gemelinii, a native dune plant. This plant was chosen for bioassay due to the absence of seed dormancy and rapid germination rate of seeds (Kunkel 1984). The growth rate of A. gemelinii seedling was much slower than that of the rice seedlings, as A. gemelinii is a slow growing plant (0.5 m annually). Current results confirm previous reports of shoot length promotion through fungal culture filtrate treatment (Choi et al. 2005). For both bioassay experiments nutrient free water–agar media were used which is needed for the determination of the sole effect of culture filtrate on seedlings growth.
Gibberellins quantification
Physiologically active GAs along with GA5, GA9, GA12, GA15, GA19 and GA24 were detected by GC-MS SIM. Quantities of GA3, GA4 and GA7 were more than those produced by G. fujikuroi under the same growth conditions (Fig. 2). GC-MS SIM spectra for bioactive GAs are given in supplementary Figs. 1, 2, 3 and 4.

Concentrations of the CK-2-2 fungal isolate GAs against G. fujikuroi. Of the 4 bioactive GAs, only GA1 was produced in lower concentration (0.5 ng/ml) by CK-2-2 than by G. fujikuroi (1.3 ng/ml), while the non-bioactive GAs were produced in approximately equal concentrations except GA24 (1.8 ng/ml), which was almost twice the concentration of GA24 by G. fujikuroi (0.7 ng/ml)
Detection of higher quantities of GA3, GA4 and GA7 in culture filtrates of fungal isolate CK-2-2 as compared to G. fujikuroi exerted no better elongation effect on shoot and whole length of seedlings. This might be due to the presence of some growth suppressors in the culture filtrate of CK-2-2.
Identification and phylogenetic analysis
The analysis of sequence homology report through BLAST and neighbour joining tree construction from aligned data identified the fungal isolate CK-2-2 as a new strain of A. phaeospermum (99% sequence homology of ITS regions and 100% bootstrap support) (Fig. 3). The ITS sequence of this strain has been submitted in GenBank database under accession number EU821332. The fungal strain was deposited in Korean Agricultural Culture Collection, and was named as A. phaeospermum KACC 43901. For fungal identification, ITS region sequencing has gained importance in a short time, because of the presence of highly conserved 5.8S gene that helps in the identification of higher taxonomic level, as well as the presence of highly variable flanking genes (ITS1 and 4) useful for the identification at a lower taxonomic level (Sugita and Nishikawa 2003). Neighbour-joining construction is a rapid method of relating query sequence with homologous sequences while bootstrapping provides the necessary statistical analysis to overcome possibilities of errors in identification (Yang and Khuri 2003).

Identification of fungal isolate CK-2-2 by phylogenetic analysis. Neighbour joining tree (1000 bootstrap replications) was constructed using 17 taxa (16 refrence sequences and 1 clone). Bootstrap support of 100 for subclade of CK-2-2 with A. phaeospermum (99% sequence homology) strongly suggest this fungal isolate as a new strain of A. phaeospermum (A. phaeospermum KACC43901). The tree was drawn to scale (0.02) that represents evolutionary distances in units of base substitutions per site, computed by maximum composite likelihood method. Bootstrap values less than 50 are not shown. Gaps are treated as missing data and eliminated from dataset during tree construction
The current research project was carried out to isolate competent fungi from sand dunes which could be used for dune re-vegetation. We conducted experiments with waito-c rice and Atriplex gemelinii seedlings in plant growth chambers and obtained growth promotion influenced by the presence of gibberellins in the fungal culture filtrate. The genus Arthrinium has never been reported for the production of gibberellins before and the current study reports about GA production in Arthrinium phaeospermum for the first time. Further study on the characterisation of gene cluster coding for gibberellins and the development of optimised gibberellins producing media for A. phaeospermum KACC43901 is needed.
References
Addy HD, Piercey MM, Currah RS (2005) Microfungal endophytes in roots. Can J Bot 83:1–13
Agut M, Calvo MA (2004) In vitro conidial germination in Arthrinium aureum and Arthrinium phaeospermum. Mycopathol 157:363–367
Bacon CW, White JF (2000) Microbial endophytes. Marcel Dekker, New York
Choi WY, Rim SO, Lee JH et al (2005) Isolation of gibberellins producing fungi from the root of several Sesamum indicum plants. J Microbiol Biotechnol 15(1):22–28
Franck C, Lammertyn J, Nicolaï B (2005) Metabolic profiling using GC-MS to study biochemical changes during long-term storage of pears. In: Mencarelli F, Tonutti P (eds) Proceedings of 5th international postharvest symposium. Acta Hort 682:1991–1998
Gaskin P, MacMillan J (1991) GC-MS of gibberellins and related compounds. Methodology and a library of reference spectra. Cantocks Enterprises, Bristol
Girard M, Lavoie C, Theriault M (2002) The regeneration of a highly disturbed ecosystem: a mined peat land in southern Quebec. Ecosystems 5:274–288
Hasan HAH (2002) Gibberellin and auxin producing plant root fungi and their biosynthesis under salinity-calcium interaction. Rostlinná Výroba 48(3):101–106
Higgs RE, James AZ, Jeffrey DG et al (2000) Rapid method to estimate the presence of secondary metabolites in microbial extracts. Appl. Environ. Microbiol 67(1):371–376
Kawaide H (2006) Biochemical and molecular analysis of gibberellin biosynthesis in fungi. Biosci Biotechnol Biochem 70:583–590
Kim KD (2005) Invasive plants on disturbed Korean sand dunes. Estuar Coast Shelf Sci 62:353–364
Kunkel G (1984) Plants for human consumption. Koeltz Scientific Books. ISBN 3874292169
Lee IJ, Foster K, Morgan PW (1998) Photoperiod control of gibberellin levels and flowering in sorghum. Plant Physiol 116:1003–1011
MacMillan J (2002) Occurrence of gibberellins in vascular plants, fungi and bacteria. J Plant Growth Regul 20:387–442
Morakotkarn D, Kawasaki H, Seki T (2006) Molecular diversity of bamboo-associated fungi isolated from Japan. FEMS Microbiol Lett 266:10–19
Scannerini S, Fusconi A, Mucciarelli M (2001) The effect of endophytic fungi on host plant morphogenesis. In: Seckbach J (ed) Cellular origin and life in extreme habitats. Symbiosis. Kluwer Academic Publishers, Dordrecht, pp 427–447
Siegel MR, Bush LP (1997) Toxin production in grass/endophyte associations. In: Carroll GC, Tudzynski P (eds) The mycota. Springer-Verlag, Heidelberg, pp 185–207
Sugita T, Nishikawa A (2003) Fungal identification method based on DNA sequence analysis. Reassessment of the methods of the pharmaceutical society of Japan and the Japanese pharmacopoeia. J Health Sci 49(6):531–533
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Vandenkoornhuyse P, Baldauf SL, Leyval C et al (2002) Extensive fungal diversity in plant roots. Science 295:2051
Vazquez MM, Cesar S, Azcon R et al (2000) Interaction between arbuscular mycorrhizal fungi and other microbial inoculants (Azospirillum, Pseudomonas, Trichoderma) and their effects on microbial population and enzyme activities in the rhizosphere of maize plants. Appl Soil Ecol 15:261–272
Yamada A, Ogura T, Degawa Y et al (2001) Isolation of Tricholoma matsutake and T. bakamatsutake cultures from field-collected ectomycorrhizas. Mycoscience 42:43–50
Yang C, Khuri S (2003) PTC: an interactive tool for phylogenetic tree construction. In: Proceedings of IEEE Computer Society Bioinformatics Conference. Stanford University, p 476
Acknowledgment
This research is a part of ‘Eco-technopia 21 project’ supported by the Korea Ministry of Environment.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Khan, S.A., Hamayun, M., Kim, Hy. et al. A new strain of Arthrinium phaeospermum isolated from Carex kobomugi Ohwi is capable of gibberellin production. Biotechnol Lett 31, 283–287 (2009). https://doi.org/10.1007/s10529-008-9862-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10529-008-9862-7