
The major histocompatibility complex (MHC), with its extraordinary levels of genetic variation, is thought to be an essential aspect of the ability of an organism to recognize different parasites and pathogens. It has also been proposed to regulate reproductive processes in many aspects. Here we examine the genetic variation of the second exon of the MHC class II B genes of the crested ibis, an endangered species known to descend from just two breeding pairs rediscovered in 1981. Only five alleles are identified by single-strand conformation polymorphism (SSCP) analysis of 36 samples taken from both wild and captive populations, and a comparatively low level of divergence between MHC alleles is observed. We suggest that representative sampling of individuals with most of the different MHC allele genotypes to constitute a founder population, together with the monitoring of the pathogen status of candidate sites before release, is of great importance for raising the success rate of reintroduction for the crested ibis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
The importance of genetic diversity over the long term (i.e., the maintenance of adaptive evolutionary potential) as well as the short term (i.e., the maintenance of reproductive fitness) makes it a primary focus for conservation genetics (Frankham et al., 2002). Maintenance of genetic diversity is particularly of major concern for bottlenecked species, since population bottlenecks may reduce the amount of genetic variation and consequently result in lowered fitness, reduced potential for future adaptation, as well as elevated extinction risk. Furthermore, when reintroduction is being scheduled upon the increase of individual number in the post-bottleneck populations, it is fundamental to evaluate the genetic variation in the source populations to provide an indicator of the long-term fitness of the reintroduced populations.
The genes in the major histocompatibility complex (MHC) are thought to be of important adaptive significance, particularly for resistance to parasites and pathogens (Hedrick and Kim, 2000), and perhaps for mate choice (Penn and Potts, 1998) and maternal–fetal interaction (Lokki and Laitinen, 2001). The extraordinary levels of genetic variation in many MHC genes, the highest known in any vertebrate genes, are thought to be an essential aspect of the ability of an organism to recognize different parasites and pathogens (Hedrick and Kim, 2000). It has been suggested that populations with reduced MHC variability would be particularly vulnerable to infectious disease (Edwards and Potts, 1996). Therefore, investigating the extent of MHC variation for bottlenecked species should provide insight into their potential resistance or susceptibility to various diseases, which would be of great importance for the reestablishment of self-sustaining populations after their population bottlenecks.
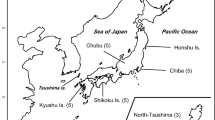
Few species have experienced a more severe population bottleneck than the crested ibis Nipponia nippon (order Ciconiiformes, family Threskiornithidae). It is regarded as “Endangered” in the 2004 IUCN Red List of Threatened Species (http://www.redlist.org) and is listed as an endangered species on the State Key Protected Wildlife List of P.R. China (Wang, 1998). Historically, the species was once a common sight throughout the Russian Far East, Korea, and Japan, through to central China (Fig. 1; Yamashina, 1975). Habitat loss, hunting, and environmental pollution decimated this species so that by the late 1970s, wild populations were extinct in Korea and the Russian Far East (Hankcock et al., 1992). The death of the very last indigenous crested ibis in Sado, Japan, on October 10, 2003 finally marked the extinction of the relict Japanese crested ibis population (http://www.redlist.org).

Distribution of crested ibis Nipponia nippon (Bird Life International, 2001). The only known wild population currently exists in Yangxian County and surrounding areas.
In China, the rediscovery of seven wild crested ibis in Yangxian, Shaanxi Province, in 1981, including two breeding pairs and their three nestlings, renewed hope for the species (Liu, 1981). This magnificent bird has since been the focus of intensive conservation efforts, and the Shaanxi Crested Ibis Nature Reserve was therefore established in 1990, in which the only wild population in the world has been gradually recovering. Meanwhile, special attention has been paid to captive breeding of this rare bird. The captive rearing and breeding program began at the Shaanxi Crested Ibis Breeding Centre in 1990 with the rescue of sick or injured chicks and the collection of some of the eggs deserted by parental birds in the wild (Xi et al., 2001, 2002). By the year 2002 the total number of crested ibis, including the wild and captive populations in China, had recovered to nearly 300 individuals (Ding and Liu, 2002). The population has continued to increase since, and the first attempt at reintroduction was made in 2004 (Xi, personal communication).
Despite the endangered status of crested ibis, surprisingly little has been published on its genetic diversity, with the exception of the previous study on its noncoding mitochondrial DNA control region, which did indeed reveal an extremely low level of genetic diversity for the species (Zhang et al., 2004). Therefore, this contribution presents the very first study to investigate the genetic variation in its functional nuclear loci from both wild and captive populations by examining the highly variable second exon of the MHC class II B genes. Our aims were i) to reveal levels of polymorphism in the MHC class II B genes of this highly endangered species and ii) to provide feasible strategies for its reintroduction in the near future.
MATERIALS AND METHODS
Sample Source
In total, 36 samples were randomly obtained during 1990 to 2000. Ten samples were collected from wild birds at the Shaanxi Crested Ibis Nature Reserve, and 26 samples were collected from captive birds in the Shaanxi Crested Ibis Breeding Centre, Yangxian, Shaanxi (Table I).
Laboratory Procedures
Total genomic DNA was isolated and purified by proteinase K digestion and phenol/chloroform extraction following standard procedures (Sambrook et al., 1989). First a pair of highly degenerate primers (305 and 306, developed by Edwards et al., 1995) was used to amplify a portion of the second exon of the MHC class II B genes in nine randomly selected individuals (25% of the total population studied). PCR fragments of interest were eluted by a UNIQ-5 Column DNA Gel extraction kit (Sangon, China), ligated into pMD 18-T vectors (TaKaRa, China), and sequenced following the extension dideoxy-chain termination method with universal primers (M13+/M13−) and the BigDye terminator cycle sequencing kit (Perkin-Elmer) according to the manufacturer’s instructions, then analyzed on an ABI PRISM 377-96 DNA sequencer. The sequences obtained included both MHC class II B exon 2 sequences and some non-MHC sequences. Based on 11 MHC sequences, we designed a pair of primers for efficient amplification of a 205 bp portion of the crested ibis MHC class II B exon 2: CIMF (5′-TTATTTTAATGGGACCGAGCGGG-3′) and CIMR (5′-CCGTAATTGTGGCGGCAGTAC-3′). Polymerase chain reaction (PCR) amplification was carried out on a PTC-200 Peltier Thermal Cycler. A 50 μL reaction was performed using 50 ng of template DNA, 5 μL of 10×reaction buffer, 1.5 mM MgCl2, 150 mM of each dNTP, 0.25 mM of each primer, and 1.5 units of TaKaRa ExTaq DNA Polymerase (TaKaRa, China). The thermal cycling profile was as follows: an initial hot-start at 95°C for 4 min; 30 amplification cycles of denaturation at 94°C for 45 s, annealing at 57°C for 45 s, and extension at 72°C for 45 s; followed by a final incubation at 72°C for 10 min to ensure complete extension of the PCR products. The PCR amplification products were separated and eluted by agarose gel electrophoresis and UNIQ-5 Column DNA Gel extraction kit (Sangon, China).
Single-strand conformation polymorphism (SSCP) analysis, which resolves sequences with the same length based on sequence-dependent conformation (Sunnucks et al., 2000), was used to allow a highly detailed analysis of the MHC sequences. To perform nondenaturing SSCP electrophoresis, a mixture of 3 μL of the purified PCR products and 3 μL of denaturing loading dye (95% formamide, 10 mm NaOH, 0.25% bromophenol blue, 0.25% xylene cyanol) was heated at 95°C for 10 min and cooled in ice water prior to being loaded onto SSCP gels. The gels contained 8% 37.5:1 acrylamide/bisacrylamide with 2.5% crosslinking, and sequences were separated in 0.5×TBE running buffer at 20°C by 150 V for 14 h on the Dcode System (Bio-Rad). All individuals were processed twice using the SSCP procedure to avoid categorizing PCR artifacts as new alleles (such as amplified Taq errors or recombinant sequences generated by template jumping by Taq). After separation, the gels were fixed in 10% acetic acid for 30 min, washed with dH2O, and silver-stained following Bassam et al. (1991). Bands of interest were then eluted by the Polyacrylamide Gel DNA Purification Kit (Tianwei-Biotech, China). Genetic differences between alleles were verified by sequencing the reamplified products, and sequences were identical in all replicates and from both strands.
Statistical Treatment
Sequences were aligned using the Clustal X program (Thompson et al., 1997). Initial sequence comparisons and measures of variation were performed using MEGA version 2.1 (Kumar et al., 2001), and MEGA was also used to calculate the relative rate of nonsynonymous (d N ) and synonymous (d S ) substitutions according to modified Nei and Gojobori (1986) and applying the correction of Jukes and Cantor (1969) for multiple hits.

Amino acid sequences for the crested ibis MHC class II B sequences. The primer sequences are not shown. The amino acid residues are given in the international single-letter code. For the variable positions, the corresponding nucleotide sequences are also given. The chicken amino acid sequence (Genbank no. AJ616893) is given as reference. A dot indicates that the amino acid or the nucleotide is the same as in the consensus sequence. An asterisk denotes an antigen binding site. Letters with shaded background denote synonymous amino acid variations. Numbering of amino acid positions is based on Brown et al. (1993).
RESULTS
All of the samples successfully amplified the 205 bp portions of the second exon of the MHC class II B genes, and SSCP gels resulted in band patterns consistent with five alleles (Fig. 2). Four overlaps in alleles between the wild and the captive population were observed, yet allele Nini *0205 was detected in only one muscle sample collected in 1990 from the wild population. Except for the rare allele Nini *0205, there was no allelic difference between different tissues. The correlation between allele distribution and sampling year is given in Table I. In the total population, allele Nini *0203 demonstrated the highest observed frequency (0.431), followed by Nini *0201 (0.292) and Nini *0202 (0.208); alleles Nini *0204 and Nini *0205 demonstrated the lowest frequency (0.056 and 0.014, respectively).
The amino acid sequences omitting the primers are given in Fig. 2, along with the corresponding nucleotide sequences for variable positions. Of the 162 bp nucleotide sequences used for alignment, 14 (8.6%) were variable. The pairwise nucleotide differences between alleles varied from 4 to 12 nucleotides, with an average of 7.4±1.9 nucleotides. Of the 54 amino acids, 16.7% (nine amino acids) variability resulted in five different alleles. The most divergent alleles (Nini *0201 and Nini *0204) differed by nine amino acids; the most similar alleles (Nini *0202 and Nini *0203) differed by only two amino acids. The average number of pairwise amino acid differences was 4.8±l.5.
For all the variable codon positions, at least two different amino acids were observed at each. In positions 74 and 75, there were both synonymous and nonsynonymous variations. The amino acid positions, documented using X-ray crystallography to be important in the antigen binding site (ABS) (Brown et al. 1993), are indicated by an asterisk in Fig. 2. In the total sequence aligned, 14 positions are ABS and 40 are not. Of the 14 ABS positions, 50.0% (7) showed variation; among the non-ABS positions, 5.0% (2 out of 40) showed variation. In the whole population, 10 times as many ABS as non-ABS are variable (0.500/0.050). The estimated rates of nonsynonymous and synonymous substitutions for ABS and non-ABS amino-acid positions are given in Table II. For the ABS positions, the nonsynonymous rate exceeded the synonymous rate, and the ratio d N /d S was 2.22, suggesting selection for diversity in these positions. For the non-ABS positions, the nonsynonymous rate was less than the synonymous rate, and the ratio d N /d S was 0.75.
DISCUSSION
MHC Variation
As in chickens, accurate determination of the number of the MHC class II genes in natural populations of birds must await the large-scale characterization of contiguous segments of DNA via cloning and sequencing (Edwards et al., 1995; Guillemont et al., 1988; Xu et al., 1989), and single loci are difficult to amplify because of the sequence similarity of priming sites caused by conversion during concerted evolution or recent duplication (Hess and Edwards, 2002). This, combined with the near absence of any MHC studies pertaining to the order Ciconiiformes, has made it difficult to identify the number of class II B genes present. Therefore, although no more than two sequences were detected within one individual, we still hold a conservative attitude to generalize that those sequences were amplified from a single locus. Our sequence-specific amplification gave a broad estimate of the MHC class II B exon 2 variation within this species, which would be the first step for an in-depth investigation of the MHC class II B genes for both the crested ibis and other species in the order Ciconiiformes.
For species that have gone through known population bottlenecks, e.g., Przewalski’s horse (Hedrick et al., 1999), the Mexican wolf (Hedrick et al., 2000a), and several pinniped species (Hoelzel et al., 1999), limited numbers of MHC alleles were observed. Similarly, only five alleles were obtained in the present study for the crested ibis, which indicates the limited genetic diversity in the MHC class II genes for this species. Obviously the limited initial genetic diversity due to the extremely small founder population plays the determinant role in causing the current low genetic diversity at these functional loci. Meanwhile, the effect of chance events is much greater in small than in large populations, which could also contribute to the current situation of low MHC diversity in the crested ibis. The disappearance of allele Nini *0205 in individuals from later sampling years might be good evidence of allele loss, or at least the transition of rare allele frequencies to an extremely low level after generations.
Moreover, documented evidence has been presented that a pattern of a low number of MHC alleles, with a high degree of divergence between alleles, was observed in bottlenecked species such as the Seychelles warbler (Richardson and Westerdahl, 2003), the Arabian oryx (Hedrick et al., 2000b), and the American bison (Mikko et al., 1997). A comparatively low level of divergence, however, between the crested ibis MHC alleles (e.g., the nucleotide differences and amino acid variations) was observed in this study, which indicates the high degree of overlap between the repertoires of peptides that the respective alleles can present.
Because MHC variants appear to be related to resistance to pathogens and parasites, MHC variation may be an excellent indicator of adaptive evolutionary potential (Hedrick, 1996). As suggested in previous studies on Atlantic salmon and Soya sheep, survival from an infection and resistance to a pathogen were associated with specific MHC alleles (Langefors et al., 2001; Lohm et al., 2002; Paterson et al., 1998). In light of the limited number of MHC alleles surviving in the crested ibis, the candidate alleles in the gene pool of this species for response to different pathogens is feared to be limited. Meanwhile, the high similarity shared between different MHC alleles would enhance the susceptibility of the whole species to certain disease infections, since diseases that evade MHC recognition in one individual are likely to evade recognition in other individuals in populations with high MHC sequence uniformity (Jeffery and Bangham, 2000). As the population of crested ibis has increased, however, there is growing evidence that reduced fitness is preventing the species from establishing self-sustaining populations. Statistics show that from 1981 to 1997, mortality due to disease was up to 39.97% in the crested ibis populations (Zhou et al., 2001a). Susceptibility to tuberculosis, septicemia, peritonitis, Escherichia coli, and a high load of helminths had directly affected the survivorship of nestlings and the viability of adults (Fan et al., 2004; Zhou et al., 2000, 2001b). Low MHC diversity does not preclude life and population growth, but it does make a population more likely to succumb to an epidemic of parasites and infectious diseases at a time of exposure to such a pathogen (Parham, 1999). Therefore, the paucity of MHC diversity observed in this study remains a real threat to the long-term survival of the crested ibis, and avoiding further loss of MHC diversity should be given the first priority in the conservation of this rare bird.
Reintroduction Implications
An understanding of a species’ genetic background is critical to successful reintroduction, for genetics can contribute information to aid in both the choice of individuals and the sites for release. MHC uniformity may increase the susceptibility of populations to pathogens and parasites; therefore, a founder population for reintroduction should be selected based on the present genetic data to contain representative samples of individuals with most of the different MHC allele genotypes. Consequently, we would hope, a pathogen capable of killing one individual may not be able to kill all in such a population. On the other hand, due to the high mortality commonly observed during the first stage of reintroduction programs, it is strategic to avoid reintroducing many of the individuals with rare MHC alleles at the very beginning, such as individuals with allele Nini *0205 or any other new rare alleles detected in further studies. When survivorship has increased, genetically more valuable individuals may be added gradually until the full range of genetic diversity in the source population is represented in the wild.
Mortality from pathogens and parasites may be a significant extinction threat for endangered species, and it is also one of the greatest challenges for most reintroduction programs. With the low genetic diversity at the MHC genes, the crested ibis population may survive some assaults but is likely to succumb to others, and it is feared that their adaptive variation potential for responding to novel pathogens may not be sufficient. Therefore, the monitoring of pathogen status of the candidate sites before release, including an epidemiological study of local avian species and a helminthological study of parasite–host relationships, is of great importance for raising the success rate of reintroduction. The crested ibis has occasionally died from infections of newcaste disease brought in by local poultry species (Fan et al., 2001). Such occurrences make the environmental pathogen study more necessary and urgent. It is hoped that reintroduction projects may be developed according to the recommendations of this paper, and that efforts taken for the recovery of this highly endangered bird will both enrich its own conservation and be applied to wider conservation biology to the aid of all endangered species that have experienced a similar history of genetic bottlenecks.
REFERENCES
Bassam, B. J., Caetano-Anolles, G., and Gresshoff, P. M. (1991). Fast and sensitive silver staining of DNA in polyacrylamide gels. Anal. Biochem. 196(1):80–83.
Bird Life International (2001). Threatened Birds of Asia: The Bird Life International Red Data Book, Bird Life International, Cambridge, UK.
Brown, J. H., Jardetzky, T. S., Gorga, J. C., Stem, L. J., Urban, R. G., Strominger, J. L., and Wiley, D. C. (1993). Three-dimensional structure of the human class II histocompatibility antigen HLA-DRI. Nature 364:33–39.
Ding, C. Q., and Liu, D. P. (2002). Crested Ibis Nipponia nippon. Chin. J. Zool. 37:84.
Edwards, S. V., Grahn, M., and Potts, W. K. (1995). Dynamics of Mhc evolution in birds and crocodilians: Amplification of class II genes with degenerate primers. Mol. Ecol. 4:719–729.
Edwards, S. V., and Potts, W. K. (1996). Polymorphism of gene in the major histocompatibility complex (MHC): Implications for conservation genetics of vertebrates. In Smith, T. B., and Wayne, R. K. (eds.), Molecular Genetic Approaches in Conservation, Oxford University Press, New York, pp. 214–236.
Fan, G. L., Yang, Z. Q., Gao, G. G., Bin, J. J., Qiao, H. L., Huang, Z. X., and Hou, Y. F. (2004). The histopathological observations of young crested ibis infected with Escherichia coli. Chin. J. Zool. 39(3):44–46.
Fan, G. L., Zhou, H. C., Yang, M. Q., Pu, P., Yang, Z. Q., Cao, Y. H., Fu, W. K., and Lu, B. Z. (2001). The pathological observation of young crested ibis newcaste disease. J. Northwest Sci. Tech. Univ. Agric. For. (Nat. Sci. Ed.) 29(6):79–82.
Frankham, R., Ballou, J. D., and Briscoe, D. A. (2002). Introduction to Conservation Genetics, Cambridge University Press, Cambridge, UK.
Guillemot, F., Billault, A., Pourquie, O., Behar, G., Chausse, A. M., Zoorob, R., Kreibich, G., and Auffray, C. (1988). A molecular map of the chicken major histocompatibility complex: The class II genes are closely linked to the class I genes and the nucleolar organizer. EMBO J 7(9):2775–2785.
Hancock, J. A., Kushlan, J. A., and Kahl, M. P. (1992). Storks, Ibises, and Spoonbills of the World, Academic, London.
Hedrick, P. W. (1996). Conservation genetics and molecular techniques: A perspective. In Smith, T. B., and Wayne, R. K. (eds.), Molecular Genetic Approaches in Conservation, Oxford University Press, New York, pp. 459–477.
Hedrick, P. W., and Kim, T. J. (2000). Genetics of complex polymorphisms: Parasites and maintenance of MHC variation. In Singh, R. S., and Krimbas, C. B. (eds.), Evolution Genetics: From Molecules to Morphology, Cambridge University Press, Cambridge, UK, pp. 204–234.
Hedrick, P. W., Lee, R., and Parker, K. M. (2000a). Major histocompatibility complex (MHC) variation in the endangered Mexican wolf and related canids. Heredity 85:617–624.
Hedrick, P. W., Parker, K. M., Gutierrez-Espeleta, G. A., Rattink, A., and Lievers, K. (2000b). Major histocompatibility complex variation in the Arabian oryx. Evolution 54:2145–2151.
Hedrick, P. W., Parker, K. M., Miller, E. L., and Miller, P. L. (1999). Major histocompatibility complex variation in the endangered Przewalski’s horse. Genetics 152:1701–1710.
Hess, C. M., and Edwards, S. V. (2002). The evolution of the major histocompatibility complex in birds. Bioscience 52(5):423–431.
Hoelzel, A. R., Stephens, J. C., and O’Brien, S. J. (1999). Molecular genetic diversity and evolution at the MHC DQB locus in four species of pinnipeds. Mol. Biol. Evol. 16(5):611–618.
Jeffery, D. J. M., and Bangham, C. R. M. (2000). Do infectious diseases drive MHC diversity? Microb. Infect. 2:1335–1341.
Jukes, T. H., and Cantor, C. R. (1969). Evolution of protein molecules. In Munro, H. N. (ed.), Mammalian Protein Metabolism, Academic, New York, pp. 21–132.
Kumar, S., Tamura, K., Jakobsen, I. B., and Nei, M. (2001). MEGA2: Molecular Evolutionary Genetics Analysis Software. Arizona State University, Tempe.
Langefors, A., Lohm, J., Grahn, M., Andersen, O., and von Schantz, T. (2001). Association between major histocompatibility complex class II B alleles and resistance to Aeromons salmonicida in Atlantic salmon. Proc. R. Soc. Lond. B 268:479–485.
Liu, Y. Z. (1981). Rediscovery of crested ibis Nipponia nippon in Qinling Mountain. Chin. J. Zool. 27:237.
Lohm, J., Grahn, M., Langefors, A., Andersen, O., Storset, A., and von Schantz, T. (2002). Experimental evidence for major histocompatibility complex allele-specific resistance to a bacterial infection. Proc. R. Soc. Lond. B 2114:2029–2034.
Lokki, M. L., and Laitinen, T. (2001). Role of major histocompatibility complex class III genes in recurrent spontaneous abortions. Front. Biosci. 6:E23–E29.
Mikko, S., Spencer, M., Morris, B., Stabile, S., Basu, T., Stormont, C., and Andersson, L. (1997). A comparative analysis of Mhc DRB3 polymorphism in the American bison (Bison bison). J. Hered. 88:499–503.
Nei, M., and Gojobori, T. (1986). Simple methods for estimating the numbers of synonymous and nonsynonymous nucleotide substitutions. Mol. Biol. Evol. 3:418–426.
Parham, P. (1999). Virtual reality in the MHC. Immunol. Rev. 167:5–15.
Paterson, S., Wilson, K., and Pemberton, J. M. (1998). Major histocompatibility complex variation associated with juvenile survival and parasite resistance in a large unmanaged ungulate population (Ovis aries L.). Proc. Natl. Acad. Sci. U.S.A. 95:3714–3719.
Penn, D. J., and Potts, W. K. (1998). How do major histocompatibility complex genes influence odor and mating preferences? Adv. Immunol. 69:411–436.
Richardson, D. S., and Westerdahl, H. (2003). MHC diversity in two Acrocephalus species: The outbred great reed warbler and the inbred Seychelles warbler. Mol. Ecol. 12:3523–3529.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual, 2nd edn., Cold Spring Harbor Laboratory Press, New York.
Sunnucks, P., Wilson, A. C. C., Beheregaray, L. B., Zenger, K., French, J., and Taylor, A. C. (2000). SSCP is not so difficult: The application and utility of single-stranded conformation polymorphism in evolutionary biology and molecular ecology. Mol. Ecol. 9:1699–1710.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., and Higgins, D. G. (1997). The Clustal X Windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25:4876–4882.
Wang, S. (1998). China Red Data Book of Endangered Animals: Aves, Science Press, Beijing.
Xi, Y. M., Lu, B. Z., and Fujihara, N. (2001). Captive rearing and breeding of the crested ibis, Nipponia nippon. J. Poult. Sci. 38:213–224.
Xi, Y. M., Lu, B. Z., Zhang, Y. M., and Fujihara, N. (2002). Restoration of the crested ibis, Nipponia nippon. J. Appl. Anim. Res. 22:193–200.
Xu, Y. X., Pitcovski, J., Peterson, L., Auffray, C., Bourlet, Y., Gerndt, B. M., Nordskog, A. W., Lamont, S. J., and Warner, C. M. (1989). Isolation and characterization of three class II major histocompatibility complex genomic clones from the chicken. J. Immunol. 142(6):2122–2132.
Yamashina, Y. (1975). The feeding of Japanese crested ibis. In Temple, S. A. (ed.), Endangered Birds, University of Wisconsin Press, Madison, pp. 161–164.
Zhang, B., Fang, S. G., and Xi, Y. M. (2004). Low genetic diversity in the endangered crested ibis Nipponia nippon and implications for conservation. Bird Conserv. Int. 14:183–190.
Zhou, H. C., Fan, G. L., Cao, Y. H., Fu, W. K., Lu, B. Z., and Xi, Y. M. (2000). The pathological diagnosis of the death of a domesticated crested ibis. Acta Univ. Agric. Boreali-Occidentalis 28(2):60–63.
Zhou, H. C., Fan, G. L., Lin, Q., Yang, M. Q., Zhang, L. L., Cao, Y. H., Lou, X. R., and Lu, B. Z. (2001a). The pathological observations of crested ibis Eustrongyloides sp. J. Northwest Sci. Tech. Univ. Agric. For. (Nat. Sci. Ed.) 29(5):27–29.
Zhou, H. C., Yang, M. Q., Fan, G. L., Fu, W. K., Cao, Y. H., Li, H. L., and Zheng, J. J. (2001b). The pathological diagnosis of the death of the two crested ibis. J. Northwest Sci. Tech. Univ. Agric. For. (Nat. Sci. Ed.) 29(3):69–72.
ACKNOWLEDGMENTS
We are grateful to Shaanxi Crested Ibis Breeding Centre for providing all the samples used in this study, as well as to Dr. Wan Q.H., Dr. Wu H., Dr. Wu H.L., Dr. Xu Y.C., and Mr. Wood for their valuable comments on the manuscript. This work was supported by a grant from the State Key Basic Research and Development Plan of P.R. China (No. G2000046906) and a special grant for crested ibis from the State Forestry Administration, P.R. China.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhang, B., Fang, SG. & Xi, YM. Major Histocompatibility Complex Variation in the Endangered Crested Ibis Nipponia nippon and Implications for Reintroduction. Biochem Genet 44, 110–120 (2006). https://doi.org/10.1007/s10528-006-9018-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10528-006-9018-1