
Abstract
We examined the influence of crop management (crop density, nitrogen fertilization level) and landscape parameters [areal proportion of oilseed rape (OSR), Brassica napus L. var. oleifera Metzg. (Brassicaceae), and proportions, lengths and distances of various non-crop habitats] on the parasitism of stem weevils (Ceutorhynchus spp.; Coleoptera: Curculionidae) and pollen beetles (Meligethes spp.; Coleoptera: Nitidulidae) by parasitic wasps (Hymenoptera: Ichneumonidae). Larvae of host insects were sampled in 29 winter OSR fields located in landscapes ranging from structurally simple to complex. Spatial scales of landscape effects were considered at eight radii (250–2,000 m) centered in the studied OSR fields. Stem weevil parasitism was unaffected by crop management variables but negatively related to roadside strip length at radius 250 m and grassy fallow area at a radius of 500 m. Pollen beetle parasitism was positively related to the crop density and to lengths of roadside strips and hedges within a radius of 250 m. Thousand kernel mass of OSR was positively related to pollen beetle parasitism but unrelated to stem weevil parasitism.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Parasitoids are important natural enemies of many crop pests and may act as keystone species in agroecosystems (LaSalle 1993; Tscharntke 2000). Landscape structure has been shown to directly affect biological control involving parasitoids as antagonists of various pest insects (Corbett and Rosenheim 1996; Roland and Taylor 1997; Menalled et al. 1999; Thies and Tscharntke 1999; Östman et al. 2001). Studies showed that the efficacy of parasitoids may be altered by both changing agricultural practices within crop fields (Wratten and Van Emden 1995; Van Driesche and Bellows 1996; Östman et al. 2001) and by management of agricultural landscapes (Burel and Baudry 1995; Van Driesche and Bellows 1996; Tscharntke and Kruess 1999). However, most of these pest–parasitoid interactions have been studied on a single species pair by focusing either on the impact of site or landscape characteristics, but only rarely included investigations of both site and landscape effects and more than one parasitoid/pest species pair (Thies and Tscharntke 1999; Östman et al. 2001).
Oilseed rape (OSR), Brassica napus L. var. oleifera Metzg. (Brassicaceae), acreage is increasing worldwide (http://faostat.fao.org/site/567/default.aspx) and in many regions accompanied by a dramatic disproportionate increase of pesticide applications (Gianessi and Marcelli 2000). Stem-boring weevils [cabbage stem weevil, Ceutorhynchus pallidactylus Marsh., and rape stem weevil, C. napi Gyll. (both Coleoptera: Curculionidae)] and pollen beetles [Meligethes aeneus Fabr. and M. viridescens Fabr. (both Coleoptera: Nitidulidae)] are among the major insect pests in OSR across Europe (Alford 2003). Cabbage and rape stem weevils are mainly attacked by the univoltine, koinobiont parasitoids Tersilochus obscurator Aub. and T. fulvipes Grav. (Hymenoptera: Ichneumonidae), respectively (Ulber 2003), that lay their eggs through the stem and petiole tissue into the host larvae while these are mining within the stem pith. Pollen beetle larvae develop in rape flowers to the last instar and are mainly attacked by the specialized univoltine, koinobiont parasitoid T. heterocerus Aub. (Nilsson 2003). After the full-grown stem weevil and pollen beetle larvae have left the plant to pupate in the soil, these parasitoid species develop rapidly and kill the host prepupae; the adult parasitoid diapauses in its pupal cocoon until it emerges in the following spring and migrates to new oilseed rape crops (Nilsson 2003; Ulber 2003).
In the current study, we investigated the effects of crop management and landscape factors on parasitism of pollen beetles and stem weevils in 29 winter OSR fields that were embedded in a region comprising structurally simple (primarily cropland) to complex landscapes (cropland intermixed with various non-crop habitats). Because hatching parasitoids must locate new plants wherein to locate host larvae, we hypothesized that the spatial distribution and proportion of the host habitat and the abundance of hosts will affect parasitism rates (Thies et al. 2008). Non-crop habitats have been shown to enable populations of parasitoids to build-up over years and to enhance parasitoid dispersal into the OSR fields (Thies and Tscharntke 1999). However, while the general influence of areal proportions of non-crop habitats have been demonstrated, very little is known on how more linear non-crop habitats like roadside strips or hedges and the distances of these habitats to crop fields affect parasitism rates. Since parasitoid species in OSR are unable to migrate long distances (Nilsson 2003), we hypothesized that mainly structures in the vicinity of OSR fields will affect parasitism rates. Management effects like ploughing have been shown to negatively affect parasitoid populations in OSR because they are hibernating in the soil of the crop field (Klingenberg and Ulber 1994). Moreover, we hypothesized that parasitization can indirectly also be influenced by management measures that alter crop growth and crop density. Possible mechanisms for indirect crop management effects can be that (1) better fertilized crops grow faster and show a denser morphology leading to a lower parasitism rate via an increased searching time, (2) an altered crop density may also change microclimatic conditions within the crop stand, influencing parasitoid behaviour, and (3) differently fertilized crop plants may produce different olfactory and visual signals perceived by parasitoids and influence their searching behaviour and ultimately parasitism rates (Ferguson et al. 2006; Jönsson et al. 2007; Williams et al. 2007). Nitrogen availability in the soil has also been shown to increase OSR crop vigour and its ability to compensate for pollen beetle damage (Valantin-Morison et al. 2007).
In the current study we also investigated whether yield parameters would be affected by parasitoids, although pest larvae will be killed by parasitoids after they have already caused damage to the crop and possible effects on yield would most likely be detected in the following season only. Nevertheless, relationships between parasitism and crop yield could give indications to underlying tri-trophic plant–herbivore–parasitoid interactions mediated through plant conditions (Tscharntke 2000).
The specific objectives of the study were to determine (1) whether parasitism rates are dependent on pest abundance, (2) what specific crop management and/or landscape characteristics influence the parasitism of stem weevils and pollen beetles, (3) at which spatial scale the potential influence of landscape characteristics is effective, and (4) whether OSR yield is affected by parasitization of these pests. Considered crop management factors included nitrogen fertilization rate and crop density; landscape factors were assessed at eight radii (250–2,000 m) centered in the study field and included proportions of OSR, grassy fallows, woody areas, length of roadside strips and hedges and distance of the study field to the nearest grassy fallow and woody area.
Materials and methods
Study area, OSR fields and predictor variables
The study region (area about 240 km2) is located about 40 km east of Vienna, Austria (altitudes of arable land ranging from 130 to 250 m; coordinates of the central area: 16°57′ E, 48°04′ N). During the study period from April to July 2005 the average temperature was 16.9°C; the precipitation during this period amounted to 336 mm (average temperature and precipitation in these months between 1990 and 2004 was 16.0°C and 230 mm, respectively; weather data were provided by the meteorological station Bruck/Leitha located about 16 km west of the study region). Farmland in the region is located on chernozem soils and cropped according to integrated pest management guidelines (main crops: wheat, maize, barley, OSR, sunflower, sugar beet, poppy seed, grape vine) and is interspersed with semi-natural non-crop areas such as fallows, hedges and forest remnants. Within this region, we randomly selected 29 OSR fields located in landscapes comprising a complexity gradient ranging from structurally simple to more complex for collecting stem weevils and pollen beetles [see Zaller et al. (2008a, b) for further details]. Landscape variables calculated for the surroundings of each study field were areal proportions of OSR fields, grassy fallows and woody areas, length of roadside strips and hedges, and distance of the studied OSR fields to the nearest grassy fallow and woody area (Table 1). Scale effects were considered by calculating landscape variables at eight circular sectors (radius 250, 500, 750, 1,000, 1,250, 1,500, 1,750 and 2,000 m) around each study field using the software packages ArcGis 9.1 and ArcView GIS 3.3 (ESRI, Redlands, CA, USA). Field surveys were conducted in 2005 to assess the current landscape composition.
Oilseed rape (cv Californium) was sown by the farmers in late summer 2004 at an average seeding rate of 3.9 ± 0.1 kg ha−1 (mean ± SE). OSR fields (average field area 2.2 ± 2.6 ha) were fertilized and treated with herbicides and insecticides until December 2004. From January 2005 onwards, 1 ha of these OSR fields was excluded from pesticide applications and used for sampling insect pests and crop plants for the current investigation. A buffering zone of at least 10 m to the next sprayed OSR was kept.
Crop management variables included were crop density and nitrogen fertilization level because of their potential influence on parasitoid behaviour through altering stand microclimate and host plant quality or through effects on pest populations and apparency. Crop density was assessed by counting the number of OSR stems within two 1-m2 frames per field in late June 2005; nitrogen fertilization levels were obtained by a questionnaire among the participating farmers (Table 1).
Pests, parasitoids and yield parameters (response variables)
Pest sampling included (1) the cabbage and rape stem weevil larvae that tunnel in the stems and (2) pollen beetle larvae that feed on pollen in OSR flowers (Alford et al. 2003). Cabbage and rape stem weevil larvae were sampled in late April 2005 (growth stage, GS 64–67; Lancashire et al. 1991) by removing 25 randomly chosen OSR plants along a 50-m transect located in the central area of each 1-ha study plot; stems were dissected subsequently and weevil larvae therein carefully removed and counted. Pollen beetle larvae were sampled during flowering of OSR in late April 2005 (GS 64–65) at sunshine between 10 a.m. and 2 p.m. on 25 randomly chosen OSR plants along a 50-m transect located in the central area of each study plot by putting a plastic bag over the top raceme, cutting off the raceme and subsequently counting larvae in the laboratory.
Parasitism was investigated by rearing a given number of second instar pollen beetle and second to third instar stem weevil larvae (mostly 100, fewer if not sufficient large larvae were available) in separate plastic boxes containing moistened peat-free commercial garden substrate (Williams et al. 2003). Host larvae were placed onto the surface of the substrate where they immediately burrowed down in order to pupate. The boxes were kept at room conditions for up to three months to allow unparasitized adult insect pests to emerge. Emerging adult stem weevils or pollen beetles were removed and counted continuously. After 12 weeks, pupal cocoons of pollen beetle and stem weevil parasitoids were counted after thoroughly searching through the substrate using a dissecting microscope. Parasitism rate was calculated by dividing the number of parasitoid cocoons by the number of pest larvae placed into boxes.
OSR yield (g m−2) and thousand kernel mass was determined after harvesting on two randomly chosen 1-m2 areas of each OSR field in the 3rd week of June 2005 (Zaller et al. 2008a).
Statistical analysis
First, we tested response variables for normality using Shapiro–Wilk W-statistics. Only pest abundance data were log-transformed to meet criteria for regression analyses (Zar 1996). Second, we correlated pairwise the two crop management and seven landscape variables at each of the eight spatial scales to detect intercorrelations among predictor variables (Pearson correlations; P < 0.05). While the two crop management variables were unrelated (R = 0.161, P = 0.405), we found significant correlations between landscape variables at all spatial scales (Table 2). However, in order to be able to assess the effect of specific non-crop elements on pest parasitism we decided to include all variables stepwise forward in the subsequent multivariate analyses but then removed the least significant one of two intercorrelated variables (see below). Third, effects of crop management and landscape variables at different spatial scales on stem weevil parasitism rates were analyzed separately at each of the eight radii using ordinary least-squares (OLS) multiple regression models. Pollen beetle parasitism occurred only in seven of 29 study fields, therefore we analyzed these data (presence and absence of parasitism) using a generalized linear model (GLM) with a binomial error distribution and a logistic link function. Significant predictor variables were selected after performing a forward stepwise selection followed by a backward elimination procedure (Yee and Mitchell 1991). At each step of variable selection the significance of the partial effects was tested by dropping each predictor term from the model (Wald test, F-statistics, P < 0.05). Possible non-linear effects were accounted for by using restricted quadratic and cubic spline functions (Harrel 2001). To examine possible spatial autocorrelations in the residuals of our OLS and GLM models, we used Moran’s I correlograms at different spatial scales (Legendre and Legendre 1998; Lichstein et al. 2002). None of the tested lag distances showed significant standardized I-values indicating that no spatial autocorrelation remained among study fields after crop management and landscape variables were included in the model. Fourth, we conducted Pearson correlations to test relationships between pest abundance, yield parameters and parasitism rates. All statistical analyses were performed using S-PLUS 7.0 for Windows (Insightful, Seattle, WA, USA) and SPSS 15.0 for Windows (SPSS, Chicago, IL, USA).
Results
Mean abundances (±SE) of stem weevil and pollen beetle larvae were 30 ± 4 and 1,797 ± 100 m−2, respectively. Parasitism of stem weevil larvae (3.50 ± 0.45% larvae parasitised) was not correlated with numbers of stem weevil larvae m−2 (R = 0.14, P = 0.466), parasitism of pollen beetle larvae (0.48 ± 0.25% larvae parasitised) was significantly positively correlated with abundance of pollen beetle larvae m−2 (R = 0.42, P = 0.029).
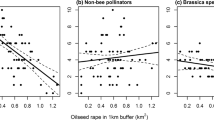
For stem weevil parasitism, OLS analyses yielded only two univariate models. At a radius of 250 m around the study fields, stem weevil parasitism was significantly negatively related to the length of roadside strips (R 2 = 0.16, F 1,27 = 4.84, P = 0.032) and at a radius of 500 m significantly negatively related to second polynomial of the proportion of grassy fallows (R 2 = 0.22, F 2,26 = 4.74, P = 0.037; Fig. 1). There were no significant multivariate models. Pollen beetle parasitism was best explained by a single multivariate GLM model (full model R 2 = 0.70) with significant positive relations to crop density (partial R 2 = 0.62, P < 0.001), roadside strip length within 250 m (partial R 2 = 0.50, P < 0.001) and the second polynomial of hedge length within 250 m radius (partial R 2 = 0.26, P = 0.017; for the sake of clarity, only the two most important variables are shown in Fig. 2). The plot of the partial effects (Fig. 2) indicated that the probability for pollen beetle parasitism was high at high crop densities and/or length of roadside strips but parasitism probability was low at low crop densities and/or short roadside strip length (Fig. 2). No significant relationships were found for landscape variables at radii >250 m.

Relationships between parasitism of stem weevils (C. pallidactylus + C. napi) and the two best explaining variables derived from stepwise multivariate OLS regression analyses conducted at each spatial scale including two crop management and seven landscape variables. Lines indicate quadratic fits through data points

Probability of parasitism of pollen beetles (Meligethes spp.) by hymenopteran parasitoids in relation to the best explaining variables derived from stepwise binomial multivariate GLM regression analyses conducted at each spatial scale including two crop management and seven landscape variables
OSR yield was not correlated with parasitism rates of stem weevils (R = 0.043, P = 0.826) and only marginally significantly negatively correlated with pollen beetle parasitism (R = −0.344, P = 0.068). However, thousand kernel mass was significantly positively correlated with the occurrence of pollen beetle parasitism (R = 0.617, P < 0.001) but unrelated to stem weevil parasitism (R = −0.289, P = 0.128; Fig. 3).

Relationships between OSR thousand kernel mass and parasitism of stem weevils and pollen beetles. Line indicates the fit of the Pearson correlation
Discussion
This study provides evidence from 29 OSR fields and two major pest insects of species-specific effects on parasitism caused by host larvae abundance, crop management or landscape variables.
For pollen beetles, our findings are the first reporting a positive relationship between parasitism and pollen beetle abundance, while in the literature results are inconsistent ranging from a negative relationship between parasitism and host density (Billqvist and Ekbom 2001) to a lack of such a relationship (Büchi 2002; Thies et al. 2003). Stem weevil parasitism was unrelated to the abundance of their host larvae. In view of the low parasitism rates this implies that the parasitoid population size was not sufficient enough to exploit the greater abundance of hosts. Generally, direct relationships between the abundance of stem weevils and their parasitoids have rarely been investigated and published results seem contradictory showing decreasing numbers of C. pallidactylus with increasing numbers of its larval endoparasitoid T. obscurator (Barari et al. 2005) due to a lack of an association between stem weevils and their parasitoids (Ferguson et al. 2006). An explanation for these species-specific relationships might be that pollen beetle larvae located in the flowers are much easier to find for the parasitoids than the stem weevils mining in the OSR petioles and stems. As a consequence, T. heterocerus might respond more directly to changes in host abundance than T. obscurator or T. fulvipes. Our findings also suggest that the more exposed pollen beetles might be more prone to disturbance or interference by parasitoids than the stem-mining weevils that did not show such a relationship. Discrepancies between findings in different studies can also be explained by regional differences of these relationships and different sampling and study designs used. Further studies are needed to elucidate the underlying mechanisms.
In a previous study conducted in this region, we could show that pollen beetles and stem weevils responded to OSR area in the surroundings (Zaller et al. 2008b; Moser et al. 2009). In the current study, however, parasitoids were unaffected by OSR area in the surroundings supporting earlier results of Thies et al. (2003) but contrasting more recent findings for pollen beetle parasitoids (Thies et al. 2008). An explanation for these inconsistent results could be that regional differences in the relationships to landscape mosaics are more pronounced for parasitoids than for pest species insects because parasitoids may have a very limited dispersal ability and may not redistribute considerably among available OSR area in a landscape from their overwintering sites (Nilsson 2003).
Parasitism of pollen beetle larvae was influenced by OSR crop density and the length of roadside strips and hedges. Thus, the greatest probability of pollen beetle parasitism was seen when pollen beetle abundance was high and additionally when both crop densities and length of roadside strips and hedges were also at high levels, whereas the lowest probability for parasitism was found when crop density and length of roadside strips and hedges was low. This suggests that higher pollen beetle abundance facilitated host searching by the parasitoids (Maini et al. 1991; Wang et al. 1997). This taken together with our previous finding that pollen beetle abundance was highest at medium crop densities (Zaller et al. 2008b) indicates that in dense OSR stands other factors than host abundance might increase parasitism. A previous study showed a general positive relationship between parasitism of pollen beetles and non-crop area (Thies et al. 2003). However, the current study is to our knowledge the first one showing which types of non-crop element might actually be responsible for this relationship. Parasitoid populations of pollen beetles may have benefited from the availability of non-crop areas in several ways. First, because the parasitoids hibernate in the soil of the previous years’ crop and are known to be negatively affected by agricultural practices like ploughing (Nilsson 1985). Non-crop areas offer supply of pollen and nectar for adult parasitoids and should therefore better enable populations to build up over years. Second, although roadside strips and hedges are non-crop elements that comprise only a very small area in the studied landscapes (on average 1 and 6 m wide, respectively; <1% area at radius 500 m), they form a network of diverse (around 50 annual and perennial plant species) perennial habitats in this arable landscape that support alternative hosts and host plants. It is not clear, however, whether and to what extend these linear structures in the landscapes are actually utilized by hymenopteran parasitoids. From observations we know that wild Brassicaceae and other non-cruciferous herbs inhabiting these roadside strips and hedges are frequently utilized by Meligethes species and most likely also by its parasitoids (see also Hoffmann and Schmutterer 1999). Other observations underline the important role of non-crop elements for univoltine parasitoids in OSR as important nectar sources enabling the parasitoids to maintain adequate food reserves when migrating from hibernation sites to oilseed rape crops (Nilsson 2003), leading to enhanced parasitism and an extended parasitoid lifetime (Wratten and Van Emden 1995). Moreover, we could demonstrate in a previous study that these linear habitats are also important for other beneficial organisms like predatory agrobiont spiders (Drapela et al. 2008). These data suggest that non-crop elements surrounding OSR fields may serve as refugial habitats for parasitoid populations (Pulliam 1988) and indicate that the specific composition of different landscape elements can contribute to the degree of parasitism.
In contrast to the parasitism of pollen beetles, parasitism of stem weevils was unaffected by crop management factors. In a previous study only soil quality affected abundances of pollen beetles and stem weevils while the crop management factor nitrogen fertilization had no effect (Zaller et al. 2008b) suggesting that insect pests are differently affected by crop management than their antagonists. The current data are to our knowledge the first showing that landscape might affect parasitism of stem weevils in OSR. The negative association with grassy fallow area in the surroundings seems to be in contrast to findings for the closely related parasitoids of pollen beetle that responded positively to non-crop area (Thies et al. 2003). This suggests that the parasitoids’ response to non-crop landscape elements differ between species. Another explanation could be that parasitoid–landscape interactions are specific for each region, therefore non-crop elements not present in our landscapes (e.g., meadows, pastures) could have positively affected parasitism rates in the other study. A further explanation could be that the study of pest–parasitoid interactions on isolated potted OSR plants located within fallow areas by Thies et al. (2003) delivered results not well comparable with those from our field-grown OSR plants. For example, if a few potted rapeseed plants are located within a fallow area, proportions between crop plants and the surrounding landscape are very different from the natural situation in the landscape. Moreover, microclimatic and nutritional conditions of the potted OSR plants will be very different from OSR plants in crop fields and thereby differently affect parasitoid–pest–crop interactions.
As expected, parasitism of both pest species groups responded to non-crop structures at rather small scales (250 and 500 m for pollen beetles and stem weevils, respectively). This seems to be in contrast to Thies et al. (2003) who found strongest correlations between pollen beetle parasitism and non-crop areas at a scale of 1,500 m. A limitation of hosts available at certain scales seemed unlikely to be responsible for these relationships because abundances of stem weevils and pollen beetles were generally high in the area (Zaller et al. 2008b). Thus, dispersal limitations of parasitic wasps might be the reason for these patterns (With et al. 1999). However, no reliable data are available on the dispersal distance of parasitoids of OSR pest insects that could substantiate this explanation.
Parasitism levels observed in the current study were rather low in comparison to those reported for pollen beetles that in the literature range from 0 to 81% (Billqvist and Ekbom 2001; Büchi 2002; Kraus and Kromp 2002) and for stem weevils from 8 to 81% (Kraus and Kromp 2002; Barari et al. 2005; Ferguson et al. 2006). Generally, parasitism levels have been shown to vary considerably between years and study regions (Nitzsche and Ulber 1998; Billqvist and Ekbom 2001; Büchi 2002; Kraus and Kromp 2002). However, additional explanations are possible. First, because adult female parasitoids only live for a few days (Nilsson 2003) unfavourable weather conditions during this time can be detrimental for their success to parasitize host larvae. In our region, precipitation during the study period was 40% higher than the long-term average, indicating that conditions for parasitization might have been unfavourable in this season. Second, it is known that parts of the host larval population will escape parasitization in some years when the coincidence between crop development and parasitoid appearance is not precise (Nilsson 2003). Third, the low parasitism rates in the current study could be due to the chosen method in which host larvae were randomly sampled and allowed to bury and pupate in soil substrate. With this method, estimates of parasitism rates are affected by host larvae mortality not due to predation or parasitism that can be remarkably high (e.g., for pollen beetle mortality up to 72%) (Büchi 2002). In contrast, parasitism data based on dissections of host larvae might overestimate the impact of parasitoids because it does not include mortality not due to parasitism (Büchs and Nuss 2000). In the current study, we were unable to address species-specific effects of crop management and landscape on parasitoids because we did not identify the parasitoid species reared from the respective host larvae. This parasitoid species-specific relationships might merit further studies aiming to better understand their specific ecological impact provided that overall parasitism rates are higher than in the current study.
The lack of a relationship between OSR yield and parasitism reflects the koinobiont life strategy of these parasitoids whose effects would most likely be detected in the following season. However, our findings on relationships between pollen beetle parasitism and OSR thousand kernel also suggests that other effects of parasitoids beside parasitism, e.g., disturbance/interference, might also play a role in reducing damage by pollen beetles (Zaller et al. 2008a). Relationships between parasitation and thousand kernel mass might also be the result of carry-over effects of pest–parasitoid interactions from previous years assuming that pest populations were considerably reduced by parasitoids in the years before. However, the most probable explanations could be that a higher thousand kernel mass is the result of better-nourished OSR plants leading to more host larvae for parasitoids, or that OSR plants loose their flower buds (equivalent to pollen beetle damage) and compensated for damage by increasing the kernel mass (Williams and Free 1979).
Our findings on the effect of crop management and landscape variables on parasitism of pests can have implications for integrated pest management strategies that aim to incorporate parasitoids into conservation biological control (Williams 2004) as they suggest that successful conservational biocontrol would mainly be effective by a concerted action of on-site measures and landscape management. However, before predictions on the influence of both crop and landscape management on pest–parasitoid interactions can be made more data from different regions including life cycles, dispersal abilities and searching behaviour of parasitoids seem imperative.
References
Alford DV (2003) Biocontrol of oilseed rape pests. Blackwell, Oxford
Alford DV, Nilsson C, Ulber B (2003) Insect pests of oilseed rape crops. In: Alford DV (ed) Biocontrol of oilseed rape pests. Blackwell, Oxford, pp 9–42
Barari H, Cook SM, Clark SJ, Williams IH (2005) Effect of a turnip rape (Brassica rapa) trap crop on stem-mining pests and their parasitoids in winter oilseed rape (Brassica napus). BioControl 50:69–86
Billqvist A, Ekbom B (2001) The influence of host plant species on parasitism of pollen beetles (Meligethes spp.) by Phradis morionellus. Entomol Exp Appl 98:41–47
Büchi R (2002) Mortality of pollen beetle (Meligethes spp.) larvae due to predators and parasitoids in rape fields and the effect of conservation strips. Agric Ecosyst Environm 90:255–263
Büchs W, Nuss H (2000) First steps to assess the importance of epigaeic active polyphagous predators on oilseed rape insect pest with soil dwelling larvae. IOBC/WPRS Bull 23:151–163
Burel F, Baudry J (1995) Farming landscapes and insects. In: Glen DM, Greaves M, Anderson HP (eds) Ecology and integrated farming systems. Wiley, London, pp 203–220
Corbett A, Rosenheim JA (1996) Impact of natural enemy overwintering refuge and its interaction with the surrounding landscape. Ecol Entomol 21:155–164
Drapela T, Moser D, Zaller JG, Frank T (2008) Spider assemblages in winter oilseed rape affected by landscape and site factors. Ecography 31:254–262
Ferguson AW, Barari H, Warner DJ, Campbell JM, Smith ET, Watts NP, Williams IH (2006) Distributions and interactions of the stem miners Psylliodes chrysocephala and Ceutorhynchus pallidactylus and their parasitoids in a crop of winter oilseed rape (Brassica napus). Entomol Exp Appl 119:81–92
Gianessi LP, Marcelli MB (2000) Pesticide use in U.S. crop production: 1997. National Center for Food and Agricultural Policy, Washington, DC
Harrel FE (2001) Regression modeling strategies. Springer, New York
Hoffmann C, Schmutterer H (1999) Parasitäre Krankheiten und Schädlinge an landwirtschaftlichen Kulturpflanzen. Ulmer Verlag, Stuttgart
Jönsson M, Rosdahl K, Anderson P (2007) Responses to olfactory and visual cues by over-wintered and summer generations of the pollen beetle, Meligethes aeneus. Physiol Entomol 32:188–193
Klingenberg A, Ulber B (1994) Untersuchungen zum Auftreten der Tersilochinae (Hym.; Ichneumonidae) als Larvalparasitoiden einiger Rapsschädlinge im Raum Göttingen 1990 und 1991 und zu deren Schlupfabundanz nach unterschiedlicher Bodenbearbeitung. J Appl Entomol 117:287–299
Kraus P, Kromp B (2002) Parasitization rates of the oilseed rape pests Ceutorhynchus napi, Ceutorhynchus pallidactylus (Coleoptera, Curculionidae) and Meligethes aeneus (Coleoptera, Nitidulidae) by Ichneumonids in several localities of eastern Austria. IOBC/WPRS Bull 25:117–122
Lancashire PD, Bleiholder T, Van den Boom T, Langelüddecke P, Stauss R, Weber E, Witzenberger A (1991) A uniform decimal code for growth stages of crops and weeds. Ann Appl Biol 119:561–601
LaSalle J (1993) Parasitic hymenoptera, biological control and biodiversity. In: LaSalle J, Gauld ID (eds) Hymenoptera and biodiversity. CABI, Wallington, pp 197–215
Legendre P, Legendre L (1998) Numerical ecology, 2nd edn. Elsevier, Amsterdam
Lichstein JW, Simons TR, Shriner SA, Franzreb KE (2002) Spatial autocorrelation and autoregressive models in ecology. Ecol Monogr 72:445–463
Maini S, Burigo G, Carrieri M (1991) Trichogramma maidis host-searching in corn vs. pepper. Redia 74:121–127
Menalled FD, Marino PC, Gage SH, Landis DA (1999) Does agricultural landscape structure affect parasitism and parasitoid diversity? Ecol Appl 9:634–641
Moser D, Drapela T, Zaller JG, Frank T (2009) Interacting effects of wind direction and resource distribution on rape pest species abundance. Basic Appl Ecol doi:10.1016/j.baae.2008.1003.1008
Nilsson C (1985) Impact of ploughing on emergence of pollen beetle parasitoids after hibernation. J Appl Entomol 100:302–308
Nilsson C (2003) Parasitoids of pollen beetles. In: Alford D (ed) Biocontrol of oilseed rape insect pests. Blackwell, Oxford, pp 73–85
Nitzsche O, Ulber B (1998) Einfluß differenzierter Bodenbearbeitungssysteme nach Winterraps auf die Mortalität einiger Parasitoiden des Rapsglanzkäfers (Meligethes spp.). J Plant Diseas Protect 105:417–421
Östman Ö, Ekbom B, Bengtsson J (2001) Landscape heterogeneity and farming practice influence biological control. Basic Appl Ecol 2:365–371
Pulliam HR (1988) Sources, sinks, and population regulation. Am Natural 132:652–661
Roland J, Taylor PD (1997) Insect parasitoid species respond to forest structure at different spatial scales. Nature 386:710–713
Thies C, Tscharntke T (1999) Landscape structure and biological control in agroecosystems. Science 285:893–895
Thies C, Steffan-Dewenter I, Tscharntke T (2003) Effects of landscape context on herbivory and parasitism at different spatial scales. Oikos 101:18–25
Thies C, Steffan-Dewenter I, Tscharntke T (2008) Interannual landscape changes influence plant–herbivore–parasitoid interactions. Agric Ecosyst Environm 125:266–268
Tscharntke T (2000) Parasitoid populations in the agricultural landscape. In: Hochberg ME, Ives AR (eds) Parasitoid population biology. Princeton University Press, Princeton, pp 235–253
Tscharntke T, Kruess A (1999) Habitat fragmentation and biological control. In: Hawkins BA, Cornell HV (eds) Theoretical approaches to biological control. Cambridge University Press, London, pp 190–205
Ulber B (2003) Parasitoids of Ceutorhynchid stem weevils. In: Alford D (ed) Biocontrol of oilseed rape insect pests. Blackwell, Oxford, pp 87–95
Valantin-Morison M, Meynard J-M, Doréc T (2007) Effects of crop management and surrounding field environment on insect incidence in organic winter oilseed rape (Brassica napus L.). Crop Protect 26:1108–1120
Van Driesche RG, Bellows TS (1996) Biological control. Chapman & Hall, London
Wang B, Ferro DN, Hosmer DW (1997) Importance of plant size, distribution of egg masses, and weather conditions on egg parasitism of the European corn borer, Ostrinia nubilalis by Trichogramma ostriniae in sweet corn. Entomol Exp Appl 83:337–345
Williams IH (2004) Advances in insect pest management of oilseed rape in Europe. In: Horowitz AR, Ishaaya I (eds) Insect pest management—field and protected crops. Springer-Verlag, Heidelberg, pp 181–208
Williams IH, Free JB (1979) Compensation of oil-seed rape (Brassica napus L.) plants after damage to their buds and pods. J Agric Sci 92:53–59
Williams IH, Büchi R, Ulber B (2003) Sampling, trapping and rearing oilseed rape pests and their parasitoids. In: Alford D (ed) Biocontrol of oilseed rape insect pests. Blackwell, Oxford, pp 145–160
Williams IH, Frearson DJT, Barari H, McCartney HA (2007) First field evidence that parasitoids use upwind anemotaxis for host-habitat location. Entomol Exp Appl 123:299–307
With KA, Cadaret SJ, Davis C (1999) Movement responses to patch structure in experimental fractal landscapes. Ecology 80:1340–1353
Wratten SD, Van Emden HF (1995) Habitat management for enhanced activity of natural enemies of insect pests. In: Glen DM, Greaves MP, Anderson HM (eds) Ecology and integrated farming systems. Wiley, London, pp 117–145
Yee TW, Mitchell ND (1991) Generalized additive models in plant ecology. J Veg Sci 2:587–602
Zaller JG, Moser D, Drapela T, Schmöger C, Frank T (2008a) Effect of within-field and landscape factors on insect damage in winter oilseed rape. Agric Ecosyst Environm 123:233–238
Zaller JG, Moser D, Drapela T, Schmöger C, Frank T (2008b) Insect pests in winter oilseed rape affected by field and landscape characteristics. Basic Appl Ecol 9:682–690
Zar JH (1996) Biostatistical analysis, 3rd edn. Prentice-Hall, Englewood Cliffs
Acknowledgments
We are grateful to the farmers for participating in this research project, the staff of the research farm Groß-Enzersdorf for providing post-harvest equipment and to the Austrian Science Fund for supporting J.G.Z., D.M. and T.D. (grant no. P16972). The regional governments of Lower Austria and Burgenland provided maps and aerial photographs of the project area. Help by Norbert Schuller, Erhard Tesarik, Sandra Goldschall, Florian Heigl in the field and laboratory is gratefully acknowledged. We also thank Bernd Ulber and Michael Eikermann (Univ. Göttingen, Germany) for their advice on rearing and determining parasitoids. Two anonymous referees gave valuable comments on an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Torsten Meiners.
Rights and permissions
About this article
Cite this article
Zaller, J.G., Moser, D., Drapela, T. et al. Parasitism of stem weevils and pollen beetles in winter oilseed rape is differentially affected by crop management and landscape characteristics. BioControl 54, 505–514 (2009). https://doi.org/10.1007/s10526-009-9212-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-009-9212-2