
Abstract
Cold storage of parasitoids to be used in biological control programs is desirable but risky for the performance of the stored parental generation as well as for its offspring. We studied the performance of cold stored and unstored parasitoids after release at different temperature regimes in the laboratory at the level of two subsequent generations in Hyssopus pallidus (Askew) (Hymenoptera: Eulophidae). This gregarious ectoparasitoid is a candidate biocontrol agent of Cydia pomonella L. (Lepidoptera: Tortricidae) and Cydia molesta (Busck) (Lepidoptera: Tortricidae) larvae, two fruit pests of high economic significance in apple cultivation. Cold storage for 14 days at 4°C imposed to the pupal stage of the parasitoid did not reduce the parasitism capacity of the parental generation, nor did it alter the female biased sex ratio of the progeny. Remarkably, this short-term storage of the parental generation exhibited a significant and consistently positive effect on offspring weight throughout all ambient temperature regimes, resulting in an increased offspring weight. Furthermore, offspring number was only reduced after release at low ambient temperatures, but not at 25°C and 30°C. Irrespective of whether the parasitoids originated from the stored or unstored group, highest parasitism rate was achieved at temperatures above 20°C. In conclusion, this candidate biocontrol agent can be cold stored for short periods without any measurable quality loss after release at most except at low ambient temperatures. Our findings suggest that H. pallidus is a thermophilous parasitoid that will perform best when applied at warm ambient temperatures in fruit orchards.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The gregarious ectoparasitoid Hyssopus pallidus (Askew) (Hymenoptera: Eulophidae) is endemic to Europe (Askew 1964; Tschudi-Rein et al. 2004) and a candidate biocontrol agent of fruit moths of the genus Cydia in orchards (Mattiacci et al. 1999; Häckermann et al. 2007). This parasitoid is relatively specialized (Zaviezo and Mills 2000), with the codling moth Cydia pomonella (L.) and the Oriental fruit moth C. molesta (Busck) being its only known hosts in Europe, as first (Askew 1964) and recent (Tschudi-Rein et al. 2004) field samplings document. Similar to other Eulophidae (Brown 1996), this species has a rapid preimaginal development, a high lifetime fecundity and a female biased sex ratio (Zaviezo and Mills 1999; Tschudi-Rein and Dorn 2001; Häckermann et al. 2007), rendering it particularly suitable for mass production. Female parasitoids use plant as well as host derived cues to locate their host (Mattiacci et al. 2000). Once they have found an apple infested by a Cydia caterpillar, they enter the fruit either through the gallery made by the caterpillar or through the calyx and parasitize the host in the center of the fruit (Mattiacci et al. 1999).
In the past decades, many novel strategies, such as mating disruption (Judd and Gardiner 2005), the sterile insect technique and microbial control (Falcon and Huber 1991; Bloem et al. 2006) have been developed to control Cydia species, but their success depends on low population pressure (e.g., Cardé and Minks 1995). These strategies are not only environmentally friendly control methods that protect the environment and human health, but they are also harmless to beneficial insects. However, despite these new strategies, insecticides remain the principal method of control for these pests (Audemard et al. 1992; Ohlendorf 1999). In apple cultivations, the codling moth is the most devastating pest (Barns 1991; Dorn et al. 1999), while the oriental fruit moth gained importance as an additional pest in recent years. Originally known as a key pest of peach, this species has lately been appearing to an increasing degree as a pest insect in apple orchards in various countries (Rothschild and Vickers 1991; Pollini and Bariselli 1993; Hughes et al. 2004). As a consequence, apple growers are confronted with the need to control both tortricid pests in their orchards, and therefore improved IPM (integrated pest management) strategies are required. Since H. pallidus successfully attacks both host species (Häckermann et al. 2007) and thus holds the potential to reduce their population pressure, it could become a useful component of IPM systems designed to keep both Cydia populations below the economic threshold level (Hein and Dorn 2008).
Short-term storage of parasitoids that are to be used in biological control programs is highly desirable, but often difficult to implement. To better supply the market in times when demands are high, a certain storage period at a suitable stage would be highly useful. Despite the fact that storage of natural enemies at low temperature has widely been studied, its practical applications are limited for several reasons: (1) storage might lead to a high loss in emergence rates (Levie et al. 2005; Lopez and Botto 2005; Pandey and Johnson 2005); (2) parasitism rate can be reduced due to cold storage (Pitcher et al. 2002; Ozder 2004; Levie et al. 2005; Lopez and Botto 2005; Chong and Oetting 2006; Luczynski et al. 2007); or (3) other life history traits such as longevity and sex ratio might be changed (Uckan 2001; Torres et al. 2002; Foerster et al. 2004), although such negative influences are not always observed (Leopold 1998). The development of a suitable storage strategy should aim at maintaining natural characteristics of the beneficial insect throughout the storage process to guarantee a high quality biological control agent (van Lenteren and Tommasini 2003). However, such cold storage conditions are still unknown for H. pallidus, which likely overwinters at the pupal stage (K. Tschudi-Rein, unpublished pilot study) that thus might be a suitable stage for storage considering the typical relationship between diapause and cold hardiness (Denlinger 1991).
Furthermore, the choice of suitable release temperatures can have a strong impact on parasitoid performance in the field (Godfray 1994). Despite the interest in H. pallidus for the control of Cydia pests in orchards (Hein and Dorn 2008), no information on optimal temperature conditions for its parasitism and fitness is available so far.
Our objective was to investigate the performance of cold stored and unstored parasitoids under different ambient temperature regimes. Specifically, we assessed (1) the influence of a short cold storage period on biological and behavioural characteristics, in particular the parasitism rate and the reproductive capacity of this species, (2) two key quality traits of the offspring, namely the pupal weight and the sex ratio, and (3) the effects of ambient temperature on parasitoid performance. This research should help to judge the potential to store this candidate biocontrol agent for short periods without quality loss, and to define the optimal ambient temperature conditions for successful field application in fruit orchards.
Materials and methods
Insect culture
The initial colony of Hyssopus pallidus originating from France was provided in 1996 by T. Unruh (USDA-ARS Research Laboratory, Wapato Washington, USA) (Tschudi-Rein and Dorn 2001), and genetically supplemented by the introduction of field-collected adult wasps from Switzerland. The host species C. pomonella was reared on wheat-germ-based artificial diet (Huber et al. 1972) complemented with sterile beech wood chips (6 mm). Adult moths were kept in wax paper lined containers for one day for ovipositon. Eggs were thereafter kept at 25°C for optimal embryonic development. Shortly before the hatching of the first instar larvae, the egg sheets were placed with the eggs facing the artificial medium. Upon eclosion, egg sheets were removed and boxes were closed and kept at 25 ± 2°C, 60 ± 10% RH and 16L:8D until all larvae had reached the last larval stage. Corrugated cardboard was added to the box to facilitate pupation.
Several 5th instar larval hosts at a time were offered to four to seven days old mated female parasitoids at a ratio of one parasitoid per host in a square plastic box (10 cm × 10 cm × 8 cm), which had two meshed net covered holes to ensure for sufficient ventilation. Since plant cues offered during ontogenesis of the parasitoid are of crucial importance for the behavioral quality of the emerged wasps (Gandolfi et al. 2003b) small pieces of fresh apple (Gandolfi et al. 2003a) were added. A droplet of undiluted honey served as food source for the female parasitoids. The wasps were removed from the box one week later. Parasitized larvae were kept under standardized laboratory conditions at a temperature of 24 ± 2°C, RH of 60 ± 10% and a regime of 16L:8D until the emergence of the parasitoid progeny 14–17 days later. Upon emergence, progeny was removed from the box and transferred to a Plexiglas cage (25 cm × 25 cm × 25 cm), and fed with honey and water. C. molesta, that were used in the experiments, were reared in the same way as described above for C. pomonella. Irrespective of the host the parasitoid had developed on, the performance of emerged adults and their offspring was similar, rendering either host species suitable for rearing (Häckermann et al. 2007).
Effect of storage regime and ambient temperature on parasitism success, offspring number and fitness
This experiment simultaneously tested the effects and interactions of storage and four different ambient temperature regimes on the parasitism behavior and the parasitism success of female H. pallidus wasps. To evaluate the influence of short-term cold storage on the success of the parasitoid, two groups of females were tested: (1) an unstored control group of females, kept in the rearing climate chamber of 25 ± 2°C, 60 ± 10% RH and 16L:8D and (2) a cold stored group of females that had been reared exactly as the control group, except that it was transferred at the pupal stage to a standard cold room of 4 ± 2°C, 80 ± 5% RH, 0L:24D, stored there for 2 weeks, and then brought back to the rearing chamber at 25 ± 2°C for emergence. Cool room temperature of 4°C, i.e., slightly above the freezing point, is common in many laboratories and biocontrol companies (J. Häckermann pers. obs.), and a 2 weeks storage period is considered to reflect the typical period for phenological delays often observed in the apple crop in spring. Individuals from both groups were weighed as mature pupae, in the case of the cold stored group immediately before storage, using a precision balance (Mettler MT 5 Delta range; 2 ± μg). To ensure that they were weighted at the same age, weighting was carried out at the black pupal stage that immediately (<1 day, J. Häckermann pers.obs.) precedes adult emergence. After emergence, parasitoids from both groups were kept in the rearing chamber at 25 ± 2°C, 60 ± 10% RH and 16L:8D and fed with undiluted honey and water for 4–7 days prior exposure to different ambient temperature regimes. All females used in the experiment had developed on C. pomonella hosts and were standardized by size (0.52–0.56 mg in pupal weight) for the experiment to ensure a minimal size range.
For assessment of ambient temperature on parasitism efficiency, a cohort of 10 individuals was set-up for each of the two storage treatments and for four ambient temperature regimes (15, 20, 25 or 30 ± 2°C all at a RH of 70 ± 5% and 16L:8D), resulting in a total of eight cohorts with 10 individuals respectively per replication. A total of 10 replicates per storage × temperature combination were tested. In the experimental setup, one standardized last instar (16–25 mg) C. molesta larva was placed into a plastic Petri dish (diameter 5.5 cm, height 1.3 cm) together with a female wasp of either of the storage treatments. A droplet of honey served as food source.
Parasitism success was assessed 24, 72 and 120 h after the onset of the experiment. Parasitoids were considered successful if eggs were deposited on the paralyzed host body. The presence of eggs was determined by examining the host’s body through a stereomicroscope without touching the developing brood. Where eggs were present, insects were kept at 25 ± 1°C and RH 65 ± 5% until completion of offspring development. The remaining test individuals of the cohort were taken back to the climate chamber with the respective temperature regime and checked again 72 and 120 h after the onset of the experiment.
Upon successful larval development of the parasitoid offspring, the number of progeny surviving to adulthood was estimated by counting the number of pupae that had successfully developed under the different temperature regimes. Furthermore the number of males per clutch was counted. To determine the influence of storage type of the parental generation and temperature regime during oviposition on the fitness of the F1 generation, all pupae that had developed on one host were weighed. The wasp’s size is directly correlated with fitness resulting in larger females being fitter than smaller ones (Godfray 1994). For the subsequent analysis, the average offspring size was calculated for each clutch, and used as basis for all further analyses.
Statistical analysis
Percentage of parasitism after 24, 72 and 120 h was arcsine square root transformed and analyzed in a two way ANOVA with storage regime, temperature and their interaction as independent variables. Bonferroni’s post hoc multiple comparison tests were used when treatments were significantly different. To evaluate the influence of temperature on the number of offspring produced per host larva, Kruskal–Wallis tests were performed. Differences within each temperature and between the cold stored and the unstored group was evaluated using a Mann–Whitney U-test.
To evaluate the influence of storage regime and temperature on the fitness of the F1 generation, offspring weight per clutch was determined. The data were square root transformed to approach normality and homogeneity of variance. An ANOVA procedure was used to determine whether cold storage of the F0 generation and the different temperatures during oviposition had an influence on offspring fitness.
Results
Effect of storage regime and ambient temperature on parasitism success
Cold storage of the parental generation at the pupal stage did not have any negative influence on the performance of the adult wasps (Table 1). However, increasing ambient temperature during oviposition significantly increased the parasitism success of H. pallidus. Cumulative parasitism rate within each temperature range was compared after 24, 72 and 120 h of exposure (see Fig. 1). At an ambient temperatures of 15°C, an exposure period of 24 h resulted in a parasitism rate of 22 ± 21% (mean ± SD) in the cold stored group and 26 ± 25% in the unstored group. Longer exposure resulted in an increased parasitism rate in both groups with a maximum reached after 120 h (cold stored 41 ± 21%, untreated 57 ± 21%). Larvae that were not parasitized after 120 hours had either pupated or died without any eggs present on the host’s body and were not considered in the analysis.

Mean percentage of parasitism of H. pallidus in two groups: half of the parasitoids had been stored at 4°C for 14 days at the pupal stage, the other half had not been stored (unstored control group). Parasitism was assessed at different temperatures (15, 20, 25 and 30°C) and different hours (24, 72 and 120 h) of adult exposure to C. molesta hosts. N = 10 for each treatment at every temperature. The column height indicates the cumulative parasitism rate after 24, 72 and 120 h of exposure. White column parts indicate the parasitism rate from 0 to 24 h, gray column parts the increase in parasitism between 25 and 72 h and black column parts the increase in parasitism rate between 73 and 120 h of exposure. Letters indicate differences within each storage group. Bonferroni’s post hoc test
At an ambient temperature of 20°C, 24 h of exposure resulted in a significantly higher parasitism rate than at 15°C (Bonferroni, P < 0.001), resulting in 63 ± 15% in the cold stored and 63 ± 21% in the unstored group. Seventy-two hours of exposure time yielded significantly higher parasitism rates than at 15°C in both groups (cold stored 85 ± 9%, untreated 81 ± 10%) (Bonferroni, P < 0.001). A further prolongation of the exposure time up to 120 h increased the parasitism rate by only 2% (cold stored 93 ± 8%, untreated 86 ± 11%).
At an ambient temperature of 25°C, 24 h of exposure resulted in a significantly higher parasitism rate than at 15 and 20°C in both the cold stored (76 ± 15%) and the unstored group (72 ± 21%). After 72 h exposure, most hosts had been parasitized (cold stored 90 ± 9%, untreated 87 ± 11%) but with longer exposure, parasitism rate was no longer significantly higher than at 20°C. One hundred and twenty hours of exposure again resulted in the highest parasitism rate (cold stored 91 ± 10%, untreated 89 ± 11%).
At a high ambient temperature of 30°C, 24 h of exposure led to a high parasitism rate in the cold stored (84 ± 12%) as well as in the unstored group (83 ± 21%). By 72 h of exposure most hosts were parasitized (cold stored 91 ± 9%, untreated 90 ± 12%), although this result was not significantly different than at 25°C (Fig. 1). A further increased exposure time (120 h) did not result in an increase in parasitism (cold stored 91 ± 9%, untreated 91 ± 11%).
Effect of storage regime and ambient temperature on offspring number, fitness and sex ratio
At low ambient temperatures, i.e. at 15°C (z = −2.4, P = 0.068) and at 20°C (z = −3.2, P = 0.004) cold storage resulted in a lower number of progeny produced per female. At more elevated ambient temperatures (25°C, z = −0.66, P > 0.5; 30°C, z = −1.36, P > 0.1), the number of offspring was similar, irrespective of whether the parental generation had been cold stored or not (Fig. 2).

Mean number ± SE of offspring produced per host larva by females ovipositing at different ambient temperatures (15, 20, 25 and 30°C). The cold stored group had been exposed to low temperatures (4°C) for 14 days, the unstored group had constantly been kept at 25°C. Letters indicate differences within each storage group. Kruskal–Wallis test
In the cold stored group, a positive effect of increasing ambient temperature on offspring number could be observed (χ2 = 19.09, df = 3, P < 0.001). Significantly more offspring were produced at 25°C (z = −3.8, P < 0.001) and 30°C (12.9 ± 5.9) (z = −2.6, P < 0.036) than at 15°C (10.3 ± 5.2). Equally, at 25°C an average of 14.1 ± 5.8 pupae were produced per clutch, which was significantly more than at 20°C (11.1 ± 6.6) (Fig. 2). In the group that had never been cold stored, temperature during oviposition did not affect the number of offspring that were produced per host larva (Fig. 2).
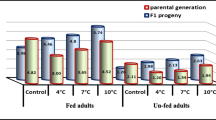
Significantly (F = 10.06, df = 1, O = 0.01) larger pupae were produced when the parental generation had been stored as pupae at 4°C for 14 days (on average 0.36 mg ± 0.07) than if they had never experienced a cold period (on average 0.32 mg ± 0.07) (Fig. 3). By contrast, neither temperature during the time of oviposition (F = 0.68, df = 3, P > 0.5) nor the experimental unity (F = 0.306, df = 9, P > 0.95) had any influence on the size of the F1 generation.

Offspring weight per clutch of females that had been stored at 4°C for 14 days (black) and females that were never exposed to low temperatures (unstored control) (white). Temperature ranges on the x-axis indicate the mean ambient temperature present during oviposition. N = 100 for each treatment/temperature combination
Neither the storage type (χ2 = 0.35, df = 1, P = 0.71) nor the temperature during oviposition (χ2 = 2.21, df = 3, P = 0.53) had an effect on the number of males produced per clutch. On average, 1.04 ± 0.5 males were produced on each host larva in the cold stored group, whereas an average of 1.11 ± 0.54 males emerged from the control group (Table 2).
Discussion
The performance of cold stored and unstored H. pallidus parasitoids was elucidated after release under different temperature regimes. Our results provide empirical evidence that this candidate biocontrol agent of the codling moth and the Oriental fruit moth can be stored for 2 weeks at 4°C without any detrimental effect on the parasitism capacity. At all ambient temperatures tested, parasitoids that had been cold stored during their pupal stage showed the same performance in parasitizing host larvae as the control group that had never experienced a period of low temperature. This result indicates that the pupal stage of this parasitoid is well suitable for short-term storage in this species. Whether longer storage results in a loss in quality as shown for different parasitoid species (Rundel et al. 2004) is yet unknown. Typically, storage only lasts for a restricted number of weeks, but even then a reduction in fitness is the rule (Posthuma-Doodeman et al. 1996). For example, fecundity can strongly be negatively affected by cold storage (Bayram et al. 2005; Pandey and Johnson 2005). Remarkably, parasitism rate of female H. pallidus was not altered by a two weeks cold storage period at the pupal stage.
Hyssopus pallidus was found to produce the same number of offspring per clutch at all temperature ranges if the parental generation had not been exposed to low temperatures during its immature phase. However, if the parasitoid had been stored at 4°C for 14 days, wasps produced the largest clutches at higher temperatures, with the least number of offspring per host larva being laid at 15°C. Although cold storage might have a negative effect on the number of offspring at low temperatures, this effect disappears if ambient temperatures during oviposition are above 20°C. Hence, the reproductive capacity in this species is not impeded after the tested cold storage when adults are released into a warm environment. Further, this finding indicates that H. pallidus may be most efficient as biological control agent under warm climatic conditions.
Further evaluations focused on the F1 generation originating from the cold stored and unstored parental generation. Interestingly, larger offspring individuals developed when the parental generation had experienced a period of low temperature during its pupal stage, whereas ambient temperature during oviposition did not have an influence on offspring weight. Weight as a fitness parameter is important because adult female weight may influence fitness by affecting the searching efficiency, longevity and fecundity of the wasps (Godfray 1994). The maintenance of all these mentioned parameters on a high level is of crucial importance for the quality of commercially produced natural enemies (van Lenteren and Tommasini 2003). In H. pallidus, it seems that cold storage elicits a reaction either in the female wasps or in its progeny, leading to an increased offspring weight at high temperatures. This finding documents that this parasitoid does not suffer from short periods of cold storage, but on the contrary even potentially benefits with larger offspring.
Further, the sex ratio of the F1 progeny was not altered when H. pallidus was cold stored as pupa. The strongly female biased sex ratio in this species (Zaviezo and Mills 1999) was maintained. This quality trait in the offspring is particularly important for parasitoid mass production in biological control programs, as only female wasps can attack hosts (Godfray 1994).
Many studies on cold storage of hymenopteran parasitoids have focused on endoparasitoids, which are stored within their host egg or host larva (Leopold 1998; Bayram et al. 2005; Pandey and Johnson 2005), whereas ectoparasitoids such as H. pallidus are much more exposed to abiotic factors like low temperatures. However, even if parasitoids are protected within host tissue they often experience detrimental effects due to cold storage (Ozder 2004; Pandey and Johnson 2005). Our results demonstrate that this ectoparasitoid does not suffer from adverse abiotic factors even if the pupae are not protected during the storage period. Furthermore the pupal stage seems to be well suitable for short-term storage in this species.
Temperature present during oviposition had a marked effect on parasitism. The highest parasitism rate was consistently achieved in both the cold stored and the unstored group, when ambient temperatures during oviposition were around 25°C or higher. Though the exposure time was not the main factor influencing the parasitism success, a clear increase in number of parasitized larvae could be observed with increasing exposure time, especially at low temperatures. This is in agreement with studies on other parasitoids that showed that temperatures around 15°C often have an inhibitory effect on the wasp’s parasitism capacity (Torres et al. 2002; Foerster and Butnariu 2004). Furthermore, Pilkington and Hoddle (2006) described that already moderate changes in climatic conditions, such as changes in temperature, can adversely affect parasitoid populations in a way that their impact on pest populations is drastically reduced. Our results indicate that for H. pallidus ambient temperature is an important abiotic factor that influences the success of the released wasps. Longer exposure time can partially compensate the adverse effect of low ambient temperatures, but temperature remains the key factor for successful parasitism activities. We therefore conclude that H. pallidus is a thermophilous parasitoid with the best performance at a mean ambient temperature of 25°C and above and that temperature is most likely the limiting abiotic factor for the parasitoid’s reproductive capacity in the field.
Based on our laboratory experiments, H. pallidus can be cold stored during its pupal stage for up to 14 days without any negative effect on its parasitism rate and reproductive capacity. On the contrary, cold storage can even have a positive effect on its progeny, resulting in larger offspring. Our results further support the hypothesis that the pupal stage might be the overwintering stage of this parasitoid, although further investigations are needed for confirmation. Warm temperatures for field releases in fruit orchards are expected to support parasitism success.
References
Askew RR (1964) On the biology and the taxonomy of some European species of the genus Elachertus spinola (Hymenoptera: Eulophidae). Bull Entomol Res 55:53–58
Audemard H, Baudry O, Bergere D, Breniaux D, Gendrier JP (1992) Integrated plant protection in apple orchards of the 21st century. Acta Phytopathol Entomol Hung 27:39–44
Barns MM (1991) Codling moth occurrence, host race formation and damage. In: van der Geest LPS, Evenhuis HH (eds) World crop pests: tortricid pests. Elsevier Science Publishers B.V., Amsterdam, pp 313–327
Bayram A, Ozcan H, Kornosor S (2005) Effect of cold storage on the performance of Telenomus busseolae Gahan (Hymenoptera: Scelionidae), an egg parasitoid of Sesamia nonagrioides (Lefebvre) (Lepidoptera: Noctuidae). Biol Control 35:68–77
Bloem S, Carpenter JE, Dorn S (2006) Mobility of mass-reared diapaused and nondiapaused Cydia pomonella (Lepidoptera: Tortricidae): effect of different constant temperatures and lengths of cold storage. J Econ Entomol 99:707–713
Brown JJ (1996) The compatibility of tebufenozide with a laboratory lepidopteran host hymenopteran parasitoid population. Biol Control 6:96–104
Cardé RT, Minks AK (1995) Control of moth pests by mating disruptions: success and constraints. Annu Rev Entomol 40:559–585
Chong JH, Oetting RD (2006) Influence of temperature, nourishment, and storage period on the longevity and fecundity of the mealybug parasitoid, Anagyrus sp nov nr. sinope Noyes and Menezes (Hymenoptera: Encyrtidae). Environ Entomol 35:1198–1207
Denlinger DL (1991) Relationship between cold hardiness and diapause. In: Lee EL, Denlinger DL (eds) Insects at low temperature. Chapman and Hall, New York and London, pp 174–198
Dorn S, Schumacher P, Abivardi C, Meyhöfer R (1999) Global and regional pest insects and their antagonists in orchards: spatial dynamics. Agricult Ecosys Environ 73:111–118
Falcon LA, Huber J (1991) Biological control of the codling moth. In: van der Geest LPS, Evenhuis HH (eds) World crop pest: tortricid pests. Elsevier Science Publishers B.V., Amsterdam, pp 355–369
Foerster LA, Butnariu AR (2004) Development, reproduction, and longevity of Telenomus cyamophylax, egg parasitoid of the velvetbean caterpillar Anticarsia gemmatalis, in relation to temperature. Biol Control 29:1–4
Foerster LA, Doetzer AK, de Castro LCF (2004) Emergence, longevity and fecundity of Trissolcus basalis and Telenomus podisi after cold storage in the pupal stage. Pasq Agro Brasil 39:841–845
Gandolfi M, Mattiacci L, Dorn S (2003a) Mechanisms of behavioral alterations of parasitoids reared in artificial systems. J Chem Ecol 29:1871–1887
Gandolfi M, Mattiacci L, Dorn S (2003b) Preimaginal learning determines adult response to chemical stimuli in a parasitic wasp. Proc R Soc Lond (Biol) 270:2623–2629
Godfray HCJ (1994) Parasitoids: behavioral and evolutionary ecology. Princeton University Press, Princeton
Häckermann J, Rott AS, Dorn S (2007) How two different host species influence the performance of a gregarious parasitoid: host size is not equal to host quality. J Anim Ecol 76:376–383
Hein S, Dorn S (2008) The parasitoid of a fruit moth caterpillar utilizes fruit components as nutrient source to increase its longevity and fertility. Biol Control DOI:10.1016/j.biocontrol.2007.10.013
Huber J, Benz G, Schmid K (1972) Breeding method and semisynthetic culture media for Laspeyresia pomonella (Carpocapsa pomonella). Experientia 28:1260–1261
Hughes J, Hern A, Dorn S (2004) Preimaginal environment influences adult flight in Cydia molesta (Lepidoptera: Tortricidae). Env Entomol 33:1155–1162
Judd GJR, Gardiner MGT (2005) Towards eradication of codling moth in British Columbia by complimentary actions of mating disruption, tree banding and sterile insect technique: five-year study in organic orchards. Crop Prot 24:718–733
Leopold RA (1998) Cold storage of insects for integrated pest management. In: Hallman GJ, Denlinger DL (eds) Temperature sensitivity in insects and application in integrated pest management. Westview press, Boulder, pp 235–267
Levie A, Vernon P, Hance T (2005) Consequences of acclimation on survival and reproductive capacities of cold-stored mummies of Aphidius rhopalosiphi (Hymenoptera: Aphidiinae). J Econ Entomol 98:704–708
Lopez SN, Botto E (2005) Effect of cold storage on some biological parameters of Eretmocerus corni and Encarsia formosa (Hymenoptera: Aphelinidae). Biol Control 33:123–130
Luczynski A, Nyrop JP, Shi A (2007) Influence of cold storage on pupal development and mortality during storage and on post-storage performance of Encarsia formosa and Eretmocerus eremicus (Hymenoptera: Aphelinidae). Biol Control 40:107–117
Mattiacci L, Hütter E, Dorn S (1999) Host location of Hyssopus pallidus, a larval parasitoid of the codling moth, Cydia pomonella. Biol Control 15:241–251
Mattiacci L, Hütter E, Schoch D, Scascighini N, Dorn S (2000) Plant-odour mediates parasitoid host handling and oviposition in an endophytic tritrophic system. Chem Ecol 10:185–192
Ohlendorf BLP (1999) Integrated pest management for apples and pears. Division of Agricultural Sciences University of California, Oakland, CA
Ozder N (2004) Effect of different cold storage periods on parasitization performance of Trichogramma cacoeciae (Hymenoptera, Trichogrammatidae) on eggs of Ephestia kuehniella (Lepidoptera, Pyralidae). Biocontrol Sci Technol 14:441–447
Pandey RR, Johnson MW (2005) Effects of cool storage on Anagyrus ananatis Gahan (Hymenoptera: Encyrtidae). Biol Control 35:9–16
Pilkington LJ, Hoddle MS (2006) Reproductive and development biology of Gonatocerus ashmeadi (Hymenoptera: Mymaridae), an egg parasitoid of Homalodisca coagulata (Hemiptera: Cicadellidae). Biol Control 37:266–275
Pitcher SA, Hoffmann MP, Gardner J, Wright MG, Kuhar TP (2002) Cold storage of Trichogramma ostriniae reared on Sitotroga cerealella eggs. BioControl 35:9–16
Pollini A, Bariselli M (1993) Cydia molesta pest on the increase and defence of pome fruits. Inform Agrario 49:19–21
Posthuma-Doodeman CJAM, van Lenteren JC, Sebestyen I, Ilovai Z (1996) Short-range flight test for quality control of Encarsia formosa. Proc Exper Appl Entomol 7:153–158
Rothschild GHL, Vickers RA (1991) Biology, ecology and control of the oriental fruit moth. In: van der Geest LPS, Evenhuis HH (eds) World crop pest: tortricid pests. Elsevier Science Publishers B.V., Amsterdam, pp 389–412
Rundel BJ, Thomson LJ, Hoffmann AA (2004) Effects of cold storage on field and laboratory performances of Trichogramma carverae (Hymenoptera: Trichogrammatidae) and the response of three Trichogramma spp. (T.carverae, T.nr.brassicae and T. funiculatum). J Econ Entomol 97:213–221
Torres JB, Musolin DL, Zanuncio JC (2002) Thermal requirements and parasitism capacity of Trissolcus brochymenae (Ashmead) (Hymenoptera: Scelionidae) under constant and fluctuating temperatures, and assessment of development in field conditions. Biocontrol Sci Technol 12:583–593
Tschudi-Rein K, Brand N, Kührt U, Dorn S (2004) First record of Hyssopus pallidus (Askew, 1964) for Switzerland (Hymenoptera: Eulophidae). Rev Suisse Zool 111:671
Tschudi-Rein K, Dorn S (2001) Reproduction and immature development of Hyssopus pallidus (Hymenoptera: Eulophidae), an ectoparasitoid of the codling moth. Eur J Entomol 98:41–45
Uckan F (2001) The effects of cold storage on the adult longevity, fecundity and sex ratio of Apanteles galleriae Wilkinson (Hym.: Braconidae). Turk J Entomol 98:41–45
van Lenteren JC, Tommasini MG (2003) Mass productuion, storage, shipment and release of natural enemies. In: van Lenteren JC (ed) Quality control and production of biological control agens: theory and testing procedure. CABI, Cambridge, pp 181–189
Zaviezo T, Mills N (1999) Aspects of the biology of Hyssopus pallidus (Hymenoptera: Eulophidae), a parasitoid of the codling moth (Lepidoptera: Olethreutidae). Environ Entomol 28:748–754
Zaviezo T, Mills N (2000) Factors influencing the evolution of clutch size in a gregarious insect parasitoid. J Anim Ecol 69:1047–1057
Acknowledgements
We thank Dr. Silke Hein and two anonymous reviewers for useful comments on the manuscript. This research, proposed by Silvia Dorn, was supported by SafeCrop Centre, funded by Fondo per la Ricerca, Autonomous Province of Trento.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Häckermann, J., Rott, A.S., Tschudi-Rein, K. et al. Cold stored ectoparasitoid of Cydia fruit moths released under different temperature regimes. BioControl 53, 857–867 (2008). https://doi.org/10.1007/s10526-007-9147-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-007-9147-4