
Abstract
Naringenin, the predominant bioflavonoid found in grapefruit and tomato has diverse bioactive properties that encompass anti-carcinogenic, anti-inflammatory, anti-atherogenic, anti-estrogenic, anti-hyperlipidemic and anti-hyperglycemic characteristics. Naringenin has not been explored for its pro-longevity traits in fruit flies. Therefore, the current study explores its influence on longevity, fecundity, feeding rate, larval development, resistance to starvation stress and body weight in male and female wild-type Drosophila melanogaster Canton-S flies. Flies were fed with normal and high fat diets respectively. The results implied hormetic effects of naringenin on longevity and development in flies. In flies fed with standard and high fat diets, lower concentrations of naringenin (200 and 400 µM) augmented mean lifespan while higher concentrations (600 and 800 µM) were consistently lethal. However, enhanced longevity seen at 400 µM of naringenin was at the expense of reduced fecundity and food intake in flies. Larvae reared on standard diet having 200 µM of naringenin exhibited elevated pupation and emergence as flies. Eclosion time was hastened in larvae reared on standard diet having 200 µM of naringenin. Female flies fed with a standard diet having 200 and 400 µM of naringenin were more resistant to starvation stress. Reduction in body weight was observed in male and female flies fed with a high fat diet supplemented with 200 and 400 µM of naringenin respectively. Collectively, the results elucidated a context- and dose-dependent hormetic efficacy of naringenin that varied with gender, diet and stage of lifecycle in flies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Aging is a progressive multifactorial metabolic phenomenon associated with a plethora of age-related complications covering neurodegenerative diseases, myopathies, inflammation, cancer, diabetes, and obesity etc. (Fontana et al. 2010). Past few decades have witnessed a surge in aging research involving pharmaceutical, botanical or dietary interventions. Emerging evidences have implicated the role of phytocompounds in modulating longevity (Bao and Fenwick 2004; Bass et al. 2007; Joseph et al. 2009, 2007; Lee et al. 2010; Wang et al. 2013). Naringenin (4′,5,7-trihyroxyflavanone) is an aglycone of naringin and the predominant flavonoid found in grapefruit (Citrus paradisi) (up to 10 % of dry weight) and tomato (Lycopersicum esculentum) (Davies et al. 1981; Kawaii et al. 1999; Semalty et al. 2010). In citrus fruits, naringenin predominantly exists as naringenin-7-neohesperoside and naringenin-7-rutinoside. Previous studies have reported anti-inflammatory, anti-carcinogenic and anti-estrogenic characteristics of naringenin (Benavente-García et al. 1997; Guthrie and Carroll 1998; Middleton and Kandaswami 1994; Ruh et al. 1995). Naringenin has been shown to possess anti-hyperlipidemic and anti-hyperglycemic properties (Mulvihill et al. 2009; Ortiz‐Andrade et al. 2008). Additionally, naringenin activates phosphatidylinositol 3-kinase, independent of insulin receptor substrate-1 phosphorylation (Borradaile et al. 2003). Furthermore, naringenin reduced malonaldehyde production by inhibiting the cytochrome P450 enzymes (Ueng et al. 1999). However, caution is warranted before un-mindful and large-scale consumption of naringenin. Intestinal drug metabolism and secretory transport systems undergo interference by constituents of grape-fruit juice, specifically naringenin (Wagner et al. 2001). Recent studies have highlighted the capability of naringenin to undergo interactions with drugs such as lovastatin and enalapril via esterase inhibition (Li et al. 2007). Other prominent drugs which interact with naringenin are calcium channel blockers (amlodopine, nisoldipine, verapamil etc.), statins (atorvastatin, cervastatin, fluvastatin, paravastatin, etc.), aphrodisiacs (viagra/sildenafil), valium (diazepam), buspirone (buspar), quinidine, quinine, prednisone, etc. (Mozayani and Raymon 2011; Wunderlich and Piemonte 2012).
Bulk of the current world population is affected by obesity as a result of uncontrolled feeding habits and unhealthy lifestyle. Obesity is a known hurdle to successful and healthy aging since it paves way for various diseases which include type 2 diabetes, neurodegenerative disorders, strokes, etc. (Chibnall 2010). A study dating back to 1955 had shown that mice fed with a high fat diet from as young as 1 month of age significantly shortened lifespan (Silberberg and Silberberg 1955). Another study has highlighted the decreased survival of mice when fed with a high fat diet which was independent of body weight (Muller et al. 2013). Since naringenin has not yet been inspected for its impact on longevity in flies, the objective of the current study was to assess the effects of naringenin on survival and development in Drosophila melanogaster fed with normal and high fat diets. Also, our aim was to assess the concentration range within which naringenin exerted the beneficial effects without hampering normal physiological processes. The various parameters evaluated were longevity, fecundity, feeding rate, percentage pupation, percentage eclosion, eclosion time, resistance to starvation and body weight. Since naringenin is a novel compound used in longevity research, both positive and negative effects of naringenin were analysed over a wide range of concentrations in male and female flies fed with normal and high fat diets respectively.
Materials and methods
Fly husbandry and diet preparation
Experiments were performed with wild type Drosophila melanogaster Canton-S (CS) flies reared at temperature 25 ± 1 °C on 12:12 h light: dark cycle under standard fly rearing conditions. Prior to the assays, flies were bulk reared in 300 ml polypropylene bottles having 30 ml of standard cornmeal diet (9 % cornmeal, 4 % dextrose, 2 % sucrose, 3 % yeast extract; all concentrations given in w/v). Each bottle housed 50 flies to avoid overcrowding. Approximately 50 eggs were carefully collected over 24 h and transferred to new bottles. Eggs were allowed to hatch followed by larval pupation. For all assays, newly eclosed flies were collected over 48 h and allowed to mate for another 48 h. Flies were then segregated according to their sex and transferred to either the standard sugar-yeast (SY) diet (10 % sucrose, 10 % yeast extract; all concentrations given in w/v) or high fat diet (HFD) (10 % sucrose, 10 % yeast extract, 2 % palmitic acid; all concentrations given in w/v) with or without naringenin (Sigma-Aldrich, India, N5893). All diets had 1 % agar (w/v), 0.3 % propionic acid (v/v) and 3 % methylparaben (v/v). For naringenin supplemented diets, naringenin was dissolved in absolute ethanol and incorporated into standard diet and HFD at final concentrations of 50, 100, 200, 400, 600 and 800 µM by thorough mixing. Standard and HFD control diets contained only absolute ethanol. For developmental assays, larvae were reared in 300 ml bottles each with 30 ml of respective diets with or without naringenin. For remaining assays, flies were reared in 50 ml polypropylene vials each with 7 ml of respective diets with or without naringenin.
Longevity assay
To measure fly longevity, single-sex flies were segregated into vials containing HFD or standard diet with or without naringenin. Flies were transferred to vials with fresh food for every 2 days, during which the number of dead flies were recorded. Assay was continued till all flies were dead. For each diet and sex, 10 replicate vials were set up with 30 flies in each vial.
Measurement of fly fecundity
Newly eclosed flies were kept as 2 males and 2 virgin females on HFD and standard diets with or without naringenin. Flies were transferred to fresh diets every day and number of eggs laid was counted every day for 15 days. For each diet, 10 replicates were set up with two males and two females in each replicate.
Measurement of fly feeding rate
Food intake was measured using the capillary feeder method (CAFE) with slight modifications (William et al. 2007). Single-sex flies were allowed to feed on HFD or standard diets with or without naringenin for 15 days. Flies were transferred to capillary feeding chambers housing 2 flies per chamber. The feeding capillaries were filled with agar-free liquid HFD or standard diets with or without naringenin. Two capillary chambers were set up without flies to measure evaporation of food. The volume of food intake was recorded once every 24 h for seven consecutive days. Average food intake for 7 days per fly was then calculated. For each diet and sex, 10 replicate chambers were set up with 2 flies in each chamber.
Measurement of larval development
Freshly hatched, age matched 1st instar larvae were collected from bulk fly stock cultures and carefully transferred to bottles with HFD or standard diet with or without naringenin. Larvae were allowed to rear and pupate. Each pupa was numerically marked on the side of bottles and observed further. Time required by individual larvae to pupate and eclose as a fly was monitored and noted down. Percentage of larvae that had pupated or flies that had eclosed was calculated. For each diet, 5 replicates were set up with 50 larvae in each bottle.
Starvation resistance
Newly eclosed adult young flies were allowed to feed on HFD and standard diet with or without naringenin for 15 days. In the post-treatment, groups of single sex flies were kept in vials containing 1 % agar and transferred to fresh vials for every 5 h. Number of dead flies was recorded during each transfer. To prevent desiccation, vials were provided with filter paper strips dipped in distilled water. For each diet and sex, 10 replicate vials were set up with 30 flies in each vial.
Body weight
Newly eclosed, single sex flies were allowed to feed on HFD and standard diets with or without naringenin for 15 days. Flies were then subjected to mild etherisation followed by immediate measurement of body weight in batches of 10 flies. For each diet, 10 replicate vials were set up with 10 flies in each vial.
Statistical analysis
Results from longevity and starvation stress assays were calculated following Kaplan–Meier survival estimation and analysed by ANOVA with suitable post hoc tests. Remaining assays were analysed by ANOVA with suitable post hoc tests. Data represented as mean with error bars denoting (S.E.M.) or confidence intervals (C.I.) wherever applicable. All data were analysed using the Graph-pad prism software. For all assays, p values for levels of significance are represented as * <0.05, ** <0.01 and *** <0.001.
Results and discussion
Effect on fly longevity
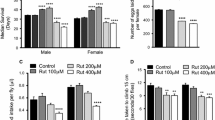
To examine the effect of naringenin on longevity, Canton-S flies were reared on standard and high fat diet (HFD) supplemented with varying concentrations of naringenin (50, 100, 200, 400, 600 and 800 µM). Median lifespan of male and female flies did not differ significantly when fed with standard diet having 50 and 100 µM of naringenin (Fig. 1a). In male flies fed with standard diet, significant pro-longevity was observed only at 400 µM of naringenin with median lifespan being 52 days (13.43 % increase, p < 0.05) as compared to 46 days in control. Median lifespan of male flies was 36 days (22.20 % decrease, p < 0.001) at 600 µM naringenin and 32 days (30.09 % decrease, p < 0.001) at 800 µM naringenin, both of which were significantly shorter as compared to control. In female flies fed with standard diet, significant pro-longevity was observed with median age of 45 days (16.92 % increase, p < 0.05) at 200 µM of naringenin and 47 days (22.62 % increase, p < 0.001) at 400 µM of naringenin as compared to 38 days in standard control (Fig. 1a). Higher concentrations of naringenin significantly shortened lifespan in female flies fed with standard diet, with median lifespan of 33 days (14 % decrease, p < 0.05) at 600 µM of naringenin and 28 days (27.4 % decrease, p < 0.001) at 800 µM of naringenin (Fig. 1a).

Effect of naringenin on fly longevity. a Survival of adult male and female flies fed with standard diet supplemented with varying concentrations of naringenin. Interaction between diet and sex was p < 0.001, two-way ANOVA. b Survival of adult male and female flies fed with high fat diet supplemented with varying concentrations of naringenin. Interaction between diet and sex was p < 0.001, two-way ANOVA. Data represented as median survival of flies (in days) with error bars denoting standard error of mean. Survival was calculated using Kaplan–Meier survival analysis. Symbols represented as *p < 0.05, **p < 0.01, ***p < 0.001, two-way ANOVA, Bonferroni post hoc test (control vs. naringenin supplemented diets). For each sex, replicates = 10, n = 30 flies
Recent studies have highlighted the reduced life expectancy associated with consumption of high fat diet (Chibnall 2010; Silberberg and Silberberg 1955). Median lifespan of male and female flies fed with HFD did not differ significantly at 50 and 100 µM of naringenin (Fig. 1b). As compared to median age of 31 days in HFD control, male flies survived longer only at 400 µM of naringenin with lifespan of 35 days (18.91 % increase, p < 0.05); longevity decreased significantly with median lifespan of 25 days (17.34 % decrease, p < 0.05) and 26 days (17.77 % decrease, p < 0.05) at 600 and 800 µM of naringenin respectively. In female flies fed with HFD, significant lifespan extension was observed at 200 and 400 µM of naringenin with median lifespan of 34 days (23.30 % increase, p < 0.01) and 35 days (25.85 % increase, p < 0.01) days respectively, as compared to 28 days in HFD control (Fig. 1b). However, higher concentrations of naringenin exhibited a sharp decrease in longevity in female flies with median lifespan of 21 days (24.34 % decrease, p < 0.01) and 22 days (23.26 % decrease, p < 0.01) at 600 and 800 µM of naringenin respectively (Fig. 1b).
In flies fed with HFD and standard diet respectively, 200 µM of naringenin was sufficient to prolong mean lifespan in female flies while significant lifespan increase was observed only at 400 µM of naringenin in male flies. The results obtained were in similar to the observations of a previous study in which C. elegans fed with 100 µM of naringenin failed to show any significant increase in lifespan (Grünz et al. 2012). Furthermore, higher concentrations of naringenin (600 and 800 µM) were lethal for male and female flies fed with HFD and standard diets respectively. This reiterates that dose–response relationships are crucial during evaluation of pro-longevity effects of a compound (Appenzeller-Herzog and Hall 2012; Singh et al. 2009). Interestingly, naringenin at 400 µM had disparate effects on percentage increase in mean lifespan of male and female flies fed with same diets (i.e., HFD and standard diet respectively). Thus, lifespan extending efficiency of a compound might be differentially influenced by factors such as gender or diet, as is evident from our data and previously published investigations (Bass et al. 2007; Wang et al. 2013).
Effect on fertility and feeding rate in flies
Various compounds that extended lifespan exhibited deleterious effects on standard physiological traits (Appenzeller-Herzog and Hall 2012; Chen et al. 2011; Li et al. 2007; Meijer and Codogno 2008). Altered reproduction and feeding behaviours are common trade-offs associated with prolonged lifespan (Appenzeller-Herzog and Hall 2012; Singh et al. 2009; van Acker et al. 2000; William et al. 2007). Thus, to ascertain whether naringenin prolonged lifespan at the expense of altered fecundity and feeding rate, we subjected flies fed with respective HFD and standard diet to varying concentrations of naringenin. As shown in Fig. 2a, fecundity in female flies fed with standard diet remained unaffected till 200 µM of naringenin, beyond which a significant and progressive decline in reproductive output was observed at 400 µM (p < 0.001), 600 µM (p < 0.001) and 800 µM (p < 0.001) of naringenin. Similarly, female flies fed with HFD exhibited reduced fecundity with 400 µM (p < 0.01), 600 µM (p < 0.001) and 800 µM (p < 0.001) of naringenin (Fig. 2b). This is a crucial observation since both 200 and 400 µM of naringenin extended lifespan in female flies fed with HFD and standard diet, respectively. However, from the fecundity data we can infer that enhanced survival of female flies at 400 µM of naringenin might be partly due to altered fecundity. This curtailed fecundity observed at higher concentrations of naringenin might be associated with diminished feeding rate. For further observation, flies were analyzed for any variations in feeding traits.

Effect of naringenin on female fecundity. a Number of eggs laid by a female on standard diet supplemented with varying concentrations of naringenin b Number of eggs laid by a female on high fat diet supplemented with varying concentrations of naringenin. Data represented as mean number of eggs laid by a female per day with error bars denoting standard error of mean. Symbols represented as **p < 0.01 and ***p < 0.001, one-way ANOVA, Dunnet’s post hoc test (control vs. naringenin supplemented diets). 10 replicate vials were set up for each diet, with two males and two females in each replicate
As evident from Fig. 3a, feeding rates in male and female flies fed with standard diet were comparable with control with no significant differences observed at 50, 100 and 200 µM of naringenin. However, a sharp reduction in feeding rate was observed at 400 µM (p < 0.001), 600 µM (p < 0.001) and 800 µM (p < 0.001) of naringenin in both male and female flies. Again, significantly diminished feeding rates were observed in male and female flies fed with HFD at higher concentrations of naringenin (400–800 µM; p < 0.001) while lower concentrations (50, 100 and 200 µM) did not have any marked effect (Fig. 3b). This can be partly explained in terms of innate lethality associated with high doses of a compound. In addition, naringenin is associated with a bitter taste (Konishi et al. 1983) and this might be another plausible explanation for the reduced food intake wherein the unsavoury taste of high concentrations of naringenin renders the diets unpalatable for flies.

Effect of naringenin on food intake in flies. a Average food intake in adult male and female flies fed with standard diet supplemented with varying concentrations of naringenin. Interaction existed with individual factors; p < 0.001 with diet p < 0.001 with sex, two-way ANOVA. b Average food intake in adult male and female flies fed with high fat diet supplemented with varying concentrations of naringenin. Interaction existed with individual factors; p < 0.001 with diet p < 0.001 with sex, two-way ANOVA. Data represented as average food intake (µl/day) with error bars denoting standard error of mean. Symbols represented as ***p < 0.001, two-way ANOVA, Bonferroni post hoc test (control vs. naringenin supplemented diets). For each diet and sex, replicates = 10, n = 2 flies
Collectively, while 200 µM of naringenin extended lifespan in female flies without affecting their feeding and reproductive capabilities, the efficacy of 400 µM of naringenin to extend lifespan in male and female flies was at the expense of curtailed fertility and feeding. Thus, critical analysis of various physiological traits in flies is warranted during experimenting with novel compounds for lifespan extension.
Effect on larval development
To determine the effects of different naringenin concentrations on larval development, we analyzed the percentage of pupae formation, the percentage of flies that eclosed following pupation and the eclosion time of new imagoes. Compared to control, standard diet supplemented with naringenin at 200 µM was associated with elevated percentage of pupae formation (p < 0.05) whereas a sharp decline in pupae formation was observed with naringenin at 600 µM (p < 0.01) and 800 µM (p < 0.001) (Fig. 4a). In HFD supplemented with varying concentrations of naringenin, no noticeable difference in pupae formation was observed at 50–400 µM, beyond which a significant drop in percentage of pupae formation was observed at 600 µM (p < 0.05) and 800 µM (p < 0.05) (Fig. 4b).

Effect of naringenin on larval development. a Percentage pupation in larva reared on standard diet supplemented with varying concentrations of naringenin. b Percentage pupation in larva reared on high fat diet supplemented with varying concentrations of naringenin. Data represented as percentage pupation with error bars denoting confidence intervals at p = 0.05. c Percentage eclosion in larva reared on standard diet supplemented with varying concentrations of naringenin. d Percentage eclosion in larva reared on high fat diet supplemented with varying concentrations of naringenin. Data represented as percentage eclosion with error bars denoting confidence intervals at p = 0.05. e Eclosion time of larvae reared on standard diet supplemented with varying concentrations of naringenin. f Eclosion time of larvae reared on high fat diet supplemented with varying concentrations of naringenin. Data represented as mean eclosion time in hours with error bars denoting standard error of mean. Numbers of pupae or flies in a given sample were monitored every 8 h and expressed as % of larvae that had pupated or flies that had eclosed by the given time period. Symbols represented as *p < 0.05, **p < 0.01 and ***p < 0.001, one-way ANOVA, Dunnet’s post hoc test (control vs. naringenin supplemented diets). For each diet, replicates = 5, n = 50 larvae
Next, we investigated the number of flies that eclosed after pupation following naringenin treatment. As shown in Fig. 4c, more number of flies emerged only at 200 µM of naringenin (p < 0.05) whereas naringenin at 50, 100 and 400 µM had no impact on eclosion percentage as compared to standard diet control; eclosion percentage was negatively affected with naringenin at 600 µM (p < 0.001) and 800 µM (p < 0.001). In HFD supplemented with naringenin, no significant differences in eclosion percentage were observed at 50–400 µM whereas significant decrease in eclosion percentage was observed at 600 µM (p < 0.001) and 800 µM (p < 0.001) (Fig. 4d).
Finally, we assessed changes in eclosion time with varying concentrations of naringenin. Eclosion time of flies was significantly shortened in standard diet supplemented with naringenin at 200 µM (p < 0.05 (Fig. 4e). Eclosion time remained unchanged in HFD supplemented with naringenin at 50–400 µM of whereas higher concentrations at 600 and 800 µM significantly stretched the eclosion time in flies (p < 0.001) (Fig. 4f).
Since early developmental stages in flies crucially influence adult-longevity, we hypothesised that concentrations of naringenin that extended lifespan in adult flies could positively affect larval development and vice versa. The data obtained were partly consistent with our hypothesis. Although 200 and 400 µM of naringenin extended lifespan in adult flies, positive effects of naringenin on larval development were observed only at 200 µM. Furthermore, this positive influence was restricted only to the standard diet. Higher concentrations of naringenin consistently proved to be lethal during developmental and adult stages of flies.
Effect on starvation stress in flies
Different naringenin concentrations were analyzed for resistance to starvation stress in male and female flies fed with standard diet and HFD respectively. For male flies fed with standard diet, there was no resistance to starvation when fed with naringenin at 50–400 µM as compared to control (Fig. 5a). Male flies were highly sensitive to starvation stress with naringenin at 600 µM with median lifespan of 24 h (p < 0.001) and 800 µM with lifespan of 23 h (p < 0.001) as compared to mean lifespan of 31 h in control. However, female flies were significantly resistant to starvation stress when fed with standard diet supplemented with naringenin at 200 and 400 µM with median lifespan being 45 h (p < 0.001) and 47 h (p < 0.001) respectively, as compared to 38 h in control (Fig. 5a). Female flies had median lifespan of 32 h (p < 0.001) with 600 µM of naringenin and 31 h (p < 0.001) with 800 µM of naringenin, which were significantly shorter as compared to control.

Effect of naringenin on resistance to starvation in flies. a Survival following starvation stress in adult male and female flies fed with standard diet supplemented with varying concentrations of naringenin. Interaction between diet and sex was p < 0.001, two-way ANOVA. b Survival following starvation stress in adult male and female flies fed with high fat diet supplemented with varying concentrations of naringenin. Interaction between diet and sex was p < 0.01, two-way ANOVA. Data represented as median survival of flies (in hours) with error bars denoting standard error of mean. Survival was calculated using Kaplan–Meier survival analysis. Symbols represented as ***p < 0.001, two-way ANOVA, Bonferroni post hoc test (control vs. naringenin supplemented diets). For each sex, replicates = 10, n = 30 flies
In male and female flies fed with HFD supplemented with naringenin at 50–400 µM, no significant differences were observed in median lifespan following starvation stress (Fig. 5b). Male flies survived shorter at higher concentrations of naringenin with median lifespan of 22 h (p < 0.001) at 600 µM and 21 h (p < 0.001) at 800 µM as compared to 26 h in control (Fig. 5b). Similarly, female flies were more sensitive to starvation stress when fed with naringenin at 600 µM with age of 26 h (p < 0.001) and 800 µM with median lifespan of 24 h (p < 0.001) at as compared to median lifespan of 31 h in control (Fig. 5b).
Taken together, naringenin at 200 and 400 µM made female flies fed on a standard diet more resistant to starvation stress. No positive effects on starvation stress were observed in female flies fed with HFD. Consistent with our previous data, higher concentrations of naringenin negatively affected starvation stress on flies of both sexes fed with HFD and standard diet respectively.
Effect on body weight
Different concentrations of naringenin were examined to observe any changes in the body weights of male and female flies fed with HFD and standard diet respectively. No changes in body weights of male and female flies fed with standard diet were observed with naringenin till 400 µM beyond which a significant reduction in body weight was observed at 600 µM (p < 0.001) and 800 µM (p < 0.001) in flies of both sexes (Fig. 6a). Previous investigations have reported the efficacy of naringenin in preventing obesity as a result of diet induced weight gain in rodents (Hsu and Yen 2008; Mulvihill et al. 2009). Similar trend was observed in flies fed with HFD. Interestingly, body weights of male and female flies fed with HFD progressively reduced with higher concentrations of naringenin (200–800 µM, p < 0.001, Fig. 6b). However, the observed body weight of flies fed with naringenin at 600 and 800 µM is below the normal range of 0.7 to 0.9 mg in males and 1.1 to 1.3 mg in females (Jumbo-Lucioni et al. 2010) which further accentuates the drawbacks associated with higher concentrations of naringenin.

Effect of naringenin on body weights in flies. a Mean body weight of adult male and female flies fed with standard diet supplemented with varying concentrations of naringenin. Interaction between diet and sex was p < 0.001, two-way ANOVA. b Mean body weight of adult male and female flies fed with high fat diet supplemented with varying concentrations of naringenin. Interaction between diet and sex was p < 0.001, two-way ANOVA. Data represented as mean weight (mg) per fly with error bars denoting standard error of mean. Symbols represented as ***p < 0.001, two-way ANOVA, Bonferroni post hoc test (control vs. naringenin supplemented diets). For each sex, replicates = 10, n = 10 flies
Modulatory effects of naringenin
Naringenin is a flavonoid with diverse bioactive properties (Guthrie and Carroll 1998; Mulvihill et al. 2009; Mulvihill and Huff 2010; van Acker et al. 2000). In addition, naringenin shares structural similarities with the extensively studied resveratrol (Manach et al. 2004). Studies concerning compound mediated lifespan extension in flies usually focus more on the efficacy of a certain dose of the compound while failing to report adversities associated with higher doses. Moreover, compound mediated lifespan extension does not necessarily imply enhancement in physiological traits (Joshi et al. 1996; Le Bourg 2001; Miquel and Economos 1979). Thus, the current study aimed to illustrate effects of varied concentrations naringenin on survival and development in flies fed with standard and high fat diets. Results from different assays reflected a hormetic influence of naringenin on fly longevity. Naringenin at lower concentrations (200 and 400 µM) enhanced fly survival and development while higher concentrations (600 and 800 µM) indicated an opposite outcome. Naringenin at 200 µM enhanced fly survival without negatively affecting physiological traits while increased mean lifespan at 400 µM was associated with reduced fecundity and food intake in flies. Previous investigations have highlighted that bioactive compounds might be beneficial at low doses while detrimental at high doses (Calabrese et al., 2015; Hoffmann 2009). Such a biphasic dose response phenomenon is known as hormesis, and the conditions, such as flavonoids, polyphenols and various other natural and synthetic molecules which cause hormesis are known as hormetins (Rattan, 2012). Thus it is of importance to understand the limits of positivity and negativity (Le Bourg 2015). Data from this study emphasize the significance of dose–response relationships while assessing a novel compound for pro-longevity attributes. Furthermore, potency of naringenin was context dependent that varied with diet, gender and developmental stage of flies. Also, as stated previously, naringenin is known to interact with various common drugs (Mozayani and Raymon 2011). For naringenin, there exists a narrow range of concentration within which it exerts it efficacy. Thus, naringenin should not be treated as a panacea. Even though naringenin possesses various physiologically beneficial capabilities, strong caution is warranted whilst unmindful consumption of bulk portion of grapefruits. Further studies are required to elucidate the plausible mechanisms by which naringenin modulates fly-longevity on different diets.
References
Appenzeller-Herzog C, Hall MN (2012) Bidirectional crosstalk between endoplasmic reticulum stress and mTOR signaling. Trends Cell Biol 22:274–282
Bao Y, Fenwick R (2004) Phytochemicals in health and disease. CRC Press, Boca Raton, FL
Bass TM, Weinkove D, Houthoofd K, Gems D, Partridge L (2007) Effects of resveratrol on lifespan in Drosophila melanogaster and Caenorhabditis elegans. Mech Ageing Dev 128:546–552
Benavente-García O, Castillo J, Marin FR, Ortuño A, Del Río JA (1997) Uses and properties of citrus flavonoids. J Agric Food Chem 45:4505–4515
Borradaile NM, de Dreu LE, Huff MW (2003) Inhibition of net HepG2 cell apolipoprotein B secretion by the citrus flavonoid naringenin involves activation of phosphatidylinositol 3-kinase, independent of insulin receptor substrate-1 phosphorylation. Diabetes 52:2554–2561
Calabrese EJ, Gaurav D, Rachna K, Ivo I, Vittorio C (2015) What is hormesis and its relevance to healthy aging and longevity? Biogerontology 16:693–707
Chen Z, Li Y, Han J, Wang J, Yin J, Li J, Tian H (2011) The double-edged effect of autophagy in pancreatic beta cells and diabetes. Autophagy 7:12–16
Chibnall J (2010) Healthy brain aging: a road map. Clin Geriatr Med 26:116Dietz
Davies JN, Hobson GE, McGlasson W (1981) The constituents of tomato fruit—the influence of environment, nutrition, and genotype. Crit Rev Food Sci Nutr 15:205–280
Fontana L, Partridge L, Longo VD (2010) Extending healthy life span—from yeast to humans. Science 328:321–326
Grünz G, Haas K, Soukup S, Klingenspor M, Kulling SE, Daniel H, Spanier B (2012) Structural features and bioavailability of four flavonoids and their implications for lifespan-extending and antioxidant actions in C. elegans. Mech Ageing Dev 133:1–10
Guthrie N, Carroll K (1998) Inhibition of mammary cancer by citrus flavonoids. In: Manthey JA, Buslig BS (eds) Flavonoids in the living system. Springer, Berlin, pp 227–236
Hoffmann GR (2009) A perspective on the scientific, philosophical, and policy dimensions of hormesis. Dose-Response 7:08–023
Hsu CL, Yen GC (2008) Phenolic compounds: evidence for inhibitory effects against obesity and their underlying molecular signaling mechanisms. Mol Nutr Food Res 52:624–625
Joseph JA, Shukitt-Hale B, Lau FC (2007) Fruit polyphenols and their effects on neuronal signaling and behavior in senescence. Ann N Y Acad Sci 1100:470–485
Joseph JA, Shukitt-Hale B, Willis LM (2009) Grape juice, berries, and walnuts affect brain aging and behavior. J Nutr 139:1813S–1817S
Joshi A, Shiotsugu J, Mueller LD (1996) Phenotypic enhancement of longevity by environmental urea in Drosophila melanogaster. Exp Geront 31:533–544
Jumbo-Lucioni P, Ayroles JF, Chambers MM, Jordan KW, Leips J, Mackay TF, De Luca M (2010) Systems genetics analysis of body weight and energy metabolism traits in Drosophila melanogaster. BMC Genom 11:297
Kawaii S, Tomono Y, Katase E, Ogawa K, Yano M (1999) Quantitation of flavonoid constituents in citrus fruits. J Agric Food Chem 47:3565–3571
Konishi F, Kamiya S, Esaki S (1983) Synthesis and taste of naringenin 7-O-(2-O-α-l-lyxopyranosyl-β-d-galactopyranoside) and its dihydrochalcone derivative. Agric Biol Chem 47:1633–1635
Le Bourg É (2001) Oxidative stress, aging and longevity in Drosophila melanogaster. FEBS Lett 498:183–186
Le Bourg É (2015) Fasting and other mild stresses with hormetic effects in Drosophila melanogaster can additively increase resistance to cold. Biogerontology 16:517–527
Lee K-S et al (2010) Curcumin extends life span, improves health span, and modulates the expression of age-associated aging genes in Drosophila melanogaster. Rejuvenation Res 13:561–570
Li P, Callery PS, Gan L-S, Balani SK (2007) Esterase inhibition by grapefruit juice flavonoids leading to a new drug interaction. Drug Metab Dispos 35:1203–1208
Manach C, Scalbert A, Morand C, Rémésy C, Jiménez L (2004) Polyphenols: food sources and bioavailability. Am J Clin Nutr 79:727–747
Meijer AJ, Codogno P (2008) Autophagy: a sweet process in diabetes. Cell Metab 8:275–276
Middleton E Jr, Kandaswami C (1994) The impact of plant flavonoids on mammalian biology: implications for immunity, inflammation and cancer. Chapman and Hall, The flavonoids London
Miquel J, Economos AC (1979) Favorable effects of the antioxidants sodium and magnesium thiazolidine carboxylate on the vitality and life span of Drosophila and mice. Exp Geront 14:279–285
Mozayani A, Raymon L (2011) Handbook of drug interactions: a clinical and forensic guide. Springer, Berlin
Muller AP, de Oliveira Dietrich M, de Assis AM, Souza DO, Portela LV (2013) High saturated fat and low carbohydrate diet decreases lifespan independent of body weight in mice. Longev Healthspan 2:10
Mulvihill EE, Huff MW (2010) Antiatherogenic properties of flavonoids: implications for cardiovascular health. Can J Cardiol 26:17A–21A
Mulvihill EE et al (2009) Naringenin prevents dyslipidemia, apolipoprotein B overproduction, and hyperinsulinemia in LDL receptor–null mice with diet-induced insulin resistance. Diabetes 58:2198–2210
Ortiz-Andrade R et al (2008) Antidiabetic and toxicological evaluations of naringenin in normoglycaemic and NIDDM rat models and its implications on extra-pancreatic glucose regulation. Diabetes Obes Metab 10:1097–1104
Rattan SIS (2012) Rationale and methods of discovering hormetins as drugs for healthy aging. Exp Op Drug Disc 7:439–448
Ruh MF, Zacharewski T, Connor K, Howell J, Chen I, Safe S (1995) Naringenin: a weakly estrogenic bioflavonoid that exhibits antiestrogenic activity. Biochem Pharmacol 50:1485–1493
Semalty A, Semalty M, Singh D, Rawat M (2010) Preparation and characterization of phospholipid complexes of naringenin for effective drug delivery. J Inclusion Phenom 67:253–260
Silberberg R, Silberberg M (1955) Life span of mice fed a high fat diet at various ages. Can J Biochem Physiol 33:167–173
Singh R et al (2009) Autophagy regulates lipid metabolism. Nature 458:1131–1135
Ueng Y-F, Chang Y-L, Oda Y, Park SS, Liao J-F, Lin M-F, Chen C-F (1999) In vitro and in vivo effects of naringin on cytochrome P450-dependent monooxygenase in mouse liver. Life Sci 65:2591–2602
van Acker FA, Schouten O, Haenen GR, van der Vijgh WJ, Bast A (2000) Flavonoids can replace α-tocopherol as an antioxidant. FEBS Lett 473:145–148
Wagner D, Spahn-Langguth H, Hanafy A, Koggel A, Langguth P (2001) Intestinal drug efflux: formulation and food effects. Adv Drug Del Rev 50:S13–S31
Wang C et al (2013) The effect of resveratrol on lifespan depends on both gender and dietary nutrient composition in Drosophila melanogaster. Age 35:69–81
William WJ et al (2007) Prandiology of Drosophila and the CAFE assay. Proc Natl Acad Sci 104:8253–8256
Wunderlich SM, Piemonte J (2012) Food and drug interactions. In: Mozayani A, Raymon L (eds) Handbook of drug interactions. Springer, Berlin, pp 479–497
Acknowledgments
We thank Dr. N.B. Ramachandra at Mysore University, National Drosophila Stock Centre, India, for providing the wild-type Canton-S fly strain.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of Interest
No potential conflicts of interest were disclosed.
Rights and permissions
About this article

Cite this article
Chattopadhyay, D., Sen, S., Chatterjee, R. et al. Context- and dose-dependent modulatory effects of naringenin on survival and development of Drosophila melanogaster . Biogerontology 17, 383–393 (2016). https://doi.org/10.1007/s10522-015-9624-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10522-015-9624-6